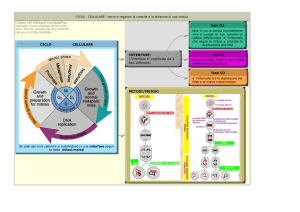
CICLO
CELLULARE
E’ IL MECCANISMO ESSENZIALE PER CUI TUTTI GLI ORGA=
NISMI SI RIPRODUCONO.
RIPRODUCONO.
NELLE SPECIE UNICELLULARI (BATTERI, LIEVITI…
LIEVITI…)) CIA=
SCUNA DIVISIONE CELLULARE GENERA UN NUOVO ORGA=
NISMO COMPLETO.
NELLE SPECIE MULTICELLULARI VICEVERSA LUNGHE E
COMPLESSE DIVISIONI CELLULARI SONO NECESSARIE
PER PRODURRE UN ORGANISMO FUNZIONANTE.
…NEGLI
EUCARIOTI PLURICELLULARI
TRE PROCESSI SONO NECESSARI PER FAR SI
CHE DA UNA CELLULA (ZIGOTE
(ZIGOTE)) SI SVILUPPI
UN ORGANISMO COMPLETO:
1)
LA SINGOLA CELLULA DEVE CRESCERE E DIVIDERSI
PER DARE ORIGINE A PIU’ CELLULE (CICLO
(CICLO CELLULARE);
CELLULARE);
2) LE CELLULE CHE DA ESSA ORIGINANO DEVONO
ACQUISIRE VIA VIA DELLE PROPRIETA’ PECULIARI,
TRASCRIVENDO SPECIFICI GENI CHE LE PORTINO A
FORMARE I VARI TESSUTI DELL’ORGANISMO
(DIFFERENZIAMENTO
DIFFERENZIAMENTO);
);
3)
MANTENIMENTO DELL’OMEOSTASI CELLULARE
ATTRAVERSO UN OPPORTUNO CONTROLLO
DELLA MORTE CELLULARE (APOPTOSI
(APOPTOSI))
QUANDO UNO DI QUESTI MECCANSIMI NON FUNZIONA
CORRETTAMENTE POSSONO INSORGERE MALATTIE NEOPLASTICHE
E DEGENERATIVE.
IL CICLO CELLULARE
Interfase (Fasi G1, S e G2)
(ca. 2020-24 h. – DI CUI
1010
-12 h. DI FASE S)
Mitosi (ca. 1 h.)
OPPURE
Meiosi
(da molte ore a decenni)
IL CICLO CELLULARE – USCITA DAL CICLO
NEGLI ORGANISMI PLURICELL.
(es.:UOMO) ALCUNE CELLULE
PROLUNGANO ENORMEMEMENTE
LA DURATA DELLA FASE G1,
POICHE’ NON SONO PRESENTI
I SEGNALI IDONEI A FAR
PARTIRE LA PROLIFERAZIONE
OPPURE PERCHE’ LA CELLULA SI E’
ESTREMAMENTE SPECIALIZZATA
(DIFFERENZIATA) E IN QUEL PARTICOLARE TESSUTO
NON E’ RICHIESTA CRESCITA ULTERIORE.
QUANDO LA CELLULA NON SI DIVIDE PIU’ PER UN LUNGO
PERIODO DI TEMPO, SI DICE CHE E’ “USCITA DAL CICLO”
CELLULARE E SI TROVA IN FASE G0 (Es.: NEURONI O CELL. MUSC
MUSC.)
.)
ALTRI TESSUTI, INCEVE,
DOPO UN LUNGO PERIODO DI
“QUIESCENZA” IN FASE G0,
POSSONO, SOTTO APPROPRIATI
STIMOLI RIENTRARE NEL CICLO
E RINNOVARSI (ES: FEGATO).
INFINE ALTRI TIPI DI TESSUTO
(ES: TESS. EPITELIALI) SONO
IN CONTINUA RIGENERAZIONE.
LE CELLULE CHE SI DIVIDONO
IN CONTINUAZIONE SONO LE
CELLULE STAMINALI.
STAMINALI.
CARATTERISTICA BASE DI QUESTE
CELLULE E’ CHE HANNO UN CICLO
CELLULARE “ASIMMETRICO”:
IN GENERE UNA CELLULA FIGLIA
CONTINUA A RIMANERE STAMINALE, MENTRE L’ALTRA INIZIA
UN PERCORSO DI DIFFERENZIAMENTO, CONTINUANDOSI A
DIVIDERE, MA CON UN NUMERO DI DIVISIONI LIMITATO NEL
TEMPO.
IN FASE G1 LE CELLULE SONO DIPLOIDI (2n) – CIOE’ AVRANNO UN
NUMERO DI COPPIE DI CHR
CHR.. TIPICO DELLA SPECIE CONSIDERATA.
