IL TESSUTO EPITELIALE
Generalità
Il tessuto epiteliale è un tessuto non vascolarizzato, formato da cellule a stretto contatto, con scarsa
sostanza intercellulare interposta. Esso poggia sempre su uno strato più o meno abbondante di
tessuto connettivo dai cui capillari riceve, per diffusione, le sostanze nutritive e l’ossigeno di cui
necessita. In tutte le sedi, tra epitelio e strato connettivo sottostante si interpone una sottile
membrana basale.
Il tessuto epiteliale,
ampiamente rappresentato nel corpo umano, possiede la caratteristica di
svolgere funzioni diverse in base alla propria localizzazione anatomica. Tra queste ricordiamo:
•
protezione contro danni meccanici, insulti chimici o fisici ed invasione di microorganismi
(superficie esterna del corpo);
•
prevenzione dalla disidratazione (superficie esterna del corpo);
•
secrezione (ghiandole)
•
escrezione di cataboliti (tubuli renali);
•
assorbimento di nutrienti (intestino);
•
recezione di sensibilità mediante recettori tattili, termici e dolorifici in esso contenuti (pelle,
cavità nasali, lingua, orecchio).
In base alle funzioni svolte possiamo distinguere diversi tipi di epiteli:
•
Epiteli di rivestimento: rivestono la superficie esterna del corpo, le cavità interne del corpo
che comunicano con l’esterno (canale digerente, apparato respiratorio, canale urogenitale, dotti
escretori delle ghiandole esocrine), le cavità interne del corpo che non comunicano con l’esterno
(cavità pericardica, cavità pleuriche e cavità peritoneale), la superficie interna dei vasi sanguigni e
linfatici.
Gli epiteli di rivestimento assumono nomi specifici in base alla localizzazione: epidermide è il
nome dell’epitelio che riveste la superficie esterna del corpo; endotelio è il nome che viene
utilizzato per definire l’epitelio che riveste la superfice interna dei vasi sanguigni e linfatici; il
termine mesotelio viene invece utilizzato per definire l’epitelio che riveste le cavità pericardica,
pleurica e peritoneale e la superficie degli organi in esse contenuti (es. stomaco, intestino, cuore,
polmoni). Il mesotelio che riveste gli organi interni è definito mesotelio viscerale, mentre quello che
riveste le superfici interne del corpo è definito mesotelio parietale.
In tutte queste sedi anatomiche, l’epitelio poggia su uno strato di connettivo. A sua volta, l’insieme
dell’epitelio e dello strato connettivale sottostante viene definito con specifici nomi: la cute o pelle è
il nome che si utilizza per indicare l’insieme dell’epidermide con il tessuto connettivo sottostante; il
termine mucosa si utilizza per indicare l’insieme dell’epitelio che riveste le cavità del corpo che
comunicano con l’esterno con il tessuto
connettivo sottostante, come ad esempio la mucosa
gastrica ed intestinale, sierosa è il nome che viene dato al mesotelio ed al sottile strato di tessuto
connettivo su cui esso poggia.
Epiteli ghiandolari: posseggono funzione secernente. Formano la componente secernente
(parenchima) delle ghiandole esocrine ed endocrine.
•
Epiteli sensoriali: sono epiteli specializzati nel rilevare sensazioni e trasmettere le
informazioni al sistema nervoso centrale. Negli epiteli sensoriali le cellule che hanno la funzione di
ricevere e trasmettere le sensazioni al sistema nervoso sono disperse tra le cellule epiteliali. Queste
cellule sono provviste di recettori specifici, mediante i quali possono ricevere gli stimoli, ma non
debbono essere confuse con le cellule nervose. Le cellule epiteliali sensoriali, infatti, non
posseggono l’assone e sono invece circondate dalle espansioni terminali di fibre nervose
appartenenti a neuroni sensoriali il cui soma è localizzato nei gangli cerebro-spinali. Appartengono
a questa categoria le cellule gustative della lingua, le cellule acustiche e i neuromasti dell’orecchio
interno. Fanno eccezione le cellule olfattive della mucosa olfattiva e le cellule dei coni e dei
bastoncelli della retina, che sono cellule nervose vere e proprie.
In questo capitolo ci soffermeremo sugli epiteli di rivestimento.
INDICE
Generalità
12.1 Caratteristiche citologiche delle cellule epiteliali
12.1.1 Citoscheletro
12.1.2 Specializzazioni della superficie laterale
12.1.3 Specializzazioni della superficie apicale
12.1.4 Specializzazioni della superficie basale
12.1.5 Polarità
12.2 Classificazione degli epiteli di rivestimento.
12.2.1 Epitelio pavimentoso semplice.
12.2.2 Epitelio cubico semplice.
12.2.3 Epitelio cilindrico semplice.
12.2.4 Epitelio pavimentoso stratificato.
12.2.5 Epitelio cilindrico o cubico pluristratificato.
12.2.6 Epitelio pseudostratificato.
12.2.7 Epitelio di transizione.
Bibliografia
Caratteristiche citologiche delle cellule epiteliali
Le cellule epiteliali presentano caratteristiche morfologiche variabili da mettersi in relazione con le
molteplici funzioni dell’epitelio (protezione, assorbimento, secrezione, etc.)
