1
INDICE ALLEGATO [ 6 ]
CENNI SU:
Gli Organuli Cellulari
• IL SISTEMA DI
ENDOMEMBRANE:
• Il Reticolo Endoplasmatico
Liscio e Rugoso
• L’apparato di Golgi
• I Lisosomi e i Perossisomi
• I Mitocondri
• Esocitosi Endocitosi Fagocitosi
Il Cloroplasto e la fotosintesi
clorofilliana
1
2
Inizio allegato [ 6 ]
DENTRO LA CELLULA
La cellula
Introduzione
La cellula è l’unità fondamentale di ogni organismo vivente; le sue
principali caratteristiche sono di essere in grado di vivere
autonomamente e soprattutto di riprodursi.Se pensiamo
all’evoluzione della vita sulla Terra, possiamo considerare la
cellula come la struttura all’interno della quale i processi chimicofisici che hanno dato origine alle molecole biologiche, e che sono
alla base delle attività vitali, sono potuti avvenire in modo
regolare e ordinato. La capacità di produrre reazioni ordinate e
regolari è stata necessaria affinché tali reazioni potessero ripetersi
in maniera identica, fornendo un primo esempio di fenomeno
riproducibile.Vi sono due tipi di cellule: le procariote, con una
struttura molto semplice, senza nucleo; e le eucariote, nelle quali
il materiale genetico all’interno della cellula è raccolto in un
nucleo, racchiuso in un involucro che lo separa dalla restante
parte, detta citoplasma. Sono eucariote tutte le cellule animali e
vegetali. Ci riferiremo a un eucariote quindi come a una “cellula
tipo" con caratteristiche funzioni fondamentali comuni a tutte le
cellule umane.Durante l’evoluzione della vita, l’associazione e la
cooperazione fra cellule è risultata vantaggiosa, dando origine a
organismi a più cellule. Da qui la necessità per le cellule di
differenziarsi per svolgere funzioni diverse; all’interno dello stesso
organismo troveremo quindi cellule molto diverse fra loro come
struttura, dimensioni e forma a seconda delle funzioni che
svolgono.Tutte queste cellule sono però riferibili al modello
generale della cellula eucariote. Prima di descrivere in particolare
la struttura e le funzioni della cellula tipo passeremo in rassegna
le basi chimiche della vita, cioè gli elementi e le sostanze chimiche
che costituiscono la materia vivente e quindi il nostro prototipo
cellulare.
2
3
Struttura e organizzazione della cellula
Introduzione
La cellula è l’unità fondamentale degli organismi viventi, ed è in grado di vivere in maniera
autonoma e di riprodursi. Nonostante la sua autonomia, la cellula in realtà non è in grado di
sopravvivere al di fuori del suo ambiente, a meno che non venga coltivata artificialmente in
speciali terreni di coltura. Questo perché ogni cellula si è adattata alle condizioni ambientali in
cui si trova e si è specializzata, negli organismi superiori (come quello umano), a svolgere
determinate funzioni necessarie all’organismo intero, perdendo la capacità di sopravvivenza
autonoma. Per esempio, una cellula muscolare è in grado di contrarsi conferendo a un
organismo la capacità di movimento, ma non è assolutamente in grado di difendersi da
un’infezione virale, al contrario di un linfocito. Quindi due cellule di questo tipo, isolate
dall’organismo, non sono in grado di sopravvivere, mentre all’interno di esso, dove sfruttano le
condizioni di cooperazione con le altre cellule, trovano un ambiente ideale.
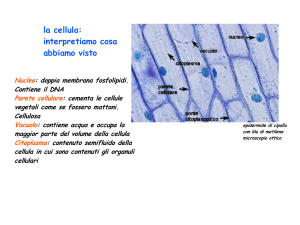
La cellula eucariote ha un volume maggiore rispetto a quella procariotica e contiene una serie
di strutture dette organelli, o organuli. Una caratteristica delle cellule è di avere una
membrana, detta plasmatica o cellulare, che le delimita rispetto all’ambiente esterno e
conferisce loro identità. Vi è poi il materiale genetico (DNA) organizzato in cromosomi e
contenuto nel nucleo, che è una struttura anch’essa delimitata da una membrana nucleare.
