Divisione Magnoliophyta o Angiospermae
Classificazione scientifica
Dominio: Eucariota (Eukaryota o Eukarya/Eucarioti)
Regno: Plantae (Plants/Piante)
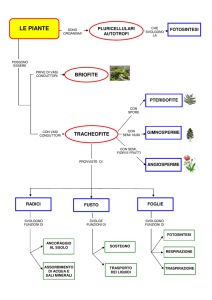
Sottoregno: Tracheobionta (Vascular plants/Piante vascolari)
Superdivisione: Spermatophyta (Seed plants/Piante con semi)
Divisione: Magnoliophyta o Angiospermae
Alla divisione Magnoliophyta o Angiospermae appartengono, classicamente, due grandi classi:
• Dicotiledoni o Magnoliopsida
• Monocotiledoni o Liliopsida
Le Magnoliofite (o Magnoliophyta) sono una vasta divisione di piante note anche sotto il nome di
Angiosperme (o Angiospermae). Il Codice Internazionale di Nomenclatura Botanica (ICBN)
permette, per i taxa di rango superiore alla famiglia, di usare indipendentemente nomi descrittivi
tradizionali come Angiospermae o regolarmente derivati da un genere "tipo" come Magnoliophyta,
che deriva dal genere Magnolia.
Apparse nel Cretaceo (circa 130 milioni di anni fa), le angiosperme hanno rapidamente soppiantato
le felci e le gimnosperme.
Fanno parte della divisione delle Magnoliofite circa 300.000 specie botaniche, erbacee, arboree,
arbustive e lianose. Sono oggi le piante più diffuse ed importanti, sia dal punto di vista ecologico
che economico.
La sistematica delle Angiosperme
Secondo una visione tradizionale della sistematica basata essenzialmente sui caratteri
morfologici, le angiosperme venivano divise nelle due classi delle monocotiledoni (Liliopsida) e
dicotiledoni (Magnoliopsida). All’interno di queste ultime, venivano individuate in passato un
piccolo numero di sottoclassi sulla base delle caratteristiche del perianzio: piante con fiori privi sia
di calice che di corolla oppure provvisti di un solo verticillo; piante a fiori con corolla dialipetala;
piante a fiori con petali fusi in corolla gamopetala. Tra queste sottoclassi veniva individuata una
linea evolutiva che andava dal perianzio ridotto o assente, alla corolla a elementi liberi, a quella
gamopetala. Sulla base di un’impostazione sistematica di questo tipo, tutte le angiosperme della
flora mondiale vennero inquadrate dal sistematico tedesco Engler in uno schema tassonomico molto
articolato che arrivava fino al rango di genere (Engler & Prantl Die Natürlichen Pflanzenfamilien,
1887-1915). Partendo dall’ordinamento di Engler, famiglie e generi sono stati ordinati in una
sequenza sistematica su presunte basi filogenetiche, dal genere ritenuto meno evoluto a quello
ritenuto più evoluto. Schematizzando molto, nelle dicotiledoni si andava dalle cosiddette amentifere
– piante arboree con fiori anemogami a perianzio nullo o ridotto riuniti in amenti di famiglie come
Salicaceae, Betulaceae, Fagaceae – fino alle più complesse composite (Asteraceae), con fiori a
corolla gamopetala riuniti in infiorescenze specializzate a capolino. Nelle monocotiledoni, da
famiglie con fiori regolari come Alismataceae, Juncaceae, Liliaceae, alle Orchidaceae con fiori
irregolari complessi.
Questa impostazione sistematica ha goduto a lungo di un vasto seguito in gran parte d’Europa.
Sequenze di famiglie e generi basate su un simile schema di riferimento sono tuttora alla base
dell’ordinamento seguito in flore autorevoli anche recenti (Flora Europaea, Flora d’Italia di
Pignatti) e nelle collezioni di molti erbari.
Tuttavia, indagini sistematiche condotte nell’ultimo mezzo secolo utilizzando altri dati oltre a
quelli morfologici (dati anatomici, cromosomici e biosistematici in genere, dati biochimici) hanno
messo in luce legami filogenetici e linee evolutive molto diversi da quelli ipotizzati da Engler e in
particolare si è riconosciuto il carattere in genere derivato dei fiori anemofili rispetto a quelli
entomofili. Sulla base di queste nuove conoscenze sono stati proposti vari schemi di classificazione
delle piante a fiore di impostazione più rigorosamente filogenetica, in molte delle quali scompare la
stessa distinzione tra monocotiledoni e dicotiledoni.
Tra questi, uno dei più accettati è stato a lungo quello di Arthur Cronquist (in seguito integrato e
modificato da altri autori), in cui le angiosperme più primitive non sono più ritenute le amentifere,
ma quelle a fiori con elementi numerosi disposti a spirale, del tipo delle Magnoliaceae,
Nymphaeaceae, Ranunculaceae, ecc. In questo schema (figura1), le piante a fiore sono divise in una
decina di sottoclassi, il cui numero e la cui delimitazione hanno subìto via via numerose modifiche,
col progredire delle conoscenze.