Come già menzionato, le cellule epiteliali sono molto ravvicinate le une alle altre, con scarsa o nulla
sostanza intercellulare interposta. In virtù di questa loro organizzazione spaziale, nelle cellule
epiteliali si possono distinguere le seguenti zone o superfici:
•
una superficie apicale o libera: tale superficie è rivolta verso l’esterno, inteso come esterno
del corpo nel caso di epiteli che rivestono la superficie esterna del corpo, o interno del corpo
nel caso di epiteli che rivestono le superfici interne del corpo,
•
una superficie basale: tale superficie è rivolta verso il tessuto connettivo e poggia sulla
membrana basale, uno strato di materiale prodotto dalle cellule epiteliali stesse e ricco di
proteine e polisaccaridi;
•
una superficie laterale a contatto con le cellule adiacenti.
Le caratteristiche strutturali diverse di ciascuna superficie sono alla base delle diverse funzioni del
tessuto epiteliale.
Da un punto di vista citologico, le principali caratteristiche delle cellule epiteliali sono dovute alla
presenza di :
1. abbondante citoscheletro;
2. specializzazioni della superficie apicale;
3. specializzazioni della superficie basale;
4. specializzazioni della superficie laterale.
12.1.1 Citoscheletro
Tutte le cellule posseggono un citoscheletro. La sua funzione più evidente è quella di fornire un
supporto meccanico alla cellula e di mantenerne costante la forma. La solidità e la resistenza del
citoscheletro sono caratteristiche importanti in tutte le cellule, ma diventano cruciali in alcuni tipi
cellulari come le cellule epiteliali, che sono sottoposte a considerevoli sollecitazioni meccaniche.
In aggiunta ai microtubuli di tubulina ed ai microfilamenti di actina, il citoplasma delle cellule
epiteliali è particolarmente ricco di un tipo di filamenti definiti filamenti intermedi, di circa 8-12 nm
di spessore, un valore compreso tra quello di 25 nm dei microtubuli e quello di 7 nm dei
microfilamenti. I filamenti intermedi, contrariamente agli altri componenti del citoscheletro,
responsabili del movimento dinamico delle vescicole cellulari, svolgono un ruolo essenzialmente
strutturale, consolidando la forma tipica della cellula e fissando la posizione di certi organuli nel suo
interno.
I filamenti intermedi sono costituti da proteine fibrose appartenenti alla famiglia delle cheratine. Le
cheratine si trovano esclusivamente nelle cellule epiteliali e nei derivati dell’epidermide (peli,
unghie) e rappresentano una famiglia composta da molte differenti proteine filamentose espresse in
differenti combinazioni. Nelle cellule differenziate degli epiteli stratificati, le cheratine
costituiscono circa l’80% del contenuto proteico totale. La funzione principale delle cheratine è
quella di fornire un sostegno meccanico alle cellule epiteliali, anche se la rilevanza di tale funzione
varia con il tipo di epitelio. Nell’epitelio pluristratificato, esposto costantemente a sollecitazioni
meccaniche, questa funzione risulta, ad
esempio, preminente.
Non così
nell’epitelio
monostratificato che riveste gli organi interni, come le sierose, o in quello che riveste la superficie
interna degli alveoli polmonari, poichè tali tipi di epiteli non sono soggetti a sollecitazioni
meccaniche. Nell’epitelio che riveste la superficie interna dell’intestino, infine, le cheratine
svolgono un ruolo importante nel mantenere la polarità cellulare.
Nei filamenti intermedi, monomeri di cheratina si associano a formare dei dimeri e questi a loro
volta dei tetrameri o proto filamenti. Più proto filamenti si associano parallelamente fra loro in
maniera sfalsata, creando dei lunghi proto filamenti. In ultimo, i proto filamenti si allineano a
formare i lunghi filamenti intermedi di cheratina.
I filamenti di cheratina mostrano una distribuzione perinucleare; dal nucleo si dirigono verso la
periferia della cellula, collegandosi ai desmosomi (vedi dopo): tale collegamento contribuisce al
mantenimento dell’adesione cellulare e conferisce sostegno meccanico alle cellule epiteliali.
12.1.2 Specializzazioni della superficie apicale
La superficie libera delle cellule epiteliali esibisce modificazioni collegate a funzioni come
l’assorbimento ed il trasporto.