Il rimanente contenuto cellulare è detto citoplasma: nel suo interno si trovano organelli, come i
mitocondri o i centrioli, immersi in una sostanza gelatinosa, il citosol Il citoplasma è separato
in differenti scomparti da un sistema di membrane interne che delimitano strutture con
funzioni specializzate come il reticolo endoplasmatico, l'apparato del Golgi, lisosomi e
perossisomi. Vediamo ora in dettaglio quali sono e come sono organizzate le funzioni delle
strutture che costituiscono la cellula.
Organulo
All'interno della cellula eucariota sono presenti vari organelli od organuli, che svolgono differenti
funzioni necessarie alla sua sopravvivenza. Gli organelli, assieme al citosol, formano il citoplasma
Durante la mitosi vengono divisi in modo sommario fra le due cellule figlie, perché i mitocondri e i
cloroplasti si duplicano da soli. Essi costituiscono ulteriore differenza fra le cellule. Le procariote
non possiedono altri organelli che i ribosomi. Le cellule animali possiedono il reticolo
endoplasmatico (che sostiene la cellula, mentre le cellule vegetali non ne hanno bisogno, perché
possiedono la parete cellulare che svolge già questa funzione), i lisosomi mentre sono prive del
vacuolo vero e proprio, mentre possiedono introflessioni della membrana omonime e dalla
medesima funzione.
Le teorie sulla nascita di alcuni organelli sono varie e controverse. Una molto accreditata sostiene
che si tratti di organismi a sé stante fagocitati dalle cellule in tempi preistorici che hanno cominciato
a vivere in simbiosi con loro, fino a diventarne parte. A favore di questa teoria c'è il fatto che molti
organelli (ad esempio i cloroplasti e i mitocondri) sono in grado di dividersi autonomamente.
3
4
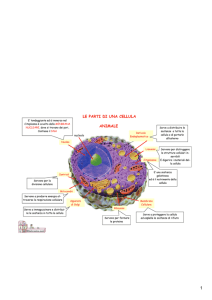
Gli organuli in figura:
1. Nucleolo
2. Nucleo
3. Ribosoma
4. Vescicola
5. Reticolo endoplasmatico rugoso
6. Apparato del Golgi
7. Citoscheletro
8. Reticolo endoplasmatico liscio
9. Mitocondrio
10. Vacuolo (nelle cellule vegetali e nei funghi)
11. Citoplasma
12. Lisosoma
13. Centriolo
Altri organuli:
Endosoma
Perossisoma nelle cellule vegetali: Cloroplasto
4
5
. Gli organelli del sistema di endomembrane svolgono funzioni specifiche: il r.e. e app. di golgi
provvedono alla sintesi e allo smistamento di proteine e lipidi ad altri organelli o all’esterno
della cellelua.Gli endosomo e i lisosomi sono destinati alla raccolta,degradazione e digestione
di particelle e sostanze importate nelle cellule dall’ambiente esterno. Le vescicole membranose
sono addette al trasporto di sostanze di origine intra ed extra cellulare verso le rispettive
destinazioni.
Ribosomi e reticolo endoplasmatico
I ribosomi sono gli organelli cellulari più numerosi; sono costituiti da due subunità, una
maggiore e una minore, che si dissociano alla fine di ogni ciclo di sintesi di una proteina.
Chimicamente, i ribosomi sono ribonucleoproteine, cioè strutture costituite da nucleotidi e
proteine. Le grosse molecole di RNA ribosomiale (rRNA) formano un’impalcatura sulla quale si
organizzano spontaneamente decine di tipi di proteine ribosomiche.