OGNI CROMOSOMA (DI
(DI UNA COPPIA) SARA’ COSTITUITO DA UN
UNICO CROMATIDIO (CIOE’ DA UNA SOLA MOLECOLA DI DNA),
CARATTERIZZATO DA UN CENTROMERO E DUE BRACCIA [p (corto) E
q (lungo)]. DURANTE LA FASE S, LA CELLULA CONTINUA AD AVERE
UN ASSETTO DIPLOIDE (2n) MA OGNI CROMOSOMA SARA’ COSTI=
TUITO DA DUE CROMATIDI FRATELLI, UNITI IN PROSSIMITA’ DEL
CENTROMERO DA MOLECOLE PROTEICHE DETTE COESINE
COESINE..
NOTA: IN QUESTO CASO NELLA FIGURA
NOTA:
“2n” E “4n” STA AD INDICARE LA
QUANTITA’ DI DNA, O MEGLIO, IL
NUMERO DI FILAMENTI DI CUI
CIASCUN CROMOSOMA E’ COSTITUITO!
STUDIO DEL CICLO CELLULARE NEL LIEVITO
1) ORGANISMI CHE SI RIPRODUCONO
RAPIDAMENTE (COME I BATTERI)
2) PROLIFERANO IN UNO STATO
APLOIDE-- QUESTO E’ IMPORTANTE
APLOIDE
PER ISOLARE E STUDIARE MUTAZIONI
CHE INATTIVANO UN GENE
GRAZIE A LORO SONO STATI IDENTIFICATI
GENI cdc CHE REGOLANO IL CICLO
I MUTANTI cdc SONO DETTI CONDIZIONALI
IN RELAZIONE ALLA TEMPERATURA.
STUDIO DEL CICLO CELLULARE NELLO XENOPUS
LA FECONDAZIONE DELL’UOVO DI XENOPUS INNESCA UNA SERIE DI DIVISIONI
DETTE DI CLIVAGGIO – UNA SINGOLA CELLULA GIGANTE SI DIVIDE SENZA
CRESCERE ULTERIORMENTE PER GENERARE UN EMBRIONE CONTENENTE
MIGLIAIA DI CELLULE PIU’ PICCOLE. DOPO 11 DIVISIONI, NEL GIRO DI CA
CA.. 7H. SI
PRODURRA’ UN EMBRIONE FATTO DA 212 (4096) CELLULE. SI ALTERNANO SOLO FASI S ED M
DI CA
CA.. 15 MIN
MIN.. CIASCUNO, SENZA FASI G!
VANTAGGI DEGLI STUDI IN XENOPUS:
• CICLO SEMPLIFICATO
• GRANDI DIMENSIONI
E’ RELATIVAMENTE FACILE INIETTARE SOSTANZE
NELL’UOVO DELLO XENOPUS PER DETERMINARE
L’EFFETTO SULLA PROLIFERAZIONE – IN TAL MODO
SONO STATI INDIVIDUATI NUOVI GENI REGOLATORI DEL
CICLO.
IL BrdU E’ UN ANALOGO ARTIFICIALE DELLA TIMIDINA.
I NUCLEI CHE HANNO INCORPORATO BrdU SONO VISUALIZZATI
COLORANDOLI CON Ab ANTI
ANTI--BrdU
BrdU..
INDICE DI MARCATURA
MARCATURA:: DALLA PROPORZIONE DI CELLS
MARCATE NELLA POPOLAZIONE SI PUO’ STIMARE LA DURATA
DELLA FASE S (COME FRAZIONE DELLA INTERA DURATA
DEL CICLO CELLULARE)
INDICE MITOTICO:
MITOTICO: PROPORZIONE DI CELLULE IN MITOSI
DI NORMA IN UNA POPOLAZIONE DI CELLULE CHE STANNO
PROLIFERANDO RAPIDAMENTE MA IN MODO ASINCRONO
IN OGNI MOMENTO CIRCA IL 30/40 % SONO IN FASE S
IL CITOFLUORIMETRO E’ LO STRUMENTO DI
ELEZIONE PER LO STUDIO DEL CICLO CELLULARE
TRE CATEGORIE DI CELLULE
IN BASE AL LEGAME DELLO IODURO
DI PROPIDIO (PI):
1) CELLULE IN FASE G1 (CHE ANCORA
DEVONO REPLICARE IL DNA);
2) CELLULE IN FASE G2/M (CHE HANNO
REPLICATO IL DNA);
3) CELLULE IN FASE S (QUANTITA’ INTER=
MEDIA DI DNA.
CONTROLLO DEL CICLO CELLULARE
PROTEINE CHE REGOLANO IL CICLO
SCOPERTE ALLA FINE DEGLI ANNI
1980.
