Principi di genetica - Robert J. Brooker
Copyright © 2010 – The McGraw-Hill Companies srl
SOLUZIONI AI PROBLEMI DEL CAPITOLO 24
Domande concettuali
C1. Un pool genico è costituito da tutti i geni presenti in una particolare popolazione. Ogni tipo di gene nel pool
genico può esistere in una o più forme alleliche. La prevalenza di un allele all'interno del pool genico viene
descritta dalla sua frequenza allelica. Se un gene è monomorfico, la frequenza allelica è prossima al 100%. Se è
polimorfico, ogni allele ha una frequenza compresa tra 1 e 99%. La somma di tutte le frequenze alleliche per un
particolare gene equivale a 100%.
C2. Una popolazione è un gruppo di individui che si incrociano tra loro. Consideriamo una popolazione di scoiattoli
in una foresta. Nel corso di molte generazioni, potrebbero succedere diverse cose a questa popolazione. Un
incendio nella foresta, per esempio, potrebbe diminuire drasticamente il numero degli individui e perciò causare
un collo di bottiglia. Questo diminuirebbe la diversità genetica della popolazione. Un nuovo predatore può entrare
nella regione e la selezione naturale potrebbe favorire gli scoiattoli che sono più abili nello sfuggire al predatore.
Un’altra possibilità è che un gruppo di scoiattoli nella popolazione migri in una nuova regione dove fonda una
nuova popolazione.
C3. Il termine polimorfismo genetico si riferisce al fenomeno per il quale un gene è presente nella popolazione in due
o più forme alleliche. La mutazione è la fonte della variabilità genetica.
C4.
A. Frequenza fenotipica e frequenza genotipica.
B. Frequenza genotipica.
C. Frequenza allelica.
C5. Quando un carattere è polimorfico, ciò significa che individui diversi mostrano variabilità fenotipica rispetto a
questo carattere. Per esempio, le petunie possono avere fiori rossi oppure bianchi. Il colore dei fiori è polimorfico
nelle petunie. Quando un gene è polimorfico, esiste in due o più forme alleliche. A livello molecolare, gli alleli di
un determinato gene hanno sequenze di DNA diverse. Queste differenze potrebbero essere minime (per esempio
una variazione di una singola base) oppure potrebbero coinvolgere inserzioni o delezioni importanti. Alleli diversi
possono causare differenze a livello fenotipico. Per esempio, nelle piante di pisello di Mendel, l'allele T
determinava piante alte, mentre l'allele t causava piante nane. Tuttavia, non sempre gli alleli determinano
differenze fenotipiche. Una sostituzione di una singola base in un gene può non influenzare la sequenza
aminoacidica del polipeptide codificato (per esempio la base variata potrebbe essere la base vacillante) oppure un
cambiamento di una sola base potrebbe alterare la sequenza aminoacidica ma non in modo da alterare la funzione
proteica. Perciò, il polimorfismo genetico non sempre determina un polimorfismo fenotipico.
C6.
A. La frequenza genotipica dell’omozigote malato è 1/2500, ossia 0,004. Questo equivale a q2. La frequenza
allelica corrisponde alla radice quadrata di questo valore, che equivale a 0,02. La frequenza del corrispondente
allele sano corrisponde a 1 – 0,02 = 0,98.
B. La frequenza dell’omozigote malato è 0,004; per l’omozigote sano, (0,98)2 = 0,96; e per l’eterozigote, 2 (0,98)
(0,02), che equivale a 0,039.
C. Se è noto che una persona è portatore eterozigote, le probabilità che questa particolare persona scelga un altro
portatore sano come partner sessuale è uguale alla frequenza dei portatori eterozigoti nella popolazione, che
equivale a 0,039, ossia al 3,9%. Le possibilità di un incrocio casuale tra due individui eterozigoti della
popolazione è 0,039 x 0,039 = 0,0015, ovvero 0,15%.
C7. In assenza di altre forze, l'inbreeding non altera le frequenze alleliche, perché non favorisce la trasmissione di un
allele a discapito di un altro. Esso semplicemente aumenta la probabilità di omozigosi (che è una frequenza
genotipica) Nelle popolazioni umane, l'inincrocio incrementa la frequenza degli individui omozigoti per gli alleli
patologici rari e recessivi .
