
La variabilità genetica nei batteri
Le due cellule figlie sono geneticamente identiche tra
loro e alla cellula madre
La variabilità genetica nei batteri
Nella maggioranza dei casi le mutazioni sono deleterie…..
La variabilità genetica nei batteri
…..ma in qualche caso possono conferire un vantaggio!
Le basi molecolari delle mutazioni
Mutazione: è un cambiamento ereditario nella sequenza delle basi nel
genoma di un organismo
Mutazioni puntiformi
possono risultare per sostituzione di una base nel DNA o per inserzione e
delezione di una base (microinserzioni e microdelezioni). Il cambiamento
del fenotipo dipende dal punto esatto in cui la mutazione è situata nel gene
e da quale prodotto è normalmente codificato dal gene.
Mutazioni che coinvolgono molte paia di basi
Le delezioni possono coinvolgere la perdita di centinaia o migliaia di paia di
basi; non revertono se non per ricombinazione.
Le inserzioni si verificano per aggiunta di nuove basi al DNA e inattivano il
gene in cui accadono. Molte mutazioni da inserzione sono dovute a
sequenze di DNA specifiche, lunghe 700-1400 bp, dette sequenze di
inserzione. Le mutazioni da inserzione possono revertere.
Effetti della sostituzione di una base
mutazione mutazione mutazione
missenso nonsenso
silente
Scivolamento dello schema di lettura (frameshift)
Frequenza di mutazione
La frequenza con cui avvengono i diversi tipi di mutazioni è estremamente
variabile. Errori nella replicazione del DNA ricorrono con una frequenza di
10-7 10-11 per coppia di basi per singolo ciclo di replicazione. Per un gene di
1000 basi la frequenza è dell’ordine di 10-4 10-8 per generazione
Retromutazione e Soppressione
Un revertante è un ceppo in cui viene ripristinato il fenotipo selvatico
Reversione vera (retromutazione)
Reversione di secondo sito (soppressione)
mutazione frameshift
mutazione in un secondo gene
Reversione di mutazione nonsenso
Reversione di delezioni e inserzioni
Grosse delezioni possono revertere solo mediante processi che coinvolgono
la ricombinazione genetica.
Alcune inserzioni (elementi trasponibili) possono revertere per escissione.
La Mutagenesi
Le mutazioni possono essere:
Spontanee
Indotte da mutageni
(frequenza molto bassa)
Mutageni chimici
Analoghi delle basi
Agenti alchilanti
Agenti intercalanti
Mutageni fisici
Radiazioni non ionizzanti:UV
Radiazioni ionizzanti:
raggi gamma
raggi X
raggi cosmici
Analoghi delle basi
Simili nella struttuta a purine e pirimidine del DNA ma difettosi nell’appaiamento
Il più delle volte la replicazione avviene
normalmente ma occasionalmente può
verificarsi un errore che verrà fissato
come mutazione nel successivo ciclo di
replicazione
T:A
BrU
BrU
:A
BrU
:G
:A
C:G
Agenti alchilanti
Nitrosoguanidina, Acido nitroso, Idrossilamina sono responsabili
di reazioni di metilazione, deaminazione, ecc.
Mutageni molto potenti e inducono mutazioni ad alta frequenza
rispetto agli analoghi delle basi.
Reagendo sul DNA sono in grado di introdurre cambiamenti
anche in assenza di replicazione
Agenti intercalanti
Acridine (arancio di acridina, proflavina): sono molecole planari
che si inseriscono tra due basi del DNA, separandole
Questa conformazione anormale può portare a microinserzioni e
microdelezioni durante la replicazione
Radiazioni non ionizzanti: raggi UV
Le basi degli acidi nucleici assorbono efficientemente la
radiazione ultravioletta. Il picco di assorbimento è a 269 nm
La radiazione UV a 260 nm è l’agente letale più efficiente
Il suo effetto più conosciuto sul DNA è la formazione di dimeri
di pirimidine (due C o T adiacenti vengono legate covalentemente
tra loro). Induzione del sistema SOS.
La presenza di dimeri aumenta la probabilità che la DNA
polimerasi inserisca in questa posizione un nucleotide errato.
La radiazione UV è uno strumento molto utile nell’isolamento di
mutanti
Radiazioni ionizzanti
Sono radiazioni a lunghezza d’onda corta (raggi X, gamma, raggi
cosmici). Più potenti dei raggi UV e altamente penetranti.
Causano la ionizzazione dell’acqua e di altre sostanze con la
produzione di radicali liberi come lo ione ossidrile (OH-)
La loro azione mutagena è indiretta in quanto il danno è prodotto
dai radicali liberi.