Ogni proteina viene sintetizzata attraverso un processo in cui molecole di RNA messaggero
escono dal nucleo e si attaccano ai ribosomi, in cui la sequenza di RNA viene tradotta nella
corrispondente sequenza di aminoacidi assemblati a formare la proteina. Il ribosoma è quindi
un apparecchio per sintetizzare le proteine, in grado di riunire nella disposizione opportuna le
molecole necessarie per la reazione di sintesi; parte delle proteine che lo costituiscono inoltre
funzionano da enzimi che catalizzano le stesse reazioni. I ribosomi dispersi nel citosol in
genere sintetizzano proteine che restano e sono utilizzate nella cellula stessa, anche se
passano dal citosol a organuli come il nucleo o i mitocondri. Molte proteine, una volta
sintetizzate, si staccano dal ribosoma e nel citosol subiscono modificazioni chimiche che le
portano ad avere le caratteristiche e la conformazione finale della proteina.
Le proteine che devono essere secrete all’esterno dalla cellula (come gli enzimi digestivi o gli
ormoni), o quelle che devono inserirsi nella membrana cellulare o nelle membrane dei
differenti organuli, sono sintetizzate da ribosomi che si trovano legati a un complesso sistema
di membrane interne, il reticolo endoplasmatico. Questo è una struttura dinamica, la cui
quantità aumenta o diminuisce a seconda dell’attività cellulare: la sua estensione rappresenta
in media circa la metà di tutte le membrane cellulari. Le membrane del reticolo endoplasmatico
formano una rete intrecciata di sacchetti appiattiti (detti cisterne), tubi e canalicoli, attraverso i
quali le sostanze possono passare. Lo spazio interno del reticolo endoplasmatico delimitato
dalle membrane è detto lume del reticolo.
Nelle cellule si possono distinguere due regioni funzionalmente e strutturalmente diverse: il
reticolo endoplasmatico ruvido (o granulare), al quale, sul lato citoplasmatico della membrana,
sono attaccati i ribosomi (questo tipo di reticolo è interessato sia alla sintesi sia al trasporto
fuori dalla cellula delle proteine); il reticolo endoplasmatico liscio, che fisicamente è una
porzione dello stesso reticolo endoplasmatico ruvido, ma è privo di ribosomi. Il reticolo
endoplasmatico liscio è importante nella sintesi dei lipidi e nel trasporto di sostanze dal reticolo
endoplasmatico ruvido all’apparato del Golgi. Mentre il reticolo endoplasmatico ruvido è
organizzato esclusivamente a cisterne, quello liscio è una rete tridimensionale di tubuli.
I ribosomi legati al reticolo endoplasmatico ruvido, che sintetizzano le proteine, si trovano
normalmente liberi nel citosol e si attaccano sulla membrana esterna del reticolo dopo aver
riconosciuto una proteina che serve da segnale e recettore. La presenza di recettori permette
alle proteine idrosolubili, che vengono sintetizzate nella porzione del ribosoma esposto al
citosol, di passare all’interno del lume del reticolo endoplasmatico ruvido, superando la barriera
del doppio strato lipidico della membrana del reticolo. Le proteine passano nel lume del reticolo
endoplasmatico ruvido non appena sono sintetizzate e prima di avere la possibilità di ripiegarsi
nella loro conformazione finale. Le proteine di secrezione vengono liberate nel lume, mentre
quelle che devono essere esposte sulle membrane rimangono ancorate alla membrana del
reticolo; in seguito, mediante un sistema di vescicole, verranno trasportate a destinazione: là
le vescicole si fonderanno con le membrane esponendo le proteine all’esterno.
5
6
All’interno del reticolo endoplasmatico ruvido la maggior parte delle proteine viene glicosilata,
ovvero vengono aggiunte alle catene proteiche delle catene di zuccheri, che le trasformano in
glicoproteine. La glicosilazione è una delle principali funzioni del reticolo endoplasmatico ruvido.
Le catene di zuccheri aggiunte alla proteina sono fornite da carboidrati legati a lipidi della
membrana del reticolo e vengono in seguito modificate nell’apparato del Golgi.