Tali modificazioni sono essenzialmente legate alla presenza di due tipi di strutture cellulari:
•
microvilli
•
ciglia
Microvilli. I microvilli sono ripiegamenti della membrana plasmatica visibili al ME (microscopio
elettronico). La presenza dei microvilli contribuisce all’aumento della superficie cellulare libera. Il
numero e la forma dei microvilli sono correlati alla capacità di assorbimento della cellula. Cellule la
cui principale funzione è rappresentata dal trasporto di fluidi e dall’ assorbimento di metaboliti
presentano, infatti, microvilli strettamente ravvicinati ed a forma di dito di guanto, mentre cellule in
cui il trasporto e l’assorbimento sono meno attivi presentano microvilli più piccoli e disposti in
maniera irregolare. L’epitelio intestinale, specializzato nell’assorbimento di nutrienti, presenta
microvilli disposti ordinatamente, che al MO (microscopio ottico) appaiono come uno strato sottile
ma ben definito al di sopra della superficie libera delle cellule, chiamato orletto a spazzola.
I microvilli possono essere particolarmente lunghi e sottili ed in questo caso vengono definiti stereo
ciglia. Stereociglia sono presenti nelle cellule epiteliali che rivestono l’epididimo e nelle cellule
sensitive dell’orecchio interno.
Ciglia. Le ciglia sono presenti sulla superficie libera delle cellule che formano epiteli specializzati
nel trasporto di liquidi o muco, come nella trachea, bronchi ed ovidutti. La struttura delle ciglia è
tipicamente costituita da microtubuli che formano l’assonema e si inseriscono in un corpo basale
presente nel citoplasma. Al MO, le ciglia appaiono come strutture sottili che si estendono sulla
superficie libera delle cellule epiteliali. Le ciglia si muovono in maniera regolare e sincrona. Il
movimento delle ciglia è prodotto dal piegamento dell’assonema. Ogni ciglio compie singolarmente
un movimento simile a un colpo di frusta; nel loro insieme le ciglia si agitano in sincronia, creando
un movimento ondulatorio sulla superficie cellulare. Il movimento sincrono è fondamentale, poiché
permette al liquido o al muco che ricopre l’epitelio di muoversi nella direzione voluta.
12.1.3 Specializzazioni della superficie basale
Come già descritto, le cellule che formano il tessuto epiteliale posseggono una superficie apicale,
una superficie laterale a contatto con le cellule adiacenti ed una superficie basale che poggia sulla
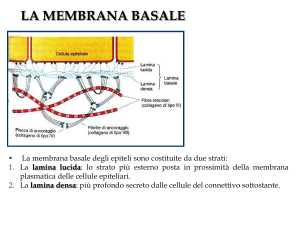
membrana basale. Il termine membrana non deve indurre in errore. In realtà si tratta di un sottile
strato privo di cellule ma ricco di proteine e polisaccaridi, secreto dalle cellule epiteliali.
La membrana basale si colora di rosso con l’eosina e per questo è poco evidente alla colorazione
H/E. Essa invece è ben visibile al MO in presenza di particolari colorazioni (colorazione PAS e
coloranti tipici per i glucosaminoglucani o GAG.
La struttura della membrana basale è complessa e l’utilizzo del ME è stato di grande aiuto nel
chiarirla. Al ME la membrana basale appare come uno strato di 50-100 nm di spessore. Questo
strato viene definito lamina basale. La lamina basale è a sua volta formata da due strati: la lamina
lucida, uno strato chiaro trasparente agli elettroni, posto appena al di sotto delle cellule epiteliali e la
lamina densa, uno strato scuro elettron-denso al di sotto della lamina lucida.
La lamina basale è costituita da fibre collagene, proteoglicani e glicoproteine (laminina, entactina,
fibronectina). I sottili filamenti contenuti nella lamina lucida ancorano la lamina densa alle cellule
epiteliali, mentre alcune fibre collagene ancorano la lamina basale ad uno strato definito lamina
reticolare, che risulta formata da fibre collagene prodotte dai fibroblasti del connettivo.
La membrana basale possiede ruoli importanti ed in parte non ancora completamente chiariti, ma
sicuramente rappresenta il supporto su cui poggiano le cellule epiteliali, fornisce protezione al
sottostante tessuto connettivo e regola lo scambio di macromolecole tra il connettivo e l’epitelio. La
membrana basale influenza anche il differenziamento e la proliferazione delle cellule epiteliali che
poggiano su di essa, fornendo inoltre una sorta di impalcatura che, come dei binari, guida la
migrazione delle cellule epiteliali durante lo sviluppo embrionale. (FIG. 8)
12.1.4 Specializzazioni della superficie laterale (Le giunzioni cellulari)
Come si è detto, le cellule epiteliali sono aderenti le une alle altre, ma non solo: infatti tra le
membrane delle regioni cellulari a stretto contatto, si stabiliscono delle strutture complesse
chiamate giunzioni cellulari, alla cui costruzione partecipano il citoscheletro e molti diversi tipi di
proteine.