SISTEMA DI CONTROLLO BASATO SU:
1) OROLOGIO O TIMER;
TIMER;
2) MECCANISMO DI REGOLAZIONE
DELL’ORDINE DEGLI EVENTI;
3) MECCANISMO CHE ASSICURI
CHE CIASCUN EVENTO VENGA
SCATENATO UNA SOLA VOLTA
PER CICLO;
CICLO;
4) INTERRUTTORI BINARI (ON/OFF);
5) ROBUSTEZZA
ROBUSTEZZA;;
6) ADATTABILITA
ADATTABILITA’.
’.
I PUNTI DI CONTROLLO OPERANO
GENERALMENTE ATTRAVERSO
SEGNALI INTRACELLULARI
NEGATIVI.
LA TRANSIZIONE TRA UNA FASE E L’ALTRA DEL CICLO E’
SOTTOPOSTA A CONTINUE VERIFICHE,
VERIFICHE, PER ESSERE SICURI CHE
LE PROCEDURE CARATTERISTICHE DI UNA FASE SIANO STATE
COMPLETATE.
3 PRINCIPALI “CHECK“CHECK-POINT”:
1) CHECKPOINT DI INGRESSO IN FASE S (TRANSIZIONE
(TRANSIZIONE G1-S);
2) CHECKPOINT DI INGRESSO IN FASE M (TRANSIZIONE
(TRANSIZIONE G2-M);
3) CHECKPOINT DI COMPLETAMENTO FASE M (METAFASE
(METAFASE –
CITODIERESI))
CITODIERESI
REGOLAZIONE DEL CICLO CELLULARE
IL MECCANISMO FONDAMENTALE CHE REGOLA IL CICLO CONSISTE
IN UNA MODIFICAZIONE POSTPOST-TRADUZIONALE QUALE LA
FOSFORILAZIONE DEI RESIDUI SERINA E TREONINA DI
SPECIFICHE PROTEINE CELLULARI. ATTORI CHIAVE IN QUESTO
PROCESSO SONO UNA CHINASI (CDK
(CDK)) ED IL SUO STRETTO
INTERATTORE (CICLINA
(CICLINA).
). A LORO VOLTA LA LORO ATTIVITA’ E’
REGOLATA!
I CAMBIAMENTI CICLICI NEI LIVELLI DELLE CICLINE PORTANO ALL’ASSEMBLAGGIO
ED ALL’ATTIVAZIONE CICLICI DEI COMPLESSI CICLINACICLINA-CDK. QUESTA ATTIVAZIONE
SCATENA A SUA VOLTA EVENTI DEL CICLO CELLULARE.
IMPEGNO DELLA
INNESCO DELLA INNESCO DEL MACCHINARIO
CELLULA A DUPLICARE DUPLICARE DEL DNA
DELLA MITOSI
IL DNA
QUATTRO CLASSI DI CICLINE E CORRISPETTIVE CDK,
CIASCUNA DEFINITA DALLO STADIO DEL CICLO CELLULARE
NELLA MAGGIORPARTE DELLE CELLULE UNA QUARTA CLASSE DI CICLINE (CICLINE
(CICLINE G1) AIUTA
A PROMUOVERE IL PASSAGGIO ATTRAVERSO START (O PUNTO DI RESTRIZIONE).
NELLE CELLULE DI LIEVITO ESISTE UN’UNICA CDK CHE, A SECONDA DELLO STADIO DEL CICLO,
SI LEGA A VARIE CICLINE.
NELLE CELLULE DEI VERTEBRATI CI SONO 4 CDK:
CDK: DUE INTERAGISCONO CON LE CICLINE G1;
UNA CON LE CICLINE G1/S ED UNA CON LE CICLINE M.
IN ASSENZA DI CICLINA IL SITO ATTIVO DI CDK E’ PARZIALMENTE
OSCURATO!!
OSCURATO
AL MOMENTO DEL LEGAME DELLA CICLINA CDK SI ATTIVA SOLO
PARZIALMENTE..
PARZIALMENTE
NEL MOMENTO IN CUI LA KINASI ATTIVANTE CDK (CAK
(CAK)) FOSFORILA
UN AA. VICINO AL SITO ATTIVO DELLA CDK, SI HA L’ATTIVAZIONE
COMPLETA DEL COMPLESSO CICLINA/CDK!
LA CONCENTRAZIONE DI CICLINE STA ALLA BASE DELL’ATTIVAZIONE DELLE CDK.
QUESTE ULTIME POSSONO ANCHE ESSERE ATTIVATE O INIBITE DA SPECIFICI CICLI
DI DEFOSFORILAZIONE/FOSFORILAZIONE!
KINASI Wee1 INIBISCE L’ATTIVITA’ Cdk
FOSFATASI Cdc25 AUMENTA ATTIVITA’ Cdk
I COMPLESSI CICLINACICLINA-CDK SONO INATTIVATI DA PROTEOLISI REGOLATA DI CICLINE
A CERTI STADI DEL CICLO.