Il reticolo endoplasmatico liscio è la “fabbrica” dove vengono prodotte le nuove membrane per
tutta la cellula: infatti al suo interno sono sintetizzati tutti i lipidi che costituiscono le
membrane, i quali si associano con le proteine provenienti dal reticolo endoplasmatico ruvido,
con cui vi è continuità. A questo punto dei processi vi sono tutti i componenti per formare le
nuove membrane. Dalle estremità del reticolo endoplasmatico liscio si staccano delle vescicole,
dette vescicole di trasporto, che vanno a fondersi con la membrana specifica dell’organulo
bersaglio, a seconda del contenuto della vescicola. La vescicola diventa parte integrante della
membrana dell’organulo bersaglio e i suoi contenuti vengono secreti all’esterno, o esposti sulle
membrane se sono ancorati a esse.
Apparato del Golgi
L'apparato del Golgi è formato da numerosi gruppi di
cisterne appiattite impilate una sull'altra e circondate da
tubuli e vescicole.
L’apparato del Golgi si trova in genere vicino al nucleo ed è formato da numerosi gruppi di
cisterne appiattite, delimitate da membrane, impilate una sull’altra e circondate da tubuli e
vescicole. Esso ha due facce distinte: una di formazione, o cis, che è strutturalmente associata
con la porzione liscia del reticolo endoplasmatico; e una di maturazione, o trans, che è quella
rivolta verso la membrana cellulare dalla quale gemmano grosse vescicole di secrezione.
La funzione dell’apparato del Golgi è di indirizzare il traffico delle molecole appena sintetizzate
6
7
verso le giuste destinazioni, dopo aver operato le modificazioni necessarie a ottenere la
conformazione definitiva delle varie molecole. In particolare, le catene di zuccheri
precedentemente legate alle proteine nel reticolo endoplasmatico vengono ampiamente
modificate con l’aggiunta o l’asportazione di determinati residui di carboidrati. Riassumendo,
vediamo il percorso che una proteina di secrezione o di membrana deve compiere all’interno di
una cellula. La proteina viene sintetizzata a livello dei ribosomi collegati al reticolo
endoplasmatico ruvido, e passa all’interno del lume del reticolo dove le vengono legate le
catene di carboidrati. Da qui passa nel reticolo endoplasmatico liscio, dove, tramite vescicole
che si generano dal reticolo stesso, viene trasferita alla faccia cis dell’apparato del Golgi. Le
vescicole si fondono con le cisterne dell’apparato del Golgi e la proteina si trova nel lume di
questo organulo dove subisce in vario modo modificazioni della struttura e dei componenti
glicidici. Tali modificazioni avvengono seguendo un percorso all’interno delle cisterne, e
portano infine la proteina, che ha assunto la sua struttura definitiva, verso la faccia trans
dell’apparato del Golgi. Da qui le proteine vengono allontanate in modo selettivo, tramite
vescicole, e raggiungono la loro destinazione “operativa”: sono secrete negli spazi
extracellulari, o diventano parte integrante della componente proteica della membrana
cellulare o degli organuli cellulari.
Lisosomi e perossisomi
I lisosomi, vescicole che si formano nell’apparato del Golgi, sono essenzialmente sacchetti
membranosi (delimitati da una membrana a doppio strato lipidico) che racchiudono enzimi
idrolitici, in grado, nella digestione intracellulare, di tagliare in maniera specifica e controllata
grosse molecole. Gli enzimi idrolitici dei lisosomi sono detti idrolasi acide e comprendono
proteasi (che tagliano le proteine), nucleasi (che tagliano gli acidi nucleici), glicosidasi, lipasi
ecc. Se i lisosomi si rompono, la cellula stessa viene distrutta, poiché gli enzimi che essi
contengono sono capaci di scindere tutti i composti principali presenti nella cellula.