Esistono numerosi tipi di giunzioni cellulari tra cellule epiteliali adiacenti. La diversa struttura delle
giunzioni cellulari consente di realizzare gradi diversi di adesione tra le cellule. In presenza di
giunzioni strette (occludenti), si verifica la fusione delle membrane cellulari di due cellule
adiacenti, che diventano così impermeabili persino al passaggio di molecole d’acqua. Esistono poi
giunzioni intermedie (aderenti e ancoranti), in cui proteine e materiale extracellulare collaborano a
far aderire due cellule vicine. In altri casi ancora, le giunzioni sono così intime (comunicanti) da
consentire il passaggio di materiale citoplasmatico da una cellula all’altra.
Le giunzioni cellulari svolgono diverse funzioni complessivamente importanti per il corretto
funzionamento dell’epitelio. Le giunzioni strette rafforzano l’unione tra le cellule, legandole
letteralmente attraverso “cuciture proteiche” che creano una barriera al passaggio di sostanze tra le
cellule. Le giunzioni aderenti e ancoranti forniscono stabilità meccanica a gruppi di cellule
epiteliali, che possono così funzionare come un’unità strutturale. Tali giunzioni consentono non
solo l’adesione tra cellule contigue, ma connettono anche l’epitelio alla sottostante membrana
basale. Le giunzioni comunicanti mettono in contatto il citoplasma di due cellule adiacenti,
consentendo la diffusione selettiva di molecole tra due cellule a contatto e facilitando così la
comunicazione cellulare. Le giunzioni cellulari contribuiscono, dunque, a fare dell’epitelio una
barriera protettiva efficace sia meccanicamente che funzionalmente.
12.1.5 Polarità
Come abbiamo visto, l’estremità apicale delle cellule epiteliali rivolta verso la superficie libera
differisce dalla estremità basale rivolta verso il tessuto connettivo. La differenza non è solo
strutturale, ma anche funzionale, ossia la porzione apicale della cellula svolge funzioni diverse dalla
porzione basale.
La differenza morfo-funzionale delle cellule epiteliali è definita polarità. La polarità è
particolarmente evidente nell’epitelio cilindrico semplice, che svolge funzioni di assorbimento,
trasporto e secrezione. Questo epitelio riveste la superficie interna dell’intestino (assorbimento) e
dei tubuli renali (trasporto), e costituisce la componente secernente delle ghiandole esocrine
(secrezione).
Un ruolo fondamentale nel mantenimento della polarità delle cellule epiteliali viene giocato dalle
giunzioni cellulari che si stabiliscono lungo le zone laterali di contatto delle cellule.
12.2 Classificazione degli epiteli di rivestimento
La morfologia e l’organizzazione delle cellule di cui è composto il tessuto epiteliale sono alla base
della sua classificazione. Infatti gli epiteli si classificano in base al numero di strati di cellule che li
compongono ed alla loro forma.
Un epitelio può essere costituito da uno o più strati di cellule, pertanto possiamo distinguere:
•
Epitelio semplice o monostratificato, quando è formato da un solo strato di cellule che
poggiano tutte sulla membrana basale;
•
Epitelio composto o pluristratificato, quando è formato da più strati di cellule, di cui solo lo
strato più interno poggia sulla membrana basale.
In base alla forma delle cellule che lo compongono, l’epitelio può essere suddiviso in:
•
Pavimentoso o squamoso, quando le cellule che lo compongono appaiono appiattite, con la
base maggiore dell’altezza;
•
Cubico, quando la base delle cellule dello strato più superficiale è all’incirca eguale alla
loro altezza;
•
Cilindrico o colonnare, quando l’altezza delle cellule dello strato più superficiale è maggiore
della base.
Negli epiteli monostratificati tutte le cellule poggiano sulla membrana basale. Esse posseggono
quindi una superficie basale ed una estremità apicale libera. Negli epiteli pluristratificati, solo lo
strato più interno di cellule poggia sulla membrana basale. Tali cellule, quindi, posseggono una
superficie basale ma non un’ estremità apicale libera. Nel caso di epiteli pluristratificati, è la forma
delle cellule dello strato cellulare superficiale, cioè quello che presenta la superficie libera, che
viene utilizzata per la classificazione.