IL MECCANISMO E’ SPESSO DIPENDENTE DAL
SISTEMA UBIQUITINA/PROTEASOMA!
UBIQUITINE – PROTEINE PICCOLE
(76 AA.) UBIQUITARIE
E1 ATTIVA LE UBIQUITINE
E2 IN ASSOCIAZIONE CON E3
PERMETTONO LA LIGAZIONE
DELLE UBIQUITINE ALLE PRO=
TEINE TARGET.
LA POLIUBIQUITINAZIONE
CONSISTE NEL LEGAME MULTIPLO
DI PIU’ UBIQUITINE. IN PARTICO=
LARE IL CC-TERMINALE DI CIA=
SCUNA UBIQUITINA LEGA UNA
LISINA DELLA UBIQUITINA CHE
LA PRECEDE.
SPESSO IN BASE AL RESIDUO DI Lys UBIQUITINATO LE PROTEINE POSSONO
SUBIRE UN DESTINO (DEGRADAZIONE) OD UN ALTRO.
NEL CICLO CELLULARE DELLE CELLULE EMBRIONALI DI XENOPUS (V.
PRIMA) IL CONTROLLO DEL CICLO CELLULARE DIPENDE QUASI ESCLUSI=
VAMENTE DA MECCANISMI POSTPOST-TRASCRIZIONALI (FOSFORILAZIONE/
(FOSFORILAZIONE/DE=
DE=
FOSFORILAZIONE…).
FOSFORILAZIONE…
).
NEI CICLI CELLULARI DELLA MAGGIORPARTE DEI TIPI CELLULARI INVECE
IL CONTROLLO TRASCRIZIONALE FORNISCE UN LIVELLO AGGIUNTO DI
REGOLAZIONE.
CONTROLLO DELLA FASE S
LA CELLULA DEVE REPLICARE SOLO 1 VOLTA IL PROPRIO GENOMA AD OGNI CICLO
CELLULARE!
SOLTANTO CELLULE IN G1 SONO COMPETENTI AD INIZIARE LA REPLICAZIONE DEL
DNA, MENTRE LE CELLULE IN G2 (CHE HANNO GIA’ COMPLETATO LA FASE S) NON
SONO IN GRADO DI REPLICARE IL PROPRIO DNA!
1) LA PROTEINA REGOLATRICE Cdc6 LEGA
IL COMPLESSO DI RICONOSCIMENTO DELLA
ORIGINE (ORC) A LIVELLO DELLE DIVERSE
ORIGINI DI REPLICAZIONE (INIZIO G1);
2) ATTACCO DELLE PROTEINE Mcm E FORMAZI=
NE DEL COMPLESSO PREREPLICATIVO(pre
PREREPLICATIVO(pre--RC
RC));
3) S-Cdk IN ASSOCIAZIONE AD UNA KINASI SCATE
NA A QUESTO PUNTO L’INIZIO DI REPLICAZIO=
NE DEL DNA;
4) S-Cdk QUINDI FA DISSOCIARE Cdc6 DA ORC DOPO
CHE UN ORIGINE HA FUNZIONATO – CONSEGUENTE
DISASSEMBLAGGIO DI pre
pre--RC
RC!!
5) S-Cdk INOLTRE FOSFORILA Cdc6 ED Mcm DETER=
MINANDO LA UBIQUITINAZIONE DI Cdc6 E LA
ESPORTAZIONE DAL NUCLEO AL CITOSOL DI
Mcm;;
Mcm
6) ANCHE M-Cdk E G1/S
G1/S-CdkAIUTANO AD EVITARE
LA RIRI-REPLICAZIONE DEL DNA!
7) ALLA FINE DELLA MITOSI TUTTA L’ATTIVITA’
CDK SI RIDUCE A ZERO: CONSEGUENZA E’ LA
DEFOSFORILAZIONE DI Cdc6 E Mcm E RICOSTITU=
ZIONE DEL pre
pre--RC
RC..
CONTROLLO DELLA FASE M
L’ATTIVAZIONE DELLA M-Cdk INIZIA CON L’ACCUMULO DELLA CICLINA M
L’ATTIVAZIONE
(LIEVITO) O CICLINA B (VERTEBRATI)
A MONTE LA KINASI
POLO FOSFORILA
Cdc25 ATTIVANDOLA
NELLE CELLULE EMBRIONALI LA SINTESI DI CICLINA M (O B) E’ COSTANTE
PER TUTTO IL CICLO CELLULARE E L’ACCUMULO DI CICLINA M RISULTA
DA UNA DIMINUZIONE DELLA SUA DEGRADAZIONE.
