Disgenesia degli ibridi in Drosophila melanogaster:
sistema PM
Il fenomeno fu descritto inizialmente nel 1977 da Margaret e James Kidwell, ed un altro
ricercatore australiano John Sved -> alcuni incroci ed in particolare tra maschi di
“popolazioni naturali” e femmine “di laboratorio” causavano degli effetti negativi nella
linea germinale della progenie. Tutti gli effetti sono stati definiti “disgenesia degli ibridi”
Questo effetto non si aveva se femmine di popolazioni naturali erano incrociate con maschi
da laboratorio.
Tra gli effetti c’era un’alta frequenza di mutazione, aberrazioni cromosomiche e sterilità
, questi effetti erano influenzati dalla temperatura
ALTA temperatura -> produceva effetti più drastici e sterilità completa
BASSA temperatura -> effetti minori ( mutazioni)
Poiché gli effetti erano descritti solo quando maschi di popolazioni naturali
erano incrociati con femmine di laboratorio i ceppi che determinavano gli effetti
furono chiamati “ceppi P” (da contributo paterno) gli altri, quelli che non davano
questi effetti furono chiamati “ceppi “M” ( da materno).
Incroci disgenici
Femmina parentale
M
P
Incrocio DISGENICO
F1
M
P
Progenie
normale
Progenie
normale
Progenie
disgenica
M
Progenie
normale
X
P
Sviluppo anormale della linea germinale
Fertilità ridotta
Elevato tasso di mutazione
Riarrangiamenti cromosomici
Linea germinale normale e disgenica
Linea germinale normale e disgenica
Caratteristica delle mutazioni prodotte a
seguito della DISGENESIA
• avevano un’alta frequenza
• revertivano con alta frequenza (non influenzata da mutageni chimici)
Avevano le stesse caratteristiche delle mutazioni
batteriche dovute a elementi IS
La DISGENESIA DEGLI IBRIDI (HD) era dovuta ad
elementi genetici mobili?
L’ipotesi fu che i cromosomi dei ceppi P portano “fattori genetici P” che sono alla base del fenomeno della
disgenesia degli ibridi
Dimostrazione del ruolo degli elementi P nella disgenesia
• erano stati appena isolati 10 mutanti white a seguito della HD
• il locus white era appena stato clonato dal gruppo di Jerry Rubin e fu quindi possibile
isolare il gene white da questi mutanti.
• furono identificate nella sequenza di tutti i mutanti delle sequenze addizionali di
DNA, variabili in lunghezza ma OMOLOGHE tra loro
• queste sequenze simili tra loro furono definite come appartenenti ad una famiglia di
elementi chiamati “elementi P”
• Gli elementi P sono presenti in tutti i ceppi definiti P e assenti dai ceppi definiti M
(analisi per Southern)
• Gli elementi P si muovono dopo HD ( analisi per ibridazione in situ)
Regione cromosomica del gene white
telomero
EcoRI
EcoRI
BamHI
HindIII
XbaI
HindIII
XbalI
+10
HindIII
BamHI
+5
SacI
SalI
SalI
SalI
EcoRI
HindIII
HindIII
SacI
EcoRI
HindIII
0
-5
-10
BamHI
BamHI
-15
centromero
Cloni isolati da mutanti white ottenuti per disgenesia degli ibridi
#6:
6.1
cloni dal mutante #6
6.4
#12:
12.3
12.4
12.6
12.12
cloni dal mutante #12
Clonaggio dell’elemento P
Costruzione di una libreria genomica da un ceppo che
aveva gli elementi P, definito 2
Screening con i frammenti “non white” ottenuti dai
mutanti white per HD
40 cloni positivi -> 12 furono caratterizzati per
mappa di restrizione
Le inserzioni più grandi erano di 2,9 kb
Le mappe di restrizione delle STESSE REGIONI isolate da
ceppi M non mostravano SEQUENZE ADDIZIONALI
Sequenze non-white usate come sonda per isolare elementi P
Cloni isolati da mutanti white ottenuti per disgenesia degli ibridi
#6:
6.1
SEQUENZE “NON-WHITE” IDENTIFICATE NEI CLONI
#12:
12.4
Cloni genomici della regione di white a confronto
Clone corrispondente al gene white
pS12.20 clone dal ceppo M
Elemento di 2,9 kb
(ELEMENTO P)
p12.2 clone da ceppo 2
(in plasmide)
12 clone da ceppo 2
(in fago )
25 clone da ceppo 2
(in fago )
Elemento di 2,9 kb
(ELEMENTO P)
p25.1clone da ceppo 2
(in plasmide)
pS25.1 clone dal ceppo M
I frammenti più lunghi, di 2.9 kb (elementi P), sono stati subclonati in plasmidi pBR322 e chiamati p 12.2 e p 25.1
Caratteristiche dell’elemento P
751 amminoacidi
TRASPOSASI ATTIVA
Traduzione
AAAAA mRNA di 2400 nt
Trascrizione nel tessuto GERMINALE
+ processing dell’ RNA
Esone 1
Esone 2
Repeat
di 31 bp
Esone 3
Esone 4
Trascrizione nel tessuto SOMATICO
+ processing dell’ RNA
UAA
TRASPOSASI NON ATTIVA
Elemento P completo
2907 bp
Repeat
di 31 bp
AAAAA mRNA di 2500 nt
576 amminoacidi
REPRESSORE
La trasposizione di P è regolata GENETICAMENTE -> avviene solo nell’incrocio maschi P x femmine M
È anche regolata in modo TESSUTO-SPECIFICO -> avviene solo nelle cellule germinali