C8. Per due alleli, la classe eterozigote è al picco massimo quando essi hanno frequenza 0,5 ciascuno. Per tre alleli, i
due eterozigoti sono a livello massimo di frequenza quando quella di ciascun allele è 0,33.
Principi di genetica - Robert J. Brooker
Copyright © 2010 – The McGraw-Hill Companies srl
C9. Se applichiamo l'equazione di Hardy-Weinberg:
BB = (0,67)2 = 0,45 ovvero 45%
Bb = 2(0,67)(0,33) = 0,44 ossia 44%
bb = (0,33)2 = 0,11 ovvero 11%.
I dati ottenuti mostrano una percentuale maggiore di omozigoti (confronta 45% con 50% e 11% con 13%) e una
percentuale minore di eterozigoti (confronta 44 % con 37%) rispetto all'atteso. Perciò, questi dati sono in accordo
con l'inincrocio, che aumenta la percentuale degli omozigoti e diminuisce la percentuale degli eterozigoti.
C10.
Siccome questo è un carattere recessivo, solo gli omozigoti per questo allele saranno in grado di arrotolare la
lingua. Se p corrisponde all’allele che detemina la capacità di arrotolare la lingua e q equivale all’allele che non
consente l’arrotolamento, l’equazione di Hardy-Weinberg predice che la frequenza degli omozigoti in grado di
arrotolare la lingua sarebbe p2. In questo caso, p2 = (0,6)2 = 0,36, ossia 36%.
C11.
A. Sì
B. I progenitori comuni sono I-1 e I-2.
C. F = ∑ [(1/2)n (1 + FA)]
F = (½)9 + (½)9
F = 1/512 + 1/512 = 2/512 = 0,0039
D. Non possiamo affermarlo.
C12.
A. Il coefficiente di inincrocio viene calcolato usando la formula
F = ∑ [(1/2)n (1 + FA)]
In questo caso, c’è un progenitore comune, I-2. Siccome non conosciamo la storia precedente dell’individuo I2, assumiamo che essa non sia inincrociata, il che rende FA uguale a zero. Il percorso di inincrocio, per IV-3
contiene cinque persone, III-4, II-2, I-2, II-5, e III-5. Perciò n = 5.
F = (1/2)5 (1 + 0) = 1/32 = 0,031.
B. Sulla base dei dati mostrati in questo pedigree, l’individuo IV-4 non deriva da inincrocio.
C13.
A. Il coefficiente di inincrocio viene calcolato usando la formula
F = ∑ [(1/2)n (1 + FA)]
In questo caso ci sono due progenitori comuni, I-1 e I-2. Siccome non conosciamo la storia precedente di I-1 e
I-2, assumiamo che non fossero consanguinei, quindi FA = 0. I due percorsi di consanguineità per IV-2
prevedono cinque persone, III-4, II-2, I-1, II-6, e III-5; III-4, II-2, I-2, II-6, e III-5. Perciò, n = 5 per entrambi i
percorsi.
F = (½)5 (1 + 0) + (½)5 (1 + 0)
F = 0,031 + 0,031 = 0,062
B. Sulla base dei dati descritti in questo pedigree, l’individuo III-4 non è inincrociato.
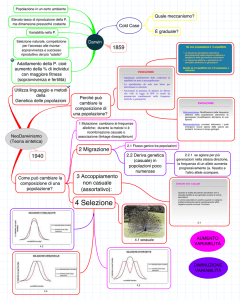
C14.
La migrazione, la deriva genetica, e la selezione naturale sono forze evolutive che alterano le frequenze
alleliche in una popolazione. La selezione naturale opera nell’eliminare gli alleli dannosi e promuove gli alleli
favorevoli. La deriva genetica comporta dei cambiamenti casuali nelle frequenze alleliche che possono infine
portare all’eliminazione oppure alla fissazione degli alleli. Si pensa sia importante nel mantenere gli alleli neutrali
in una popolazione. La migrazione è importante perché essa introduce nuove forme alleliche nelle popolazioni
vicine. Secondo la teoria neutrale, la deriva genetica è la principale responsabile della variabilità riscontrata nelle
popolazioni naturali.