La variabilità genetica nei batteri
Le mutazioni puntiformi non sono sufficienti a spiegare la diversità
del mondo microbico
Il genoma di un batterio risulta costituito da un “puzzle” di sequenze
provenienti da altri batteri che non sono suoi progenitori ma suoi
contemporanei
Indifferenti al mantenimento della loro identità, i batteri sono
capaci di acquisire da altri organismi geni che permettono loro:
di diventare resistenti ad antibiotici
causare nuove malattie
aumentare la loro possibilità di sopravvivenza
Lo scambio di DNA tra i batteri è chiamato:
Trasferimento genico orizzontale
Trasferimento genico orizzontale
Scambio di DNA tra batteri contemporanei
Trasferimento genico verticale
Eredità di un gene
dal progenitore
Significato medico del trasferimento genico
Il trasferimento genico orizzontale tra i batteri sta avendo
effetti immediati e pratici sulla salute umana
E’ stato dimostrato il trasferimento di geni:
per la resistenza ad antibiotici
responsabili della patogenicità
per la degradazione di inquinanti
(biorisanamento)
geni per la tossina
Shigella
E. coli
(ceppo che causa diarrea)
E. coli O157:H7
(causa diarrea emorragica e
danni renali)
geni per la resistenza
Ceppo X resistente
alla penicillina
Ceppo Y sensibile
alla penicillina
Ceppo Y resistente
Trasduzione
Trasformazione
Coniugazione
Trasformazione
Processo in cui il DNA presente all’esterno del batterio viene
internalizzato nella cellula ospite, integrato nel cromosoma
nella corrispondente regione omologa e stabilmente ereditato
con le divisioni cellulari
L’esperimento di Griffith (1928)
Pneumococchi da colonie liscie
(S= smooth)
cellule vive S
Pneumococchi da colonie rugose
(R= rough)
cellule vive R
L’esperimento di Griffith (1928)
cellule
morte
cellule
vive
cellule
morte
L’esame batteriologico
rivela batteri vivi di
tipo S
Trasformazione naturale
Batteri Gram (+)
Streptococcus pneumoniae e S. sanguis
Bacillus subtilis, B. cereus, B. stearothermophilus
Batteri Gram (-)
Neisseria gonorrheae
Haemophilus influenzae, H. parainfluenzae
Acinetobacter calcoaceticus
Moraxella oslensis, M. urethalis
Psycrobacter sp.
Azotobacter agilis
Pseudomonas stutzeri
Possibili benefici derivanti dalla trasformazione
Acquisizione di nuove caratteristiche genetiche
Riparo del DNA danneggiato
DNA come fonte di nutrimento
Superamento delle difese dell’ospite da parte
dei patogeni (N. gonorrhoeae)
Le fasi del processo di trasformazione naturale
sviluppo della fase di competenza
legame del DNA trasformante alle cellule competenti
ingresso del DNA trasformante
integrazione del DNA (ricombinazione)
espressione del DNA trasformante
Sviluppo della fase di competenza
Nella maggioranza dei batteri la competenza è geneticamente
regolata e si manifesta in condizioni ambientali particolari o in una
fase specifica della crescita della popolazione
Nei batteri Gram negativi ogni individuo di una popolazione sviluppa la
competenza indipendentemente
Nei batteri Gram positivi lo sviluppo della competenza è regolato da
uno scambio di messaggi chimici (feromoni) tra le cellule di una
popolazione
Batteri G negativi
Neisseria gonorrhoeae
La competenza è costitutiva. Quasi tutte le cellule di una coltura
sono competenti e non c’è nessuna relazione con la fase di crescita
Haemophilus influenzae
La competenza è indotta da condizioni che inibiscono la crescita
Batteri G positivi
Streptococcus pneumoniae
La competenza è controllata da un fattore extracellulare (ferormone) di 17 aa.
Quasi tutte le cellule di una coltura in fase esponenziale diventano competenti
quando il feromone raggiunge una concentrazione critica (quorum sensing).
Il 100% delle cellule possono diventare competenti (per pochi minuti!).
Il regulone com
Bacillus subtilis
La concentrazione critica di almeno due feromoni attiva la competenza. Si
manifesta in tarda fase esponenziale o inizio di fase stazionaria. Solo il 10%
dei batteri diventano competenti.
La trasformazione naturale in Streptococcus pneumoniae
Il fattore di competenza attiva la produzione di altre proteine (>12)
che permettono l’adsorbimento del DNA sulla superficie batterica.
I frammenti di DNA sono generalmente lunghi 15-20 Kb.