I lisosomi contengono ognuno idrolasi diverse, che svolgono l’ampia serie di differenti funzioni
digestive. Le idrolasi dei lisosomi sono sintetizzate nel reticolo endoplasmatico, come le altre
proteine, e trasferite in seguito nel lume dell’apparato del Golgi. I lisosomi si formano per
gemmazione (si staccano come goccioline di membrana) dalle cisterne più esterne
dell’apparato del Golgi. Bisogna distinguere fra due classi generali di lisosomi: i lisosomi
primari, appena formati e non ancora fusi con altre vescicole contenenti i materiali da digerire;
e i lisosomi secondari, che derivano da ripetute fusioni di lisosomi con altre vescicole. In
questo secondo tipo sono contenuti enzimi, materiale da digerire e materiale digerito.
Poiché gli enzimi contenuti nel lisosoma sono potenzialmente pericolosi, essi non possono
essere rilasciati nel citoplasma ma debbono sempre essere confinati all'interno di strutture
membranose in grado di contenerli. Il meccanismo con cui tali enzimi entrano in contatto con
le sostanze da degradare viene definito autofagìa o fagocitosi a seconda dell'origine di tali
sostanze.
Per autofagia si intende la degradazione di componenti della cellula stessa, siano essi
macromolecole o organelli destinati ad essere eliminati ,vengono avvolti da membrana
del reticolo endoplasmatico (autofagosoma)e digeriti da enzimi lisosomiali
Nella fagocitosi, grosse particelle (come un batterio o un virus) vengono inglobate (fagocitate)
dalla cellula,questa volta derivante dalla membrana cellulare (fagosoma), si fonde col lisosoma
formando un cosiddetto (fagolisosoma) dove il suo contenuto viene digerito
7
8
La fagocitosi è la capacità posseduta da diverse cellule di ingerire materiali
estranei e di distruggerli
.
Fagocitosi Processo di ingestione cellulare di particelle o sostanze estranee (dal greco,
phago, "mangiare" e kytos, "cellula"), tramite l'emissione di prolungamenti citoplasmatici. La
fagocitosi è attuata da tutti gli organismi viventi unicellulari e, negli organismi pluricellulari, da
cellule chiamate fagociti, specializzate nella difesa dell'organismo dagli invasori potenzialmente
dannosi.
È una modalità con cui la cellula "mangia" un qualcosa di grosso che sta all'esterno. La fagocitosi,
per portare materiale all'interno della cellula, richiede da parte della cellula stessa, l’emissione di
espansioni citoplasmatiche delimitate da membrana, chiamate pseudopodi, avente quest'ultima
un’impalcatura esterna formata da filamenti di actina che avvolge completamente il batterio
portandolo al suo interno.
Sia la endocitosi che la fagocitosi prevedono la deformazione del plasmalemma, con formazione di
una vescicola contenente il materiale che doveva essere internalizzato; tale formazione vescicolare
verrà indicata, rispettivamente, come endosoma o fagosoma.
Una variante della fagocitosi è l'autofagocitosi, una modalità con cui la cellula decide di degradare
dei suoi organuli per rinnovarli (dato che niente è opportuno scartare e/o buttare): avvolge
l’organulo con membrane del suo reticolo liscio. Quello che si forma in seguito sarà una grossa
vescicola chiamata autofagosoma.
Nell'uomo e negli altri animali superiori i fagociti sono cellule ameboidi che si trovano in tutti i
distretti del corpo. I fagociti di dimensioni maggiori, detti macrofagi, esercitano un ruolo
particolarmente importante nel sistema linfatico, nel fegato e nella milza. Fagociti più piccoli,
chiamati granulociti, derivano dal differenziamento di un tipo di globuli bianchi e vengono
trasportati in tutto l'organismo dalla circolazione sanguigna. Vengono attratti nei siti di infezione da
sostanze chimiche emesse dai microrganismi invasori, per raggiungere i quali sono anche in grado
di attraversare le pareti dei vasi sanguigni. Il successo della loro azione di fagocitosi dipende dalla
8
9
natura del corpo estraneo. Alcune proteine presenti nel sangue, ad esempio gli anticorpi,
normalmente rivestono le particelle estranee e attirano i fagociti che aderiscono al complesso
proteina-particella estranea e lo inglobano.