C15. Nella deriva genetica, sono le frequenze alleliche a derivare. Deriva genetica è un'espressione appropriata perché
la parola deriva implica un processo casuale. Ciononostante, la deriva può essere direzionale. Una barca può
andare alla deriva tra le sponde di un lago. Essa non fluttuerà in un percorso lineare, ma il processo di deriva
Principi di genetica - Robert J. Brooker
Copyright © 2010 – The McGraw-Hill Companies srl
altererà la sua posizione. In modo simile, le frequenze alleliche possono aumentare oppure diminuire e infine
portare all'eliminazione oppure alla fissazione di particolari alleli all'interno di una popolazione.
C16.
Una forza casuale altera le frequenze alleliche a prescindere dal fatto che i cambiamenti siano o meno
favorevoli. La deriva genetica e la migrazione sono i due modi principali con cui questo può verificarsi. Un
esempio è l’effetto del fondatore, quando un gruppo di individui migra in una nuova localizzazione, come in
un’isola. Le forze adattative aumentano il successo riproduttivo di una specie. La selezione naturale è una forza
adattativa, che tende a eliminare gli alleli dannosi da una popolazione, e ad aumentare le frequenze degli alleli
favorevoli. Un esempio potrebbe essere il collo lungo delle giraffe, che le rende in grado di nutrirsi in presenza di
alberi alti. A livello molecolare, le mutazioni vantaggiose possono alterare la sequenza codificante di un gene e
cambiare la struttura e la funzione della proteina verso una forma favorevole. Per esempio, l’allele per l’anemia
falciforme altera la struttura dell’emoglobina, e in eterozigosi, questa inibisce la sensibilità dei globuli rossi al
patogeno della malaria.
C17. Possiamo utilizzare la seguente equazione per calcolare la variazione della frequenza allelica dopo un qualsiasi
numero di generazioni:
(1 – μ)t = pt / p0
(1 – 10-4)t = 0,5/ 0,6 = 0,833
(0,9999)t = 0,833
t = 1827 generazioni
C18.
La deriva genetica è dovuta all’errore di campionamento, e l’entità dell’errore di campionamento dipende
dalla dimensione della popolazione. Nelle popolazioni piccole, la proporzione relativa degli errori di
campionamento è grande. Se la deriva genetica sta spingendo un allele verso la fissazione, ciò richiederà più tempo
se la popolazione è grande, perché l’entità degli errori di campionamento è inferiore.
C19.
A. Probabilità di fissazione = 1/2N = ½(4) = 1/8, ovvero 0,125
B. t = 4N = 4(4) = 16
C. I calcoli precedenti assumono che la dimensione della popolazione sia costante. Se la popolazione crescesse
dopo essere stata fondata da questi quattro individui, la probabilità di fissazione sarebbe inferiore e il tempo
necessario a raggiungere la fissazione maggiore.
C20.
Durante l’effetto del collo di bottiglia, le frequenze alleliche sono drasticamente alterate a causa della deriva
genetica. In casi estremi, alcuni alleli vengono persi, mentre altri possono essere fissati al 100%. L’effetto
generale è la diminuzione della diversità genetica nella popolazione. Questo rende più difficile che le specie
rispondano in modo positivo a cambiamenti ambientali. Anche le specie prossime all’estinzione affrontano un
collo di bottiglia quando il numero di individui diminuisce. La perdita della diversità genetica può rendere
ancora più improbabile che la specie superi il momento negativo.
C21. Quando due popolazioni si mescolano, entrambe le popolazioni tendono ad avere una maggiore variabilità
genetica perché ogni popolazione introduce nuovi alleli nell'altra popolazione. Inoltre, le due popolazioni tendono
ad avere frequenze alleliche simili, soprattutto quando migra una grande proporzione di individui.