Una cellula competente è in grado di legare fino a 1000 volte più
DNA di una cellula non competente
Il DNA a doppia elica viene frammentato in piccoli frammenti per
azione di nucleasi
Un’elica del DNA viene degradata da una nucleasi mentre l’elica
complementare entra nella cellula associata a proteine che la
proteggono dalla degradazione
Può entrare DNA di qualsiasi origine ma solo quello omologo viene
integrato e produrre cambiamenti ereditari nella cellula ricevente
Batterio Gram positivo
La trasformazione naturale in Haemophilus influenzae
Non vengono prodotti fattori di competenza
E’ trasformabile solo da DNA appartenente a specie correlate
Il DNA trasformante deve contenere una sequenza specifica di
11 bp. Questa sequenza è ripetuta diverse centinaia di volte nel
proprio genoma
Il DNA attraversa la membana esterna come doppia elica in
vescicole di membrana
Dallo spazio periplasmico entra nel citoplasma come singola elica
DNA trasformante
ingresso
SS-DNA
D-loop
cromosoma
RecA
DNA ligasi
endonucleasi
Pol III exo
La trasformazione artificiale
La maggior parte dei batteri possono essere trasformati artificialmente
Vengono utilizzati shock chimici e termici (CaCl2)
o elettrici (elettroporazione)
Il DNA viene introdotto integro nella cellula
DNA lineare a
doppia elica
integrazione mediante
ricombinazione omologa
DNA plasmidico
replicazione del plasmide
Lederberg
& Tatum
1946
pressione o
aspirazione
cotone
ceppo A
ceppo B
La coniugazione richiede un
contatto cellula-cellula: quando le
colture sono separate da un filtro
poroso permeabile alle
macromolecole ma non ai batteri,
la ricombinazione non avviene!
filtro poroso
Coniugazione
Pili sessuali: presenti in numero di 1-10 per cellula, sono spessi 9-10 nm
F+
F-
F
donatore
ricevente
ponte coniugativo
OriT
OriT
+
F
F-
Viene trasferito un filamento singolo di F. Nel ricevente viene sintetizzato
il filamento complementare
+
F
Una copia del cromosoma con F rimane nella cellula donatrice dopo la
replicazione della singola elica restante
F+
Il DNA può essere diviso
in 4 regioni:
1. Regione tra (35 Kb)
2. Regione della replicazione
(inc, rep)
3. Elementi trasponibili
4. Regione silente
F
H
D I
GS
IS3
gd
IS3
C
B
F
K
E
L
94 Kb
A
J
oriT inc, rep
Regione tra:
•Sintesi ed assemblaggio dell’F pilus (13 geni)
•Mating-pair stabilization (2 geni)
•Metabolismo del DNA coniugativo (5 geni)
•Regolazione del trasferimento (3 geni)
•Esclusione di superficie (2 geni)
IS2
OriT
F-
Hfr
donatore
ricevente
OriT
Interruzione dell’accoppiamento
Hfr
F-
donatore
ricevente
Viene trasferito un filamento singolo di F insieme a una copia di parte del
cromosoma del donatore. Nel ricevente viene sintetizzato il filamento
complementare
Una copia del cromosoma con F integrato rimane nella cellula donatrice
dopo la replicazione della singola elica restante
Ricombinazione dopo coniugazione con Hfr
esogenote
endogenote
Modello di Campbell
F
e
OriT
OriT
a
1
5
a
e
2
d
4
d
b
3
b
c
c
1
2
3
a
b
c
d
e
b
c
d
e
a
c
d
e
a
b
G
H
S D
I
IS3
gd
F
IS3
C
B
F
K
94 Kb
E
L
A
J
oriT
inc, rep
IS2
Gradiente di trasferimento
Il fattore F’ e la F-duzione
Si ottiene in seguito
a un errato processo
di escissione di F
La formazione di un F’
produce una delezione
nel cromosoma
In seguito ad incrocio:
F’ x Fsi produce un diploide
parziale o merizigote
Ricapitolando……
Il fattore F può trovarsi nella cellula come:
1. F+
2. Hfr
3. F’
da un incrocio……
i riceventi……
F+ x F-
diventano F+
Hfr x F-
rimangono F- ma possono
acquisire nuovi caratteri
F’ x F-
diventano F’ e diploidi
parziali per uno o più geni
Coniugazione in altri batteri Gram negativi
Salmonella:
Sono stati isolati diversi plasmidi coniugativi in ceppi
naturali di Salmonella sp., ma per gli studi di genetica
viene utilizzato l’F di Escherichia coli
Pseudomonas:
Molti plasmidi coniugativi e mobilizzabili sono stati
descritti e molto studiati in ceppi di Pseudomonas.