Incrociando quindi i due criteri utilizzati per classificare gli epiteli (numero di strati cellulari e
forma delle cellule) , otterremo i seguenti tipi:
epitelio pavimentoso semplice o monostratificato
epitelio cubico semplice o monostratificato
epitelio cilindrico semplice o monostratificato
epitelio pavimentoso composto o pluristratificato
epitelio cubico composto o pluristratificato
epitelio cilindrico composto o pluristratificato
Esistono poi alcuni tipi di epiteli che sfuggono a questa classificazione e che vengono definiti:
epitelio di transizione. Il numero degli strati cellulari e l’aspetto si modificano con lo stato
funzionale (distensione o contrazione) dell’organo.
epitelio pseudo stratificato. E’ formato da un solo strato di cellule che poggiano sulla membrana
basale, ma non tutte posseggono un’estremità apicale libera, poiché non tutte raggiungono la
superficie libera. Ne consegue che all’osservazione al MO l’impressione che se ne ricava è di un
tessuto pluristratificato, poiché i nuclei delle cellule sono disposti a diverse altezze come in un
tipico epitelio pluristratificato, ma tutte le cellule poggiano sulla membrana basale come in un tipico
epitelio monostratificato.
La morfologia di un epitelio si correla alla funzione
Gli epiteli che svolgono funzioni di assorbimento o secrezione sono in genere epiteli semplici o
pseudostratificati. L’altezza delle cellule che li formano è proporzionale all’intensità dell’attività
svolta. Gli epiteli che svolgono funzioni di trasporto transepiteliale di ossigeno, anidride carbonica,
ioni, piccole molecole, etc. sono in genere pavimentosi semplici. L’aumento del numero degli strati
cellulari negli epiteli pluristratificati si correla alla necessità di creare una barriera protettiva contro
traumi meccanici o disidratazione. In questi casi gli strati più esterni possono essere cheratinizzati,
come nell’epidermide, rendendo la funzione protettiva più efficace.
12.2.1 Epitelio pavimentoso semplice
E’ costituito da un singolo strato di cellule dalla forma appiattita, più spesse al centro, in
corrispondenza del nucleo, che può essere ovoidale o sferico, e più schiacciate in periferia. Le
cellule sono tra loro unite da giunzioni cellulari ancoranti e/o occludenti. In visione frontale, le
cellule che compongono questo epitelio ricordano le mattonelle del pavimento. L’epitelio
pavimentoso semplice è largamente rappresentato nel corpo umano. Riveste la superficie interna
degli alveoli polmonari, l’ansa di Henle e la capsula di Bowman dei nefroni, la rete testis del
testicolo, il labirinto membranoso e la parete interna del timpano, le cavità del corpo che non
comunicano con l’esterno (in questo caso prende il nome di mesotelio) ed il lume dei vasi sanguigni
e linfatici (in questo caso prende il nome di endotelio). La parete dei capillari è formata dalle sole
cellule endoteliali rivestite sulla superficie esterna della membrana basale. Immerse nello spessore
della membrana basale sono presenti speciali cellule connettivali chiamate periciti, che hanno
funzione contrattile. Nei capillari di medio calibro due o tre cellule endoteliali delimitano il lume
centrale e sono tra loro connesse da giunzioni ancoranti. In alcuni distretti anatomici, come nei
capillari che irrorano l’encefalo, tra le cellule endoteliali si possono stabilire delle giunzioni
occludenti atte a limitare la diffusione di grosse molecole e di microrganismi come i batteri, dal
sangue al liquido extracellulare del sistema nervoso centrale (SNC), contribuendo alla formazione
della cosiddetta barriera emato-encefalica. Sono presenti inoltre cellule della microglia, astrociti e
neuroni che contribuiscono alla regolazione delle giunzioni strette e delle funzioni della barriera
emato-encefalica. Nei capillari di piccolo calibro una sola cellula endoteliale può delimitare tutta la
circonferenza del vaso. Alcuni capillari hanno la parete formata da cellule endoteliali con lembi
citoplasmatici assottigliati e interrotti per la presenza di pori (o fenestrature) del diametro di 80-100
nm. Essi sono pertanto definiti capillari fenestrati e, al contrario delle cellule endoteliali dei capillari
del SNC, sono più facilmente attraversabili dalle molecole. Capillari fenestrati si trovano nei reni,
nel fegato, nell'intestino tenue, nel pancreas ed in alcuni organi endocrini. Come nei capillari
continui, anche nei capillari fenestrati esiste, sotto l'endotelio, una membrana basale continua, in cui
all'esterno si trovano periciti, che si affondano nella lamina basale stessa, e sottili fasci di fibre
collagene.
12.2.2 Epitelio cubico semplice
E’ formato da un singolo strato di cellule dalla forma cuboide, con il nucleo in posizione centrale.
Non è molto rappresentato nel corpo umano. Lo si ritrova nel rivestimento della superficie
dell’ovario (dove è chiamato epitelio germinativo) e nei condotti escretori di molte ghiandole.
Forma la parete dei follicoli tiroidei, nei quali però può trasformarsi in pavimentoso semplice o
cilindrico semplice al variare dello stato funzionale dell’organo.