C22. In tutti i casi, queste forme di selezione naturale favoriscono uno o più fenotipi perché questi hanno un vantaggio
riproduttivo. Tuttavia, le modalità differiscono a seconda che siano favoriti un singolo fenotipo o diversi
fenotipi,oppure a seconda che il fenotipo favorito sia intermedio nell’intervallo di fenotipi o corrisponda a uno o
entrambi gli estremi. La selezione direzionale favorisce un fenotipo a un estremo fenotipico. Nel tempo, la
selezione naturale favorirà la fissazione degli alleli che determinano queste caratteristiche fenotipiche. La
selezione diversificante favorisce due o più categorie fenotipiche in base ai diversi ambienti occupati dalla specie.
Esempi di selezione bilanciante sono il vantaggio dell’eterozigote e la selezione dipendente negativamente dalla
frequenza. Queste promuovono un polimorfismo stabile in una popolazione. La selezione stabilizzante favorisce
gli individui con fenotipi intermedi. Essa tende a diminuire la diversità genetica perché gli alleli che favoriscono i
fenotipi estremi vengono eliminati.
C23. La fitness darwiniana è la probabilità relativa che un genotipo sopravvivrà e contribuirà al pool genico della
generazione successiva rispetto ad altri genotipi. Al genotipo con la maggiore capacità riproduttiva viene
Principi di genetica - Robert J. Brooker
Copyright © 2010 – The McGraw-Hill Companies srl
attribuito il valore 1,0. Le caratteristiche che promuovono la sopravvivenza, la capacità di attrarre un partner,
oppure la maggiore fertilità tendono a promuovere la fitness darwiniana. Esempi sono il folto mantello dell'orso
polare, che lo aiuta a sopravvivere nel clima freddo; il piumaggio appariscente degli uccelli maschi, che serve
loro per attirare le femmine; e l'elevato numero di gameti rilasciati da alcune specie di pesci, che incrementa la
loro fertilità.
C24. Il significato intuitivo della fitness media della popolazione è la probabilità relativa con la quale i membri di
una popolazione si riprodurranno. Se la fitness media è alta, è probabile che mediamente i membri della
popolazione sopravvivano, dando origine a della progenie. La selezione naturale aumenta la fitness media di una
popolazione.
C25.
A. La mutazione casuale è la fonte della variabilità genetica che può portare alla resistenza agli antibiotici. Una
mutazione casuale può creare un allele per la resistenza agli antibiotici. Questo potrebbe avvenire in diversi
modi. Due possibilità sono:
1.
Molti antibiotici esercitano i loro effetti legandosi a una proteina cellulare fondamentale all'interno
del microrganismo e inibendone la funzione. Una mutazione casuale potrebbe avvenire nel gene che
codifica questa proteina cellulare essenziale; questo potrebbe alterare la struttura della proteina in
modo da impedire all'antibiotico di legarsi alla proteina e inibirne la funzione.
2.
I microrganismi che vengono uccisi dagli antibiotici possiedono molti enzimi, che degradano
composti simili. Una mutazione casuale potrebbe alterare un gene che codifica un enzima nella con
capacità di degradazione, in modo che l'enzima ora riconosca l'antibiotico e lo degradi.
B. Quando si verificano nuove mutazioni casuali, esse possono essere perse attraverso la deriva genetica. Questo
è particolarmente probabile quando la frequenza della mutazione è molto bassa in una grande popolazione. In
alternativa (e con minore probabilità), una mutazione casuale che conferisce la resistenza agli antibiotici
potrebbe venire fissata in una popolazione.
C. Se si verifica una mutazione che conferisce la resistenza agli antibiotici, e se la mutazione non viene persa a
causa della deriva genetica, la selezione naturale favorirà la crescita dei microrganismi che portano l'allele per
la resistenza agli antibiotici quando gli organismi vengono esposti agli antibiotici. Perciò, se gli antibiotici
sono usati in modo diffuso, questi uccideranno i microrganismi sensibili e favoriranno la proliferazione di
quelli che portano gli alleli per la resistenza agli antibiotici.
C26.
A. Vero
B. Vero
C. Falso, essa causa la perdita o la fissazione dell’allele, che determina minore diversità genetica.
D. Vero
C27.
A. La migrazione aumenterà la diversità genetica in entrambe le popolazioni. Una mutazione casuale potrebbe
verificarsi in una popolazione creando un nuovo allele. Questo nuovo allele potrebbe essere introdotto
nell'altra popolazione attraverso la migrazione.