Il plasmide FP2 di P. aeruginosa può mobilizzare anche il
cromosoma:
mobilizzazione unidirezionale
a partire da un solo sito cromosomale
Coniugazione in altri batteri Gram positivi
1.
Mediante plasmidi coniugativi simili al fattore F
(Streptomyces)
•
Mediata da FEROMONI (Streptococcus)
responsabili di aggregazione cellulare
•
Coniugazione traspositiva (Bacillus, Staphilococcus,
Streptococcus)
In Streptomyces coelicolor il plasmide SCP1 interagisce
con il cromosoma batterico in diversi siti.
Due differenze con F:
trasferimento bidirezionale
tutti i ricombinanti ottenuti sono fertili
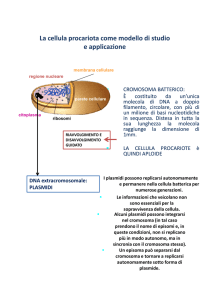
I plasmidi
Elementi genetici circolari, extracromosomali, capaci di replicazione
autonoma (replicone)
Le dimensioni variano da 1 Kb a >1000 Kb e il loro numero varia da 1 a
diverse centinaia di copie per cellula ospite
La loro informazione genetica non è essenziale per la cellula ospite ma
può conferire un vantaggio selettivo in particolari condizioni
Alcuni plasmidi possono esistere sia allo stato libero sia integrato nel
cromosoma dell’ospite (episomi)
Alcuni plasmidi si mantengono stabilmente all’interno della cellula,
altri hanno la capacità di diffondersi attraverso la coniugazione
H
F
C
B
K
E
L
A
G SD
I
IS3
Plasmidi coniugativi e mobilizzabili
gd
IS3
IS2
F
94 Kb
J
oriT inc, rep
+
F
F-
Plasmidi coniugativi e mobilizzabili
C
M
C
M
C
M
M
Plasmidi R
Codificano per enzimi capaci di distruggere o modificare gli antibiotici come
ampicillina, cloramfenicolo, tetraciclina, kanamicina ecc.
Alcuni plasmidi portano un’unica resistenza, altri fino a otto.
Possono essere anche coniugativi e diffondersi rapidamente.
Plasmidi col
Conferiscono un vantaggio al batterio portatore nei confronti di batteri della
stessa specie o specie affini.
Codificano per le batteriocine (colicine, subtilisine, ecc.) capaci di
permeabilizzare le membrane o degradare gli acidi nucleici.
Plasmidi di virulenza
Portatori di tossine; rendono il ceppo batterico più patogeno.
Il ceppo di E. coli responsabile della “diarrea del viaggiatore” è portatore di un
plasmide codificante per una enterotossina.
Plasmidi metabolici
Codificano per enzimi capaci di degradare composti aromatici, pesticidi,
zuccheri ecc. In alcuni ceppi di Rhizobium portano i geni necessari per la
nodulazione nelle leguminose.
Plasmidi di virulenza e malattie
Molti batteri risultano patogeni a causa dei loro plasmidi
Il plasmide porta geni per le tossine
“
rende il batterio più capace di infettare l’ospite
“
rende il batterio più resistente alle difese dell’ospite
Ceppi enterotossici di E. coli provocano la diarrea del viaggiatore
Tossina termolabile (LT)
Tossina termostabile (ST)
sono portate da plasmidi, a volte
anche da un solo plasmide
I ceppi enterotossici di E. coli devono essere anche capaci di colonizzare
l’intestino
Speciali fimbrie adesive sono codificate da geni di un altro plasmide
Altri batteri patogeni portano plasmidi di virulenza
Tossina tetanica di Clostridium tetani
Antratossina di Bacillus anthracis
Incompatibilità fra plasmidi
L’incompatibilità si osserva quando un plasmide entra in una cellula dove è
già presente un plasmide simile.
Il nuovo plasmide non può essere mantenuto e viene perso nei successivi
cicli di divisione batterica.