Figura 1 – Una classificazione delle Magnoliophyta in 11 sottoclassi, basata sul sistema di
Cronquist.
La classificazione di Cronquist è stata accettata da molti autori nei decenni passati, ma
attualmente è ritenuta anch’essa superata sulla base delle nuove conoscenze derivate
dall’analisi e confronto del DNA dei diversi taxa, attraverso la quale diventa possibile
ricostruire direttamente almeno in parte i legami filogenetici tra i diversi taxa. Questo tipo di
indagini è sfociato in un nuovo approccio strumentale alla sistematica (sistematica molecolare),
che ha portato a dare negli ultimi decenni nuovo impulso agli studi sulla filogenesi e
conseguentemente sulla tassonomia delle angiosperme. Le indagini sistematiche sulle
Magnoliophyta conoscono attualmente un momento di grande floridità, col risultato che nuovi
schemi tassonomici vengono proposti si può dire ogni anno.
Al momento, nessuno di questi ha trovato l’accordo della maggioranza degli studiosi ed è
accettato come risolutivo, tanto che in alcuni testi recenti le angiosperme vengono inquadrate in
generi, famiglie, ordini, ma al di sopra di questo rango sono riunite in gruppi provvisori senza
valore tassonomico.
In particolare, è stata messa fortemente in discussione la tradizionale distinzione delle
angiosperme nelle due classi delle monocotiledoni e delle dicotiledoni, secondo le regole della
nomenclatura botanica meglio denominate come classi Liliopsida e Magnoliopsida.
Inizialmente si è ritenuto che la separazione tra le due classi fosse avvenuta molto
precocemente, dal momento che tra i resti fossili del Cretaceo si possono riconoscere già forme
attribuibili all’una o all’altra e che alcune famiglie di angiosperme viventi che hanno
conservato caratteri di maggiore primitività appaiono difficilmente inquadrabili tra le
dicotiledoni o le monocotiledoni. I dati più recenti sembrerebbero invece dimostrare che solo le
monocotiledoni costituiscono un gruppo monofiletico, derivano cioè tutte da un unico antenato
comune, mentre le dicotiledoni comprenderebbero almeno due gruppi a origine indipendente:
- il grande gruppo delle “vere dicotiledoni” (o eu-dicotiledoni), caratterizzate da avere
polline con tre solchi (tricolpato). Questo gruppo comprende la maggior parte delle famiglie
tradizionalmente inquadrate nelle dicotiledoni;
- un gruppo molto più piccolo che include le famiglie che hanno mantenuto caratteri più
primitivi.
Questo secondo gruppo, che corrisponde grosso modo alla sottoclasse Magnoliidae della
classificazione di Cronquist, può essere ulteriormente suddiviso in due:
famiglie a portamento legnoso con foglie più o meno coriacee, come le Magnoliaceae
(gruppo delle “Magnoliidi arboree”);
famiglie erbacee a foglie sottili, come le Nymphaeaceae e le Aristolochiaceae, con
caratteristiche in parte simili a quelle delle monocotiledoni, di cui rappresenterebbero
il gruppo ancestrale (“Paleoerbe non monocotiledoni”). Si tratta di un gruppo
abbastanza eterogeneo.
Fig. 2 – Classificazione delle angiosperme su base filogenetica.
Tuttavia, per una sistematica elementare a carattere applicativo, la distinzione in monocotiledoni e
dicotiledoni appare ancora utile e ad essa verrà fatto riferimento anche nella trattazione delle
principali famiglie di interesse applicativo ed ecologico che è riportata più avanti.
Caratteri fondamentali
Le Magnoliofite si distinguono perché i loro semi sono avvolti da un frutto, che li protegge e ne
facilita la disseminazione. Il nome descrittivo Angiosperme significa infatti "seme protetto" (dal
Greco αγγειον, ricettacolo, e σπερµα, seme).
Il fiore delle Magnoliofite è più complesso di quello delle altre Divisioni delle Spermatofite (qui si
tratta di un semplice complesso di sporofilli senza strutture specializzate caratteristiche dei fiori
convenzionali), anche se tutte condividono la riproduzione per mezzo di semi (per questo motivo
tutte sono incluse nella Superdivisione delle Spermatophyta. Il fiore delle Magnoliofite é costituito
da elementi sterili (i sepali ed i petali: i primi che formano il calice ed i secondi la corolla) e da
elementi fertili, presenti o non nello stesso fiore: gli stami in cui si differenziano i granuli di
polline, e il carpello (o i carpelli): foglie modificate ripiegate su se stesse, in cui si distinguono,
dall'apice verso la base, lo stimma, lo stilo e l'ovario, che racchiude gli ovuli (Figura 3).