B. Le frequenze alleliche nelle due popolazioni tenderanno a essere simili tra loro, perché i loro alleli si
mescolano.
C. La deriva genetica dipende dalle dimensioni delle popolazioni. Quando le due popolazioni si mescolano,
questo porta a un aumento delle dimensioni della popolazione complessiva. In un certo senso, le due
popolazioni più piccole si comportano come una grande popolazione. Perciò, gli effetti della deriva genetica
sono attenuati quando gli individui delle due popolazioni possono migrare. L'effetto netto è che la perdita
oppure la fissazione degli alleli sono meno probabili.
C28.
A. Diversificante. Esistono diversi ambienti che favoriscono fenotipi diversi.
B. Direzionale. Più folta è la pelliccia, maggiore è la probabilità di sopravvivenza.
Principi di genetica - Robert J. Brooker
Copyright © 2010 – The McGraw-Hill Companies srl
C. Stabilizzante. Il basso peso alla nascita viene selezionato a sfavore perché esso determina una bassa
sopravvivenza. Inoltre, anche gli elevati pesi alla nascita vengono selezionati a sfavore perché potrebbero
determinare problemi nel parto, i quali potrebbe altrettanto diminuire il tasso di sopravvivenza.
D. Direzionale. Fusti e foglie forti promuoveranno la sopravvivenza nei climi ventosi.
Domande sperimentali
S1.
A. A livello del DNA, un modo sicuro per determinare la variabilità genetica è quello di clonare e sequenziare i
geni. Se lo stesso gene viene clonato da due individui diversi e le sequenze sono diverse, questo dimostra che
vi è variabilità genetica. Inoltre, molti altri metodi possono essere usati per determinare la variabilità genetica.
Per esempio, un confronto dei Southern blot ottenuti utilizzando i campioni di individui diversi potrebbe
rivelare che un gene presenta dimensioni diverse oppure che esso possiede siti di restrizione diversi.
B. A livello dell'RNA, un Northern blot può indicare la variabilità genetica. Se l'RNA codificato da due diversi
alleli presenta dimensioni diverse, un Northern blot lo metterà in evidenza.
C. A livello delle proteine, l'elettroforesi su gel può indicare la variabilità genetica. Questo metodo è descritto
nell'Appendice sul sito web. Un altro approccio è lo studio della funzione di un enzima usando dei saggi di
attività enzimatica.
S2. Il problema risolto R4 descrive in quale modo l’equazione di Hardy-Weinberg possa essere modificata per
includere situazioni con tre o più alleli. In questo caso
(p + q + r + s)2 = 1
p2 + q2 + r2 + s2 + 2pq + 2qr + 2qs + 2rp + 2rs + 2sp = 1
Poniamo p =C, q =cch, r =ch, e s = c.
A. La frequenza dei conigli albini è
s2 = (0,05) = 0,0025 = 0,25%
B. Himalaiano è dominante su albino ma recessivo rispetto a uniforme e cincillà. Perciò, conigli con mantello
himalaiano sarebbero rappresentati da r2 e da 2rs:
r2 + 2rs = (0,44)2 + 2 (0,44) (0,05) = 0,24 = 24%
In 1000 individui, circa 240 avranno il mantello himalaiano.
C. Cincillà è dominante su himalaiano e albino ma recessivo sul mantello uniforme. Perciò, gli eterozigoti con
mantello cincillà saranno rappresentati da
2qr + 2qs = [2 (0,17) (0,44)] + [2 (0,17) (0,05)] = 0,17, ossia 17%
In 1000 individui, circa 170 avranno il mantello cincillà.
S3. Se W indica l'allele per il grasso bianco e w rappresenta l'allele per il grasso giallo, assumendo equilibrio di
Hardy-Weinberg, p2 rappresenterebbe la frequenza genotipica degli animali WW, Ww sarebbe 2pq e ww sarebbe
q2. L'unica frequenza genotipica che conosciamo è quella degli animali ww.
ww = q2 = 76/5468 = 0,014
q = 0,12 che è la frequenza dell'allele w
p = 1 – q = 0,88 che è la frequenza dell'allele W
A. I portatori eterozigoti sono rappresentati da 2pq. Se usiamo i valori di p e q, che sono stati calcolati nella
parte A:
2pq = 2(0,88)(0,12) = 0,21
Principi di genetica - Robert J. Brooker
Copyright © 2010 – The McGraw-Hill Companies srl
Approssimativamente il 21% degli animali sarebbe eterozigote con grasso bianco. Se moltiplichiamo 0,21 per
il numero totale degli animali nel gregge
0,21 x 5468 = 1148 animali
S4.