NELLA MAGGIORPARTE DEI TIPI CELLULARI LA CICLINA M AUMENTA DU=
RANTE LE FASI G2 ED M A CAUSA DI UN AUMENTO DELLA TRASCRIZIONE
DEL GENE DELLA CICLINA M!
MECCANISMI MOLECOLARI IGNOTI ASSICURANO CHE LA MITOSI (M
(M-Cdk
Cdk))
SI ATTIVI SOLO DOPO CHE L’INTERO DNA SI SIA DUPLICATO.
CELLULE DI MAMMIFERO TRATTATE CON IDROSSIUREA (INIBITORE CHIMICO
DELLA SINTESI DEL DNA) NON PROGREDISCONO IN MITOSI
CELLULE DI MAMMIFERO TRATTATE CONTEMPORANEAMENTE CON IDROS=
SIUREA E CAFFEINA MOSTRANO DIFETTI NEL PUNTO DI CONTROLLO DELLA
REPLICAZIONE DEL DNA E PROGREDISCONO VERSO UNA MITOSI SUICIDA!
M-Cdk ATTIVATA DETERMINA:
• FOSFORILAZIONE E DEPOLIMERIZZAZIONE DELLE LAMINE NUCLEARI;
• CONDENSAZIONE DEI CROMOSOMI;
• RIARRANGIAMENTI DEI MICROTUBULI ED ASSEMBLAGGIO DEL FUSO
MITOTICO.
IL COMPLESSO CHE PROMUOVE L’ANAFASE (APC) FA “SCATTARE” L’INTERRUTTORE
CHE INIZIA LA SEPARAZIONE DEI CROMATIDI FRATELLI.
APC E’ UN’UBIQUITINO LIGASI CHE
UNA VOLTA ATTIVATA MEDIANTE LE=
GAME CON LA SUBUNITA’ ATTIVATRICE
Cdc20, UBIQUITINA LA CICLINA M!
IL BERSAGLIO PIU’ IMPORTANTE DI APC E’ LA PROTEINA SECURINA
SECURINA..
ALLA FINE DELLA METAFASE APC DISTRUGGE LA SECURINA LIBERANDO LA PROTEASI
SEPARASI CHE TAGLIA LE SUBUNITA’ DELLE COESINE PERMETTENDO LA SEPAZIONE
DEI CROMATIDI FRATELLI!
PUNTO DI CONTROLLO DELL’ATTACCO DEL FUSO: PROTEINE Mad2
VENGONO MASSIVAMENTE RECLUTATE SU CINETOCORI NON ATTACCATI
ALLE FIBRE DEL FUSO MITOTICO. CIO’ INIBISCE Cdc20Cdc20-APC E LA DISTRU=
ZIONE DELLA SECURINA. COSI’ LA SEPARAZIONE DEI CROMATIDI NON PUO’
AVVENIRE FINCHE’ L’ULTIMO CINETOCORE NON SI E’ ATTACCATO!
L’INATTIVAZIONE STABILE DI CDK A
SEGUITO DELLA DEGRADAZIONE DELLA
CICLINA M PORTA ALLA TRANSIZIONE
DALLA FASE M ALLA FASE G1.
QUESTA INATTIVAZIONE DI CDK E’
FRUTTO DELL’ATTIVAZIONE DI Hct1,
ACCUMULO DELLA PROTEINA CKI
(INIBITORE DELLA CDK) ED INIBIZIONE
DELLA TRASCRIZIONE DEL GENE DELLA
CICLINA.
pRb:: IL GUARDIANO DEL CICLO
pRb
L’EVIDENZIAZIONE DEI SISTEMI DI CONTROLLO DELLA
TRANSIZIONE G1 – S NEI MAMMIFERI, E’ STATA ILLUMINANTE
PER IDENTIFICARE LE ALTERAZIONI
CHE PROMUOVONO LA PROLIFERAZIONE INCONTROLLATA
DELLE CELLULE TUMORALI.
UN CONTROLLORE ESSENZIALE PER LA PROGRESSIONE DEL
CICLO DALLA FASE G1 ALLA S E’ COSTITUITO DAL COMPLESSO
E2F/DP1, CUI PUO’ ASSOCIARSI L’INIBITORE DELLA
PROGRESSIONE DEL CICLO pRb
pRb!!
SCHEMA DELLA PROGRESSIONE DEL CICLO NEI
MAMMIFERI
pRb (Non fosforilato)
E2F/DP1
c-myc
myc,, c-fos
fos,, c-jun (e vari altri protoprotooncogeni)
Cicline D
FOSFORILAZIONE DI pRb
p53: IL “GUARDIANO MOLECOLARE”
DELL’INTEGRITA’ DEL DNA
COME pRb
pRb,, p53 E’ UN ONCOSOPPRESSORE.