Rilevanza clinica
Diverse patologie, molto spesso ereditarie molto gravi, sono state associate a
malfunzionamenti dei lisosomi, dei loro enzimi o dei processi portanti alla loro formazione.
Queste malattie sono dovute alla mancanza di particolari proteine litiche, che porta
all'accumulo dei loro substrati all'interno della cellula, con conseguenti problemi nel
metabolismo cellulare.
Ad oggi si conoscono più di 40 diverse malattie genetiche causate dalla deficienza di questi
enzimi. Queste malattie, generalmente note col nome di Lysosomal Storage Diseases
(letteralmente malattie di immagazzinamento lisosomale) malattie dovute al mancato
trasporto degli enzimi lisosomiali, in cui alcuni enzimi non vengono trasportati correttamente
nei lisosomi e quindi non riescono ad assolvere la loro funzione
.
L'endosoma è un corpicciolo vescicolare, presente nella cellula, il cui compito deputato
è quello di partecipare all'endocitosi, ovvero al meccanismo cellulare che permette il transito
attraverso la membrana di macromolecole e corpuscoli, le cui dimensioni non consentono l'ingresso
attraverso i meccanismi del trasporto di membrana
9
10
.
La struttura e le funzioni della membrana plasmatica sono comuni a quelle delle membrane
intracellulari, come ad esempio la membrana nucleare.
Infine, la membrana cellulare partecipa a funzioni complesse: esocitosi (secrezione), endocitosi
(ingestione di sostanze esterne mediante la formazione di vescicole),
L'endocitosi è un processo riguardante la periferia cellulare, attraverso il quale la cellula
internalizza molecole o corpuscoli presenti nello spazio extracellulare in maniera massiva tramite
la modificazione della forma della sua membrana plasmatica, che si invagina per racchiudere il
materiale da introdurre nella cellula in una vescicola, detta "vescicola endocitica". Questa
vescicola viene quindi convogliata nel citoplasma tramite microtubuli
L'esocitosi è il processo cellulare con il quale la cellula riversa al suo esterno delle molecole
accumulate all'interno di una vescicola, tramite la fusione di quest'ultima con la membrana
plasmatica
Nella cellula si trovano altri organuli simili ai lisosomi, i perossisomi, che ne differiscono per il
contenuto enzimatico. I perossisomi contengono le catalasi, enzimi specializzati per
effettuare reazioni ossidative, che impiegano l’ossigeno molecolare. Il perossisoma, con il
mitocondrio, è il principale sito di utilizzazione dell’ossigeno nella cellula. Grossi perossisomi
delle cellule del fegato e del rene sono importanti nel neutralizzare la tossicità di numerose
molecole, ossidandole (per esempio, quasi la metà dell’alcol che beviamo viene neutralizzata
nei perossisomi).
I perossisomi esercitano molte azioni che vanno dall’ossidazione degli acidi grassi a lunga
catena (detta beta-ossidazione), alla sintesi del colesterolo e degli acidi biliari nelle cellule
epatiche, al metabolismo degli amminoacidi e delle purine ed allo smaltimento dei composti
metabolici tossici.
10
11
.
.
.
Mitocondri
I mitocondri sono le "centrali energetiche" della cellula; producono l'energia necessaria per
molte funzioni cellulari, quali il movimento, il trasporto di sostanze ecc. Essi contengono gli
enzimi necessari per far avvenire le reazioni chimiche che recuperano l'energia contenuta negli
alimenti e l'accumulano in speciali molecole di adenosintrifosfato (ATP), nelle quali si conserva
concentrata e pronta all'uso. I mitocondri sono organuli generalmente a bastoncello, ma
possono avere anche forma granulare o filamentosa. Questi organuli sono numerosi all'interno
di una cellula, ma la loro quantità può variare molto a seconda dei tessuti presi in esame: per
esempio, sono estremamente numerosi nelle cellule, come quelle renali o muscolari, in cui vi è
11
12
un continuo e grande consumo di energia.