Il fenomeno è controllato dai geni coinvolti nella regolazione della
replicazione plasmidica
Plasmidi che condividono lo stesso sistema di replicazione appartengono
allo stesso gruppo di incompatibilità (Inc) e sono tra loro incompatibili
Perchè un batterio possa contenere diversi tipi di plasmidi, questi non
devono essere strettamente correlati
Sistema host cell killing del plasmide R1
Il sistema hok/sok di alcuni plasmidi a basso numero di copie assicura
il mantenimento del plasmide all’interno di una popolazione batterica
hoc mRNA: emivita 20 min
hok
SD
128 bp
sok mRNA: emivita < 1-2 min
sok
Il sistema veleno-antidoto del plasmide F
ccdA
72 AA
antidoto (facilmente
degradabile)
blocca l’attività di CcdB
ccdB
101 AA
veleno (proteina stabile)
interagisce con la DNA girasi
Trasduzione
Modalità si scambio genetico fra batteri mediato da un virus
Trasduzione generalizzata
Trasduzione specializzata
Qualsiasi marcatore genetico può
essere trasferito da un donatore
a un ricevente
Solo marcatori specifici possono
essere trasferiti
P22, P1 per Gram(-)
PBS1 per Gram (+)
l per E. coli
SPb per B. subtilis
Anche se non tutti i fagi trasducono e non tutti i batteri possono
essere trasdotti, il fenomeno è sufficientemente diffuso da far
supporre che svolga un ruolo importante nel trasferimento genico
in natura
Lederberg e Zinder
1951
S.typhimurium
infettato da P22
filtro poroso
testa
collare
coda
guaina elicoidale
piastra basale
fibre caudali
Batteriofagi virulenti
ciclo litico
Batteriofagi temperati
ciclo litico
ciclo lisogenico
induzione
profago
Il processo di infezione virale
DNA batterico
DNA virale
Trasduzione generalizzata
I fagi capaci di dare trasduzione generalizzata devono possedere
un meccanismo di impacchettamento del DNA che permetta il
riconoscimento accidentale del DNA dell’ospite
Il fago trasducente (1/103 della popolazione
virale) può infettare un altro batterio ma non
iniziare un normale ciclo di infezione in
quanto il DNA virale è assente (particella
difettiva)
fago trasducente
La quantità di DNA
batterico trasportata
dipende dalle dimensioni del
capside. Il fago P1 di E. coli
trasporta 99 Kb (2% del
cromosoma batterico)
Nuovi alleli, o anche nuovi geni,
possono essere integrati nel
cromosoma ospite. Dal 70 al 90%
del DNA trasdotto non viene
integrato, ma può esprimersi per
un numero limitato di generazioni,
dando dei diploidi parziali
(trasduzione abortiva)
x
a+
a-
x
Trasduzione specializzata
attP
attB
l
Escissione corretta
ldgal
Escissione errata
(evento raro).
Si genera una particella
trasducente per il gene
gal.
Il fago è difettivo
Lisato a bassa frequenza di trasduzione (lisato LFT)
Trasduzione per ricombinazione
Le particelle trasducenti difettive possono aver perso alcuni geni
essenziali per la propria riproduzione
Hanno un sito di integrazione ibrido, non funzionale
Trasduttanti stabili possono derivare dalla ricombinazione tra il
cromosoma del fago e quello del batterio in seguito a due crossingover su entrambi i lati del sito gal
Trasduzione in un batterio lisogeno
I fagi ldgal possono integrarsi in un batterio lisogeno in quanto il
sito att ibrido è identico a quelli generati dal profago
Il fago normale fornisce anche i geni persi dal fago difettivo e
viene chiamato fago helper, perché aiuta il fago difettivo a
integrarsi e riprodursi
I trasduttanti sono instabili perché il profago può essere indotto
all’escissione
Lisato ad alta frequenza di trasduzione (lisato HFT)
Caratteristiche comuni ai tre meccanismi di trasferimento genico
1. Solo una porzione del cromosoma è trasferita
2. Il trasferimento è sempre polarizzato DONATORE
RICEVENTE
3. Il prodotto del trasferimento non è un vero zigote: per questo viene
chiamato MERIZIGOTE
4. Il DNA trasferito (ESOGENOTE) può essere trasmesso alla
progenie solo se viene incorporato nel genoma della cellula ricevente
(ENDOGENOTE)
CASO PARTICOLARE: trasferimento di un replicone (PLASMIDE)
I processi di trasferimento genico hanno un ruolo importante nella
variabilità genetica delle popolazioni batteriche in quanto
assicurano la diffusione di materiale genetico e quindi l’acquisizione
di nuovi caratteri



![mutazioni genetiche [al DNA] effetti evolutivi [fetali] effetti tardivi](http://s1.studylibit.com/store/data/004205334_1-d8ada56ee9f5184276979f04a9a248a9-300x300.png)







![(Microsoft PowerPoint - PCR.ppt [modalit\340 compatibilit\340])](http://s1.studylibit.com/store/data/001402582_1-53c8daabdc15032b8943ee23f0a14a13-300x300.png)