ONCOSOPPRESSORE. SOTTO LO STIMOLO DI DANNO AL
DNA, p53 TENDE A BLOCCARE LA PROGRESSIONE
ATTRAVERSO IL CICLO CELLULARE, FACENDO SI CHE GLI
EVENTUALI DANNI POSSANO ESSERE RIPARATI!
RIPARATI!
SE IL NUMERO DIMUTAZIONI ACCUMULATE E’ ECCESSIVO,
ALLORA LA CELLULA SCEGLIE LA VIA DELL’APOPTOSI
DELL’APOPTOSI!!
UNA RARA MALATTIA GENETICA E’ LA ATASSIA
TELANGECTASIA,, CAUSATA DA UN DIFETTO NELLE
TELANGECTASIA
PROTEINE CHINASI (ATM) CHE FOSFORILANO p53 IN RI=
SPOSTA AD UN DANNO AL DNA DA RAGGI X.
I PAZIENTI AFFETTI DA QUESTA PATOLOGIA SONO
MOLTO SENSIBILI AI RAGGI X E SOFFRONO DI UNA
AUMENTATA FREQUENZA DI CANCRO.
RIASSUMENDO, LA CELLULA, IN CASO DI EMERGENZA
BLOCCHERA’ LA PROGRESSIONE DEL CICLO, ENTRANDO
IN FASE G0 PER UNO DEI SEGUENTI MOTIVI:
1) DANNO AL DNA;
DNA;
2) PRESENZA NELL’AMBIENTE EXTRACELL. DI FATTORI
(ES: TGF b ) INIBITORI DI CRESCITA CELLULARE;
CELLULARE;
3) INIBIZIONE DA CONTATTO (IN CASO DI ECCESSIVO
INCREMENTO DELLA DENSITA’ CELLULARE)
SCHEMA RIASSUNTIVO DEL SISTEMA DI CONTROLLO DEL CICLO CELLULARE
MITOSI
E’ IL PROCESSO DI DIVISIONE CELLULARE CHE SI VERIFICA
NELLE CELLULE SOMATICHE ED IN QUELLE INDIFFERENZIATE
DELLA LINEA GERMINALE, ASSICURANDO ALLE CELLULE FIGLIE
UN’ASSEGNAZIONE OMOGENEA DELLE COMPONENTI CELLULARI,
INCLUSO IL DNA ORGANIZZATO PER L’OCCASIONE IN CROMOSOMI!
SI IDENTIFICANO LE SEGUENTI FASI:
FASI:
i) PROFASE;
ii) PROMETAFASE;
iii) METAFASE;
iv) ANAFASE;
v) TELOFASE;
vi) CITODIERESI O CITOCHINESI.
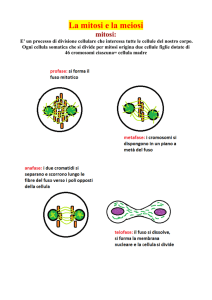
PROFASE
LA PROFASE HA INIZIO CON IL COMPATTAMENTO DEI CROMOSOMI,
GRAZIE A DEGLI ENZIMI (CONDENSINE
(CONDENSINE)) ATTIVATE DA CDK.
INIZIA AD ORGANIZZARSI ANCHE IL FUSO MITOTICO!
IN PARTICOLARE I CENTROSOMI (DA CUI SI DIPARTONO LE FIBRE DEL
FUSO) SI DISPONGONO AI POLI OPPOSTI DELLA CELLULA.
PROMETAFASE
LA PROMETAFASE SI FA COINCIDERE CON LA FRAMMENTAZIONE IN
PICCOLE VESCICOLE DELL’INVOLUCRO NUCLEARE,
NUCLEARE, CONSEGUENTE ALLA
DEPOLIMERIZZAZIONE DELLE LAMINE NUCLEARI (SEMPRE PER FOSFO=
RILAZIONE DA PARTE DI CDK).
SI INCOMINCIA A DELINEARE SEMPRE MEGLIO IL FUSO MITOTICO.
QUESTO E’ IN PARTICOLARE COSTITUITO DA:
i) FIBRE DEL CINETOCORE;
CINETOCORE;
ii) FIBRE ASTRALI;
ASTRALI;
iii) FIBRE INTERPOLARI.
INTERPOLARI.
METAFASE
I CROMOSOMI SI DISPONGONO IN POSIZIONE MEDIANA RISPETTO
I DUE POLI DEL FUSO, IN UNA STRUTTURA NOTA COME PIASTRA
METAFASICA.. I MICROTUBULI DEL FUSO, ATTACCATI AI DUE
METAFASICA
CINETOCORI DI OGNI CROMATIDIO FRATELLO, TENDONO A
STRATTONARE QUESTI ULTIMI IN DIREZIONI OPPOSTE (AI POLI
OPPOSTI DELLA CELLULA). QUANDO SI RAGGIUNGE UN EQUILIBRIO
TRA QUESTE FORZE AGENTI IN DIREZIONI OPPOSTE, I CROM.