Regolazione dello splicing alternativo nell’elemento P
Nelle cellule SOMATICHE non avviene lo splicing del terzo introne a causa del legame a
questo livello del mRNA con la proteina PSI (P-element Somatic Inibitory protein) ed
altri elementi come RNA U1 ed una proteina ubiquitaria HRP48. La mancata eliminazione
dell’introne determina la produzione di un mRNA che codifica per il REPRESSORE
Elemento P completo
Nelle cellule GERMINALI manca la proteina PSI e quindi avviene lo splicing del terzo
introne, che determina la produzione del mRNA per la TRASPOSASI
Caratteristiche degli elementi P
• L’eterogeneità trovate negli elementi P che si inseriscono nei geni a seguito della HD è
dovuta a delezioni dell’elemento
• Se la delezione è INTERNA all’elemento, ma lascia intatte le 31 bp, questo elemento si
potrà ancora muovere ma non sarà in grado di produrre la TRASPOSASI
• Se la delezione interessa anche solo una delle due ripetizioni di 31 bp, l’elemento P
non sarà più in grado di muoversi
• Le inserzioni di elementi P causano sempre una duplicazione di 8 nucleotidi nella
sequenza in cui si inserisce
• Il numero di elementi P varia in ceppi diversi di Drosophila
Diffusione degli elementi P nelle popolazioni di Drosophila
I ceppi P inizialmente identificati erano tutti provenienti da popolazioni
naturali
I ceppi di laboratorio erano tutti ceppi M
Moscerini catturati prima del 1950 (che costituivano le maggiori
collezioni presenti nei laboratori, a cominciare dai ceppi che aveva
raccolto Morgan), erano privi di elementi P
C’è stata quindi una progressiva invasione nelle popolazioni naturali di
elementi P che è cominciata dal Nord e Sud America e dall’Africa ->
Giappone -> Australia -> Europa
Potrebbero essere stati portati da virus che infettano Drosophila
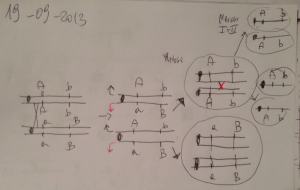
Definizione di CITOTIPO
Si definisce CITOTIPO una condizione cellulare, trasmessa per via materna
attraverso il citoplasma dell’uovo
Il CITOTIPO P è in grado di REPRIMERE l’attività degli elementi P
La madre P produce il repressore P nelle cellule follicolari somatiche e lo introduce
nell’uovo. Per cui non si verifica DISGENESIA nei seguenti incroci
Px
P
Px
M
Il CITOTIPO M è SENSIBILE all’attività degli elementi P, in quanto manca il REPRESSORE
INCROCIO NON DISGENICO
P
Citotipo P
X
Citotipo M
M
INCROCIO DISGENICO
M
Citotipo M
X
Citotipo P
Gli elementi P
nel citotipo P
sono repressi
Gli elementi P
nel citotipo M
sono ATTIVI
Progenie F1 normale
Progenie F1 DISGENICA
P
Classificazione dei ceppi di Drosophila in base al sistema PM
Ceppi P -> mostrano attività legata agli elementi P nel loro genoma ed
inducono HD; hanno citotipo P, sono cioè insensibili all’attività
degli elementi P
Ceppi M -> non hanno attività legata agli elementi P perchè nel loro genoma
non sono presenti; hanno citotipo M, sono cioè sensibili all’attività
degli elementi P
Ceppi M’ -> hanno elementi P NON FUNZIONALI, non inducono HD in quanto
non producono né trasposasi nè repressore; hanno citotipo M, sono
cioè sensibili all’attività degli elementi P e quindi in un
incrocio
M’ x
P si ha disgenesia
Ceppi Q-> hanno elementi P, citotipo P (hanno il repressore), ma non producono
la trasposasi e quindi non inducono HD infatti
M x Q non si ha disgenesia (non hanno la TRASPOSASI)
Qx
P non si ha disgenesia (hanno il REPRESSORE)
Trasferimento genico mediato dal DNA
Utilizzo del trasferimento genico mediato dal DNA:
• Funzionalità genica (GENOMICA FUNZIONALE)
• Funzionalità ed individuazione di sequenze importanti per l’espressione
in vivo di un gene
• Identificazione di NUOVE MUTAZIONI e clonaggio
• Si usa per ottenere organismi geneticamente modificati a vari scopi
Per OGM o animali transgenici si intendono animali ottenuti attraverso una manipolazione
del loro genoma inserendo uno o più geni appartenenti sia ALLA STESSA SPECIE che A
SPECIE DIFFERENTI
Nel caso di inserimento di geni della STESSA SPECIE si tende a
MIGLIORARE una capacità dell’animale nell’ambito zootecnico
Nel caso di inserimento di geni di SPECIE DIFFERENTI si crea un animale a
scopi sperimentali (animale MODELLO) oppure a scopi terapeutici
(xenotrapianti)
I vettori per il trasferimento genico devono:
- favorire l’integrazione di sequenze nel genoma
- devono essere facilmente selezionabili
In Drosophila (animale MODELLO) uno dei primi elementi
ad essere preso in considerazione perché funzionasse come
vettore per l’inserimento di sequenze nel genoma è stato
l’elemento P