12.2.3 Epitelio cilindrico semplice
E’ costituito da cellule dalla forma rettangolare. Il nucleo occupa in genere il terzo inferiore della
cellula. E’ presente in due tipi, ciliato o non ciliato. L’epitelio cilindrico semplice forma i dotti
escretori delle grosse ghiandole. E’ largamente rappresentato a livello dell’apparato digerente,
riproduttore e respiratorio. Infatti riveste la superficie interna dello stomaco, intestino, cistifellea,
utero, tube ovariche (tube di Falloppio) e piccoli bronchi. In relazione alla funzione assorbente
dell’intestino, in cui si completa la digestione degli alimenti e i prodotti finali della digestione
vengono assorbiti e trasferiti ai vasi sanguigni e linfatici, le cellule che formano l’epitelio cilindrico
semplice che tappezza l’intestino (chiamate enterociti) presentano la superficie apicale
caratterizzata da numerosi microvilli, visibili al MO come orletto a spazzola. Il ripiegamento della
membrana plasmatica sulla superficie apicale delle cellule epiteliali aumenta molto la superficie di
assorbimento e può essere quindi ritenuto un adattamento morfologico alla funzione assorbente.
La parete intestinale è formata da quattro strati: mucosa, sottomucosa, tonaca muscolare e sierosa.
La mucosa è lo strato più interno ed è a sua volta costituita da epitelio cilindrico semplice che
poggia su di uno strato di tessuto connettivo denominato lamina o tonaca propria. L’epitelio riveste
e delimita degli avvallamenti sulla superficie dell’intestino, chiamati cripte. La lamina propria, che
sostiene l’epitelio, è costituita da uno strato di tessuto connettivo grazie al quale viene assicurata
l’irrorazione sanguigna ed in cui sono presenti fibre reticolari, elastiche e collagene, macrofagi,
linfociti, plasma cellule e granulociti neutrofili. Al confine tra la mucosa e la sottomucosa è
presente una sottile lamina formata da cellule muscolari lisce definita muscularis mucosae.
L’epitelio intestinale è costituito da quattro principali tipi cellulari: 1) gli enterociti, cellule
cilindriche di notevole altezza, con nucleo ovale basale, citoplasma eosinofilo e superficie apicale
PAS positiva con orletto a spazzola. Gli enterociti posseggono una funzione di assorbimento e
rappresentano la più numerosa categoria di cellule epiteliali intestinali. 2) Le ghiandole unicellulari
o cellule caliciformi, dalla forma ovale o rotondeggiante, con nucleo basale e schiacciato,
citoplasma basofilo, metacromatico e PAS positivo. Le cellule caliciformi secernono una sostanza
di natura mucosa che ha la funzione di proteggere la superficie intestinale da traumi provocati da
agenti esterni e dall’ingresso di microrganismi. 3) Le cellule neuroendocrine o enteroendocrine,
specializzate appunto nella sintesi di ormoni. Posseggono una forma ovale o triangolare, citoplasma
pallido e sono ricche di granulazioni evidenziabili con coloranti a base d’argento. Le cellule
neuroendocrine possono anche essere messe in evidenza in maniera specifica con colorazioni
immunoistochimiche. 4)
Le cellule del Paneth, dalla forma a fiasco e citoplasma granulare
eosinofilo. Le cellule del Paneth contengono zinco, secernono lisozima e fattori di crescita e sono
le principali responsabili della secrezione di peptidi ad azione antimicrobica.
La sottomucosa è uno strato di tessuto connettivo lasso contenente fibroblasti, mastociti, vasi
sanguigni; inoltre tale tessuto connettivo contiene fibre postgangliari del sistema nervoso autonomo
simpatico, cellule gangliari e fibre del sistema nervoso autonomo parasimpatico: l’insieme di queste
fibre costituisce il plesso di Meissner.
Al di sotto della sottomucosa si estenda la tonaca muscolare, consistente in due strati di cellule
muscolari lisce. L’orientamento delle cellule è circolare nello strato più interno e longitudinale nello
strato più esterno. Un prominente plesso nervoso, chiamato plesso mioenterico o di Auerbach, giace
tra i due strati muscolari.
Infine la sierosa viscerale riveste esternamente l’intestino. Essa è composta da mesotelio e da una
sottostante lamina di tessuto connettivo lasso.
L’epitelio cilindrico semplice che riveste l’utero, le tube ovariche e piccoli bronchi presenta sulla
superficie apicale numerose ciglia ed è pertanto definito ciliato. La presenza delle ciglia è da
mettersi in relazione alla funzione di trasporto di materiale lungo l’epitelio. Le ciglia, muovendosi
all’unisono, funzionano come un tappeto trasportatore che consente il movimento di sostanze verso
una determinata direzione. Nel caso dell’epitelio tubarico, l’ovocita espulso dall’ovario viene
trasportato verso l’utero. Nell’apparato respiratorio (albero tracheo-bronchiale), il muco secreto
dalle ghiandole unicellulari frammiste alle cellule epiteliali intrappola polvere, batteri e particelle
presenti nell’aria inspirata, elementi questi che potrebbero danneggiare l’apparato respiratorio e che
vengono così raccolti e convogliati verso la faringe, deglutiti con la saliva e quindi allontanati .