SARANNO DISPOSTI IN PIASTRA METAFASICA!
ANAFASE
DOPO CHE, ATTRAVERSO UN ENNESIMO CHECKPOINT
CHECKPOINT,, LA CELLULA
SI E’ “ASSINCERATA” CHE LA DISPOSIZIONE DEI CROMOSOMI A LIVELLO
DELLA PIASTRA METAFASICA E’ CORRETTA, VIENE ATTIVATO IL
COMPLESSO MULTIPROTEICO APC (A
(ANAPHASE PROMOTING COMPLEX)
MEDIANTE FOSFORILAZIONE! TRAMITE QUESTO COMPLESSO LE COESINE
CHE TENGONO INSIEME I CROMATIDI FRATELLI DI CIASCUN CROMOSOMA
VENGONO DEGRADATEE CIASCUNO DI ESSI MIGRA AI POLI OPPOSTI
DELLA CELLULA. NEL CONTEMPO LA CICLINA B VIENE DEGRADATA (SEMPRE
DA APC), IN MODO TALE CHE L’ATTIVITA’ CDK VIENE INIBITA!
TELOFASE
ALL’INIZIO DELLA TELOFASE OGNI CROMATIDIO FRATELLO E’ GIA’ STATO
TRASPORTATO ALLE DUE OPPOSTE REGIONI DELLA CELLULA.
CELLULA.
SI RIFORMA L’INVOLUCRO NUCLEARE MEDIANTE DEDE-FOSFORILAZIONE
DELLA LAMINA NUCLEARE ED I CROMOSOMI INIZIANO A DECOMPATTARSI.
DECOMPATTARSI.
ANCHE GLI ALTRI ORGANULI CELLULARI CHE SI SONO EQUAMENTE
DISTRIBUITI AI POLI OPPOSTI DELLA CELLULA INIZIANO A
RIORGANIZZARSI!
CITOCHINESI
NELLA CITOCHINESI L’AZIONE CONTINUA DELL’ANELLO
DELL’ANELLO CONTRATTILE
PROVOCA LA SEPARAZIONE IN DUE DEL CITOPLASMA CHE FISSA LA
FORMAZIONE DI DUE NUOVE CELLULE.
MEIOSI
NEGLI ORGANISMI EUCARIOTICI CHE SI RIPRODUCONO
SESSUALMENTE, I GAMETI DEVONO CONTENERE UNA SOLA
COPIA DEL CORREDO CROMOSOMICO (DA 2n AD n)!
LA MEIOSI INTERESSA PROPRIO LE CELLULE DELLA LINEA
GERMINALE ED ENTRA IN GIOCO DURANTE LA GAMETOGENESI
GAMETOGENESI!!
L’INTERO PROCESSO PUO’ ESSERE DIVISO IN DUE
FASI:
i)
PRIMA DIVISIONE MEIOTICA (MEIOSI
(MEIOSI I):
I):
E’ UNA DIVISIONE RIDUZIONALE;
RIDUZIONALE;
ii) SECONDA DIVISIONE MEIOTICA (MEIOSI
(MEIOSI II)
II)
E’ UNA DIVISIONE EQUAZIONALE.
EQUAZIONALE.
LA MEIOSI HA TEMPI MOLTO PIU’ LUNGHI DELLA
MITOSI (NELLA NOSTRA SPECIE DURA 24 GIORNI
NEL MASCHIO E DECENNI NELLA FEMMINA)
n - indica la ploidia
c - indica il contenuto di DNA
II Meiosi
I Meiosi
2n 2c
1n 2c
1n 1c
1 cellula
2 cellule
4 cellule
MEIOSI I - PROFASE
LA PROFASE DELLA MEIOSI I E’ MOLTO LUNGA
E SI POSSONO DISTINGUERE LE SEGUENTI
SOTTOFASI:
i)
LEPTOTENE;;
LEPTOTENE
ii) ZIGOTENE
ZIGOTENE;;
iii) PACHITENE
PACHITENE;;
iv) DIPLOTENE
DIPLOTENE;;
v) DIACINESI
DIACINESI..
PROFASE – LEPTOTENE E ZIGOTENE
LA FASE INIZIALE DELLA PROFASE I MEIOTICA RICORDA LA
PROFASE MITOTICA: I CROMOSOMI SI INIZIANO A COMPATTARE,
E L’INVOLUCRO NUCLEARE SI FRAMMENTA.
UNA DIFFERENZA PERO’ E’ MOLTO IMPORTANTE: I CROMOSOMI
OMOLOGHI (DICROMATIDICI) SI APPAIANO A FORMARE
STRUTTURE NOTE COME TETRADI
TETRADI!!