A. Inuit
M = 0,913
N = 0,087
Navaho
M = 0,917
N = 0,083
Finlandesi
M = 0,673
N = 0,327
Russi
M = 0,619
N = 0,381
Aborigeni
M = 0,176
N = 0,824
B. Per determinare se queste popolazioni sono in equilibrio, possiamo usare l’equazione di Hardy-Weinberg e
calcolare i valori attesi per ciascun genotipo. Per esempio,
Inuit
MM = (0,913)2 = 0,833 = 83,3%
MN = 2 (0,913) (0,087) = 0,159 = 15,9%
NN = (0,087)2 = 0,0076 = 0,76%
In generale, i valori sono abbastanza in accordo con l’equilibrio. Lo stesso è vero per le altre quattro
popolazioni.
C. Sulla base delle simili frequenze alleliche, gli Inuit e gli indiani Navaho sembrano inincrociati, così come i
finlandesi e i russi.
S5. La prima cosa da fare è quella di determinare le frequenze alleliche. p rappresenta i, q rappresenta IA, e r
rappresenta IB.
p2 è la frequenza genotipica di ii
q2 è la frequenza genotipica di IAIA,
r2 è la frequenza genotipica di IBIB,
2pq è la frequenza genotipica di IAi,
2pr è la frequenza genotipica di IBi
2qr è la frequenza genotipica di IAIB
p2 = 721 /(721 + 932 + 235 + 112) = 0,36
p = 0,6
Poi, possiamo calcolare la frequenza allelica di IA. Ricorda che vi sono due genotipi (IAIA e Iai) che determinano il
gruppo sanguigno A.
q2 + 2pq = 932 /(721 + 932 + 235 + 112) = q2 + 2(0,6)q = 0,47
Risolvendo questa equazione otteniamo
q = 0,31
Possiamo risolvere per r
p+q+r=1
0,6 + 0,31 + r = 1
r = 0,09
Sulla base di queste frequenze alleliche, possiamo confrontare i valori osservati e quelli attesi. Per determinare
i valori attesi, moltiplichiamo le frequenze genotipiche per 2000, che è il numero totale degli individui di questa
popolazione.
p2 è la frequenza genotipica di ii = (0,6)2(2000) = 720
q2 è la frequenza genotipica di IAIA = (0,31)2(2000) = 192
Principi di genetica - Robert J. Brooker
Copyright © 2010 – The McGraw-Hill Companies srl
r2 è la frequenza genotipica di IBIB = (0,09)2(2000) = 16
2pq è la frequenza genotipica di IAi = 2(0,31)(0,6)(2000) = 744
2pr è la frequenza genotipica di IBi = 2(0,09)(0,6)(2000) = 216
2qr è la frequenza genotipica di IAIB = 2(0,31)(0,09)(2000) = 111
INSERIRE LA TABELLA
Expected numbers = Numeri attesi
Observed numbers = numeri osservati
Type = gruppo
I valori osservati e quelli attesi sono in accordo tra loro. Perciò sembra che questa popolazione sia in equilibrio
di Hardy-Weinberg
S6.
A. ΔpC = m (pD – pR)
Rispetto all’allele per l’anemia falciforme,
ΔpC = (550/10 550) (0,1 – 0,01) = 0,0047
pC = pR + ΔpC = 0,01 + 0,0047 = 0,0147
B. Dobbiamo calcolare i genotipi separatamente:
Per i 550 individui migranti,
HbAHbA = (0,9)2 = 0,81, ossia 81%
Sono attesi (0,81) 550 = 445,5 individui con questo genotipo.
HbAHbS = 2 (0,9) (0,1) = 0,18
Sono attesi (0,18) 550 = 99 eterozigoti.