12.2.4 Epitelio pavimentoso pluristratificato
Questo epitelio è formato da numerosi strati di cellule di diversa forma. Lo strato più profondo, a
contatto con la membrana basale, è formato da cellule cubiche o cilindriche che divengono
appiattite man mano che ci si sposta verso la superficie. L’epitelio pavimentoso pluristratificato è
largamente rappresentato nel corpo umano. Riveste la superficie esterna del corpo formando
l’epidermide, cioè lo strato più esterno della pelle o cute. Costituisce il rivestimento della bocca,
esofago e vagina e cornea. Nella pelle le cellule, chiamate cheratinociti, degli strati cellulari
superficiali sono cellule cosiddette “morte”, che hanno perso il nucleo e sono infiltrate di cheratina,
una proteina presente anche nei capelli e nelle unghie. In questo caso l’epitelio viene definito
cheratinizzato o corneificato. La presenza dello strato cellulare cheratinizzato possiede una
funzione protettiva, amplificata dall’estensione dello spessore dello strato di cellule cheratinizzate,
che è massimo nella pianta del piede. Le cellule morte vengono costantemente rimpiazzate dalle
cellule che provengono dagli strati più profondi e sono eliminate sottoforma di piccole scaglie (la
forfora ne è uno “spiacevole” esempio). Il tempo di rinnovamento dell’epidermide, cioè il tempo
necessario perché un cheratinocita si porti dallo strato più profondo a quello superficiale, si aggira
intorno ai 20 giorni. Durante questo processo di rinnovamento cellulare, i cheratinociti subiscono
numerosi cambiamenti morfo-funzionali che verranno descritti qui di seguito. Nelle zone non
esposte all’aria, in cui l’epitelio è umido, come nella bocca, nella vagina, nell’esofago e nella
cornea, le cellule degli strati più esterni non sono infiltrate di cheratina, o lo sono solamente in
minima parte, e non perdono il nucleo, per cui sono cellule vive. In questo caso l’epitelio viene
definito non cheratinizzato o non corneificato.
Nell’epidermide si distinguono quattro strati cellulari denominati, procedendo dall’interno verso
l’esterno, strato basale o germinativo, strato spinoso, strato granuloso e strato corneo. Laddove
l’epidermide si presenta particolarmente spessa, come nel palmo della mano o nella pianta del
piede, tra lo strato granuloso e quello corneo si frappone un ulteriore sottile strato, definito lucido.
Tutti i cheratinociti sono prodotti dalla divisione per mitosi delle cellule dello strato basale e
possono quindi essere definiti come cheratinociti staminali (uni potenti). Dai cheratinociti staminali
hanno origine i cheratinociti che si avviano verso il processo di differenziamento necessario ad una
migrazione verso lo strato più esterno ed i cheratinociti che rimangono nello strato basale con
funzione staminale. I cheratinociti dello strato basale aderiscono alla membrana basale attraverso la
presenza di emidesmosomi e sono connessi tra loro attraverso desmosomi. L'adesione alla
membrana basale è mediata dall'integrina α6β4, mentre le adesioni sulla superficie apicale e laterale
sono mediate dalle integrine α2β1 α3β1. Il citoscheletro è formato da filamenti di cheratina, o
tonofilamenti, disposti parallelamente all’asse lungo della cellula. Altri due tipi cellulari si trovano
nello strato basale: le cellule di Merkel ed i melanociti. Le cellule di Merkel, che funzionano come
recettori, hanno il compito di inviare attraverso i nervi sensitivi somatici le informazioni di
sensibilità tattile al cervello. Le cellule di Merkel sono particolarmente abbondanti sulla superficie
di mani e piedi. I melanociti producono un pigmento chiamato melanina e contenuto all’interno di
granuli provvisti di membrana plasmatica. La melanina conferisce il colore alla pelle ed ai capelli
ed aiuta le cellule vive dell’epidermide a difendersi dagli effetti dannosi dei raggi ultravioletti. La
melanina prodotta dai melanociti viene ceduta ai cheratinociti in via di differenziamento e quindi
trasportata verso gli strati più esterni dell’epidermide, dove viene eliminata con le cellule morte.