PROFASE – PACHITENE
DURANTE QUESTA FASE VIENE A FORMARSI UN COMPLESSO
MULTIPROTEICO NOTO COME COMPLESSO SINAPTONEMALE.
SINAPTONEMALE.
TALE COMPLESSO FAVORISCE IL CROSSING OVER,
OVER, SCAMBIO
DI MATERIALE GENETICO FRA I CROMATIDI NON FRATELLI
DI CROMOSOMI OMOLOGHI!
OMOLOGHI!
PROFASE – DIPLOTENE
IL COMPLESSO SINAPTONEMALE SCOMPARE ED I CROMOSOMI
OMOLOGHI, PUR TENDENDO A SEPARARSI, RIMANGONO
UNITI IN PROSSIMITA’ DELLE ZONE DOVE E’ AVVENUTO IL
CROSSING OVER (CHIASMI
(CHIASMI)!
)! SU DI UNA TETRADE POSSONO
VERIFICARSI PIU’ CHIASMI (NELL’UOMO SE NE OSSERVANO
IN MEDIA 2 O 3 PER TETRADE).
PROFASE – DIACINESI
CON LA DIACINESI SI CONCLUDE LA PROFASE E SI ATTUA IL
PASSAGGIO ATTRAVERSO LA METAFASE I.
DURANTE LA DIACINESI I CROMOSOMI SI CONDENSANO
ULTERIORMENTE, I NUCLEOLI SCOMPAIONOE SI CONCLUDE
L’ORGANIZZAZIONE DEL FUSO.
IMPORTANTI DIFFERENZE CON
LA MITOSI SONO:
i) ALLINEAMENTO DI COPPIE DI CROMOSOMI
OMOLOGHI IN PIASTRA METAFASICA;
METAFASICA;
ii)) SOLO UNO DEI DUE CENTROMERI E’ LEGATO
ii
ALLE FIBRE DEL FUSO;
FUSO;
iii)) LE COESINE VENGONO DEGRADATE
iii
SOLO IN MEIOSI II.
Preparazione del
complesso di assemblaggio
02_11.jpg
I cromosomi omologhi iniziano
ad appaiarsi si formano i
bivalenti o tetradi
Avviene il crossing
crossing-over
Gli omologhi iniziano a separarsi
ma restano uniti nei punti in cui è
Avvenuto il crossing
crossing-over
Si visualizzano i chiasmi
I bivalenti diventano più contratti
MEIOSI II
CON LA SECONDA DIVISIONE MEIOTICA, A PARTIRE
DALLE DUE CELLULE APLOIDI CON CROMOSOMI
DICROMATIDICI, SI FORMANO QUATTRO CELLULE
APLOIDI CIASCUNA CON UN SET COMPLETO DI
CROMOSOMI MONOCROMATIDICI!
IN QUESTA FASE DELLA MEIOSI VENGONO RIPERCORSE
LE STESSE IDENTICHE TAPPE DELLA MITOSI.
NOTA: DURANTE L’OOGENESI LA DIVISIONE MEIOTICA
VIENE MOMENTANEAMENTE BLOCCATA IN METAFASE II
PER POI RIPRENDERE E COMPLETARSI DOPO CHE LA CEL=
LULA UOVO E’ FECONDATA!
IL SIGNIFICATO BIOLOGICO DELLA
MEIOSI: LA VARIABILITA’ GENETICA
LA DIVERSITÀ DEI GAMETI PRODOTTI DA
UN INDIVIDUO E’ PRINCIPALMENTE
DOVUTA A:
1) CROSSING
CROSSING-OVER
2) ASSORTIMENTO INDIPENDENTE
L’UNIONE DEI CORREDI GENOMICI DI DUE
GAMETI GENERA DUNQUE UN INDIVIDUO
(ZIGOTE) UNICO ED IRRIPETIBILE.
ASSORTIMENTO INDIPENDENTE IN ANAFASE I
NELL’UOMO IL SOLO ASSORTIMENTO
INDIPENDENTE PUO’ DETERMINARE LA
FORMAZIONE DI
2^23 (DUE POSSIBILI ORIENTAMENTI AI DUE
POLI DEL FUSO, IL TUTTO ELEVATO AL NUMERO
APLOIDE DI CROMOSIMI) (8.400.000) DIVERSE
COMBINAZIONI DI DISTRIBUZIONE DEI
CROMOSOMI MATERNI E PATERNI NEI GAMETI
…A QUESTO SI AGGIUNGA LA VARIABILITA’
DETERMINATA DAL CO
CO.. E’ PRATICAMENTE
IMPOSSIBILE CHE, A MENO DEI GEMELLI
MONOZIGOTI, VENGANO A COSTITUIRSI DUE
INDIVIDUI IDENTICI PER CASO.
CASO.