HbSHbS = (0,1)2 = 0,011
Sono attesi (0,01) 550 = 5,5 individui HbSHbS.
Per la popolazione ricevente originale,
HbAHbA = (0,99)2 = 0,98
Sono attesi 9801 individui con questo genotipo.
HbAHbS = 2 (0,99) (0,01) = 0,0198
Sono attesi 198 individui con questo genotipo.
HbSHbS = (0,01)2 = 0,0001
È atteso 1 individuo con questo genotipo.
Per calcolare la popolazione totale,
(445,5 + 9801) / 10 550 = 0,971 omozigoti HbAHbA
(99 + 198) / 10 550 = 0,028 eterozigoti
(5,5 + 1) / 10 550 = 0,00062 omozigoti HbSHbS
Principi di genetica - Robert J. Brooker
Copyright © 2010 – The McGraw-Hill Companies srl
C. Dopo un ciclo di accoppiamento, le frequenze alleliche nel conglomerato (calcolate nella parte A), dovrebbero
produrre le frequenze genotipiche attese secondo l’equilibrio di Hardy-Weinberg.
Frequenza allelica di HbS = 0,0147, e HbA = 0,985.
HbAHbA = (0,985)2 = 0,97
HbAHbS = 2 (0,985) (0,0147) = 0,029
HbSHbS = (0,0147)2 = 0,0002
S7. Assumiamo che i valori di fitness relativa siano 1,0 per l'omozigote dominante e l'eterozigote e 0 per l'omozigote
recessivo. La prima cosa che dobbiamo fare è calcolare la fitness media della popolazione.
p2WAA + 2pqWAa + q2Waa = W
(0,78)2 + 2(0,78)(0,22) = W
W = 0,95
La frequenza allelica (p) nella generazione successiva è
p = p2WAA / W + pqWAa / W
p = 0,82
q = 1 – p = 0,18
Dovremmo seguire la stessa strategia generale per la seconda e la terza generazione.
Per la seconda generazione, la fitness media della popolazione equivale a 0,97. Usando la precedente equazione,
la frequenza allelica di A nella seconda generazione equivale a circa 0,85. La frequenza dell'allele recessivo nella
seconda generazione equivale a circa 0,15 e la fitness media sarebbe ora di circa 0,98. La frequenza allelica di A
nella terza generazione equivale a circa 0,87. La frequenza dell'allele recessivo nella seconda generazione
equivale a circa 0,15 e la fitness media sarebbe ora di circa 0,13.
S8.
A. La probabilità di fissazione = 1/2N (assumendo che lo stesso numero di maschi e femmine contribuisca alla
generazione successiva)
Probabilità di fissazione = ½ (2 000 000)
= 1 su 4 000 000 di possibilità
B. t = 4N
dove t = numero medio di generazioni per raggiungere la fissazione
N = numero di individui nella popolazione, assumendo che maschi e femmine contribuiscano equamente alla
generazione successiva
t = 4 (2 milioni) = 8 milioni di generazioni
C. Se l’allele per il colore blu offre un vantaggio selettivo, il valore calcolato nella parte A sarebbe leggermente
maggiore: ci sarebbe una possibilità maggiore di fissazione allelica. Il valore calcolato nella parte B sarebbe
inferiore; impiegherebbe meno tempo per raggiungere la fissazione.
S9. I coefficienti di selezione sono
sww = 1 – 0,19 = 0,81
sWW = 1 – 0,37 = 0,63
Se i ratti fossero esposti alla warfarina, non vi sarebbe più equilibrio, e la selezione naturale tenderebbe a
eliminare l'allele della resistenza perché gli omozigoti mancano della vitamina K.
S10. Potresti marcare le chiocciole con un colore e rilasciare un ugual numero di chiocciole scure e chiare in una
Principi di genetica - Robert J. Brooker
Copyright © 2010 – The McGraw-Hill Companies srl
regione con una foresta scarsamente illuminata. Successivamente, catturare le chiocciole e contarle. Sarebbe
importante avere un metodo di cattura obiettivo perché lo sperimentatore raccoglierà più facilmente le chiocciole
chiare nella foresta e quelle scure nei campi aperti. Forse potresti usare come esca qualcosa di gradevole per le
chiocciole e raccogliere solo quelle che si trovano vicino all’esca. Inoltre, con questo tipo di esperimento si
potrebbe anche osservare se avviene predazione.