Al di sopra dello strato basale si trova lo strato spinoso, formato da una decina di strati cellulari e
così definito per via dell’apparenza “spinosa” dei cheratinociti, apparenza dovuta alla presenza di
prolungamenti cellulari che si connettono con i prolungamenti cellulari di altri cheratinociti
attraverso desmosomi e conferiscono all’epidermide, oltre all’aspetto spinoso, forza e resistenza alla
trazione meccanica. Le cellule dello strato spinoso sintetizzano proteine, come l'involucrina, che
tendono ad accumularsi sul versante citoplasmatico della membrana cellulare, costituendo
l'involucro cellulare corneificato. I cheratinociti dello strato spinoso posseggono due tipi di granuli:
i melanosomi, che contengono appunto la melanina prodotta dai melanociti dello strato basale, ed i
cheratinosomi o corpi lamellari, che contengono materiale lipidico rilasciato nelle regioni più
superficiali dello strato granuloso e contribuisce a fornire una barriera impermeabile. Interdisperso
tra i cheratinociti dello strato spinoso, si trovano alcune cellule dendritiche chiamate cellule di
Langerhans, che hanno funzione difensiva. Esse agiscono infatti in maniera simile ai macrofagi,
ingerendo e distruggendo batteri, particelle estranee e cellule danneggiate.
Al di sopra dello strato spinoso si trova lo strato granuloso, formato da 3 a 5 strati di cheratinociti
appiattiti. Le cellule hanno un aspetto appunto granuloso, dovuto alla presenza nel citoplasma di
grandi quantità di granuli privi di membrana, costituiti da cheratina e cheratoialina. I cheratinociti di
questo strato, come quelli dello strato spinoso, contengono cheratinosomi, il cui contenuto lipidico
viene esocitato nello spazio extracellulare, rendendo la pelle relativamente impermeabile alla
disidratazione.
Lo strato lucido ha un aspetto traslucido, a cui deve il nome, ed è localizzato tra lo strato granuloso
e quello corneo, ma solo nell’epidermide del palmo della mano e del piede. I cheratinociti che lo
compongono sono cellule morte, dalla forma schiacciata. I cheratinociti dello strato lucido sono
infarciti di eleidina, una lipoproteina contente zolfo e derivata dalla cheratoialina, che fornisce
l’aspetto traslucido e costituisce un’efficiente protezione contro la disidratazione.
Negli epiteli pavimentosi composti cheratinizzati, lo strato corneo è lo strato più superficiale,
quello esposto all’esterno del corpo. L’intensa cheratinizzazione (anche definita corneificazione) ne
conferisce il nome. Nell’epidermide umana questo strato è formato da un numero di strati di
cheratinociti variabile tra i 15 e i 30. Lo strato corneo è asciutto, formato da cellule morte, in grado
di prevenire la penetrazione di batteri e microrganismi, e di agire contro la disidratazione dei tessuti
sottostanti, oltre a fornire una protezione meccanica contro abrasioni e traumi. I cheratinociti di
questo strato vengono periodicamente eliminati e sostituiti dalle cellule degli strati sottostanti.
L’intero strato corneo viene sostituto nel giro di 4 settimane. Alcune procedure cosmetiche, come
la microdermoabrasione, rimuovono delicatamente le cellule più esterne dello strato corneo,
conferendo alla pelle un aspetto più fresco e giovanile.
12.2.5 Epitelio cubico e cilindrico pluristratificato.
Sia l’epitelio cubico pluristratificato che quello cilindrico pluristratificato sono molto rari nel corpo
umano. Li troviamo soprattutto lungo i dotti escretori delle grosse ghiandole esocrine e nella
mucosa della laringe e della faringe. Una varietà ciliata di epitelio cilindrico pluristratificato la si
ritrova nella mucosa della faccia posteriore dell’epiglottide.
12.2.6 Epitelio di transizione
L’epitelio di transizione è definito tale poiché possiede l’importante caratteristica di potersi adattare
alle variazioni di ampiezza della superficie che riveste. Questo tipo di epitelio è tipicamente
presente nelle vie urinarie, dove riveste la superficie interna della vescica, dell’uretere e della parte
superiore dell’uretra. Quando la vescica è vuota il volume interno è ridotto, mentre quando si
riempie di urina avviene una dilatazione che provoca l’aumento del volume interno e di
conseguenza della superficie. L’epitelio di transizione segue queste variazioni, apparendo come un
epitelio pluristratificato quando la vescica è vuota, mentre lo spessore dell’epitelio si riduce ed il
numero degli strati diminuisce, poiché le cellule scivolano le una sulle altre appiattendosi, quando la
vescica è piena.
12.2.7 Epitelio pseudo stratificato
Questo tipo di epitelio è formato da un unico strato di cellule che poggiano tutte sulla membrana
basale, ma solo alcune raggiungono la superficie libera. I nuclei sono disposti ad altezze diverse e
pertanto al MO esso appare illusoriamente come un epitelio pluristratificato. E’ presente lungo le
vie genitali maschili (uretra, epididimo e deferente) e nei condotti escretori di alcune ghiandole.
Nell’apparato respiratorio l’epitelio pseudo stratificato è ciliato.