Quindi, nelle Angiosperme, gli ovuli non sono semplicemente "portati" da squame, come accade
nella ex Divisione delle Gimnosperme, ma sono racchiusi all'interno dell'ovario che, durante la
maturazione del seme, si trasforma nel pericarpo del frutto. Questa é una importante differenza
rispetto alle Gimnosperme (le piante "a semi nudi"): infatti il frutto é un organo esclusivo delle
Angiosperme, molto importante ai fini della dispersione dei semi ad opera del vento, dell'acqua o di
animali.
Il processo riproduttivo delle Angiosperme é simile a quello delle Gimnosperme, con alcune
differenze:
- il granulo pollinico completamente germinato (il gametofito maschile) contiene solo tre nuclei
anziché quattro come nelle Gimnosperme, una cellula vegetativa e due cellule spermatiche;
- il polline non viene disperso solamente dal vento, ma alla sua dispersione partecipano numerosi
animali, tra cui soprattutto gli insetti. Tuttavia, nel corso dell’evoluzione, molte specie di
Angiosperme sono tornate all'impollinazione ad opera di agenti non biologici, soprattutto il vento. I
fiori sono allora diventati piccoli, numerosi e poco appariscenti. L’impollinazione anemofila è
apparsa indipendentemente in numerosi gruppi di angiosperme sia monodicotiledoni (Poaceae) che
dicotiledoni (es. Salicaceae, Fagaceae);
- il polline non entra mai all'interno dell'ovulo ma aderisce alla superficie dello stimma, quindi, in
alcune specie, il tubetto pollinico deve allungarsi in modo considerevole attraverso lo stilo per
giungere agli ovuli;
- il gametofito femminile maturo, il sacco embrionale, é costituito solamente da 8 nuclei, e contiene
un'unica cellula uovo che non é racchiusa nell'archegonio;
- avviene il processo della doppia fecondazione: alla fecondazione partecipano entrambi i nuclei
spermatici del granulo pollinico: uno feconda la cellula uovo dando origine allo zigote, l'altro
invece si fonde con due nuclei (i nuclei polari) che si trovano al centro del sacco embrionale, e si
forma l'endosperma triploide che servirà come tessuto di riserva per lo sviluppo dell'embrione;
- gli embrioni delle Angiosperme possono avere uno o due cotiledoni (foglie embrionali),
rispettivavente nelle Mono- e nelle Dicotiledoni, che possono funzionare come organi di riserva o
come austori durante la germinazione dell'embrione (Figura 3).
Infine,l e Magnoliofite si distinguono anche per la presenza di trachee vere e proprie unite a canali
più grandi e specializzati, gli "elementi dei vasi", che rendono più efficiente il trasporto idrico, e per
essere l'unica divisione che comprende piante erbacee in senso proprio.
Nelle angiosperme, i processi riproduttivi raggiungono il massimo livello di perfezionamento e di
efficienza nel mondo vegetale, grazie a due fondamentali innovazioni premiate dalla selezione: il
carpello e il fiore.
Il carpello presenta una serie di notevoli vantaggi evolutivi:
- ulteriore protezione del gametofito femminile dal pericolo di disseccamento e di danneggiamenti;
- possibilità di aumentare il periodo di ricettività del polline, senza rischio di disseccamento per
l’ovulo;
- grazie all’allontanamento della superficie recettiva dall’oosfera, possibilità per la pianta di mettere
in atto meccanismi di incompatibilità sul percorso più lungo del tubetto pollinico, arrivando in
definitiva a scegliere il gamete maschile più “gradito” per la fecondazione;
- possibilità di contribuire attivamente alla dispersione del seme, con la trasformazione del carpello
in frutto dopo la fecondazione e le relative modificazioni in funzione dell’agente della dispersione
(frutti carnosi ingeriti dagli animali, frutti alati portati dal vento, ecc.).
Figura 3 – Fasi principali del ciclo gametofitico delle Angiosperme.
Ovuli
Funicolo
Carpelli
Embrione ed endosperma
Embrione di monocotiledone
Logge carpellari
Sacco embrionale
Sezione di antera
Seme di monocotiledone (A) e
di dicotiledone
Frutto di dicotiledone (siliqua)
Nel fiore gli organi e le strutture riproduttive sono organizzati in un complesso estremamente
specializzato che garantisce la massima protezione ai gametofiti e la massima efficienza nella
selezione e nell’incontro dei gameti.
Nelle piante a fiore raggiunge dunque la massima espressione quella tendenza evolutiva verso il
perfezionamento dei meccanismi riproduttivi che è alla base della possibilità di insorgenza di nuove
forme e nuove funzioni da sottoporre al vaglio della selezione. È proprio grazie a questo che tra le
angiosperme si è potuta evolvere quell’enorme varietà morfologica e fisiologica che ha loro
consentito di diffondersi e diventare il gruppo di vegetali attualmente dominante in quasi tutti gli
ambienti terrestri.