S11. Questo è un esempio di selezione direzionale. A lungo andare, la selezione direzionale può portare alla perdita di
alcuni alleli e alla fissazione di altri. In questo caso, gli alleli che promuovono la dimensione più piccola del
becco possono venire persi dalla popolazione, mentre gli alleli che promuovono la dimensione più grande
potrebbero essere fissati.
S12. Il maschio 2 è il padre potenziale, perché esso possiede le bande osservate nel figlio ma non nella madre. Per
calcolare la probabilità, dovresti conoscere la possibilità di osservare ciascun tipo delle bande di cui si ha
corrispondenza. Per esempio, in questo caso, il maschio 2 e il figlio hanno quattro bande in comune. Come
semplice calcolo, potremmo eliminare le quattro bande che il figlio condivide con la madre. Se la probabilità di
avere ciascuna delle bande paterne è ¼, la probabilità che questa persona non sia il padre è (1/4)4.
S13. Il DNA fingerprinting è un metodo di identificazione basato sulle proprietà del DNA. Le sequenze dei
minisatelliti e dei microsatelliti sono variabili rispetto alla dimensione delle popolazioni naturali. Questa
variabilità può essere vista quando i frammenti di DNA sono sottoposti all'elettroforesi su gel. All'interno di una
popolazione, qualsiasi coppia di individui (eccetto i gemelli monozigotici) differirà per il tipo di frammenti di
DNA, che rappresenta il DNA fingerprint o profilo genetico.
S14. La PCR viene utilizzata per amplificare il DNA se ne è presente solamente una piccola quantità (per esempio un
piccolo campione trovato nella scena di un crimine). Essa viene anche utilizzata per amplificare i microsatelliti. Il
Southern blotting, utilizzando una sonda complementare al minisatellite, è necessario per identificare in modo
specifico un limitato numero di bande (circa 20) che sono variabili nelle popolazioni umane.
S15. Un minisatellite è una sequenza ripetuta numerose volte all'interno di un genoma ed è variabile nella sua
lunghezza. Nelle popolazioni naturali, è comune trovare variabilità di lunghezza. Perciò, qualsiasi coppia di
individui che non siano geneticamente identici differirà rispetto alle dimensioni di molti dei loro minisatelliti:
Quando si esegue un’elettroforesi su gel e i minisatelliti vengono evidenziati in un Southern blot, le dimensioni
dei minisatelliti costituiscono il profilo genetico (DNA fingerprint) dell'individuo. Questo profilo è una
caratteristica unica di ciascun individuo.
S16. Questa percentuale non è troppo alta. Sulla base della loro relazione genetica, ci aspettiamo che un padre e una
figlia debbano condividere almeno il 50% delle stesse bande nell’impronta digitale del DNA. Tuttavia, il valore
può essere maggiore a quello perché la madre e il padre possono avere delle bande in comune, anche se non sono
consanguinei. Per esempio, in un locus genomico, il padre può essere eterozigote per un microsatellite di 4100 bp
e 5200 bp, e la madre può essere eterozigote con microsatelliti di 4100 bp e 4700 bp. Il padre potrebbe
trasmettere la banda da 5200 bp alla figlia, e la madre potrebbe trasmettere quella da 4100 bp. Questo darebbe
una perfetta coincidenza con entrambe le bande del padre, anche le il padre ha trasmesso solamente la banda da
5200 bp alla figlia. La banda da 4100 bp si osserva perché il padre e la madre hanno un microsatellite in comune.
Perciò, la stima del 50% di corrispondenza tra le bande nel profilo genetico di individui consanguinei è una stima
minima. Il valore può essere superiore.
S17. La percentuale minima di corrispondenza di bande si basa sulle relazioni genetiche.
A. 50%
B. 50% (in media, occasionalmente anche meno)
C. 25% (in media, occasionalmente anche meno)
D. 25% (in media, occasionalmente anche meno)