Il mitocondrio, come il nucleo, è delimitato da due membrane a doppio strato lipidico,
selettivamente permeabili. La membrana esterna è liscia; quella interna forma numerose
pieghe, dette creste, perpendicolari alla parete e più o meno lunghe. Le due membrane
racchiudono e definiscono due spazi: lo spazio intermembrana, che si trova tra le due
membrane, e lo spazio della matrice, dove è presente del materiale omogeneo contenente
enzimi, coenzimi, acqua, fosfati e altre molecole. La membrana esterna è molto più permeabile
di quella interna: contiene infatti molte copie di una proteina, che nel suo spessore dà origine a
grandi canali, permettendo il passaggio non selettivo di tutte le piccole molecole presenti nel
citosol, le quali vanno a riempire lo spazio tra le due membrane. La membrana interna,
viceversa, ha una permeabilità molto selettiva: tramite proteine di trasporto, vi passano solo le
piccole molecole che devono essere metabolizzate nello spazio della matrice. La membrana
interna, lungo le sue creste, contiene una serie di proteine enzimatiche che funzionano in
modo sequenziale, vale a dire che il prodotto di una reazione catalizzata da una proteina
servirà da substrato per l'enzima successivo. Queste sono le proteine della catena di trasporto
degli elettroni, essenziali per la produzione di molecole "ad alta energia".
Il compito dei mitocondri è quello di completare la demolizione delle molecole ingerite come
fonte di energia. Infatti, nel citosol gli zuccheri vengono demoliti con reazioni che non
utilizzano ossigeno, per cui la digestione è parziale e la resa in energia bassa. Nei mitocondri il
metabolismo degli zuccheri (ma anche quello dei lipidi) si completa con la loro ossidazione
(ciclo di Krebs). I prodotti di questa reazione vengono utilizzati dalla catena di trasporto degli
elettroni per produrre molecole ad alta energia (ATP). In questo modo, l'energia
immagazzinata nelle molecole di ATP è molto più alta: infatti da ogni molecola di glucosio
vengono prodotte 36 molecole di ATP, mentre la glicolisi a livello del citosol ne produce
soltanto 2.
I mitocondri hanno un'altra importante caratteristica: sono la sola struttura della cellula, oltre
al nucleo, che contiene materiale genetico (DNA). Il DNA dei mitocondri è relativamente scarso,
paragonabile alla quantità che troviamo nei virus, ed è costituito da molecole circolari,
organizzate in aggruppamenti distinti nella matrice del mitocondrio, ancorate alla membrana
interna. Il DNA mitocondriale reca il codice genetico necessario per la sintesi di alcune proteine
che si trovano esclusivamente all'interno del mitocondrio stesso e, poiché la membrana interna
è impermeabile alla maggior parte delle molecole, queste proteine non vengono mai rilasciate
nel citosol. Queste però non sono che una piccola parte delle proteine presenti nel mitocondrio:
le restanti vengono sintetizzate nel citosol e in seguito trasferite all'organulo. Si crea un flusso
unidirezionale di molecole dal citosol al mitocondrio.
Non essendovi scambio di sostanze dal mitocondrio al citoplasma, l'organulo deve possedere,
ed essere in grado di far funzionare, tutti i sistemi necessari per la sintesi delle proteine
codificate dal proprio DNA. Al momento della duplicazione della cellula, anche i mitocondri, a
differenza di altri organuli che vengono prodotti ex novo, si duplicano. La duplicazione avviene
per scissione (si forma una strozzatura nell'organulo, che poi si divide in due), ed è preceduta
da un accrescimento dell'organulo: proteine e lipidi, sintetizzati a livello del citoplasma,
vengono aggiunti all'organulo in grande quantità. Nel frattempo vi è la duplicazione del DNA
mitocondriale, che risulterà così suddiviso nei due mitocondri generati. Gli organuli completi
verranno poi ereditati dalle cellule figlie, casualmente, metà in una cellula e metà nell'altra,
secondo quella che viene definita eredità non mendeliana o citoplasmatica.
12
13
13
14
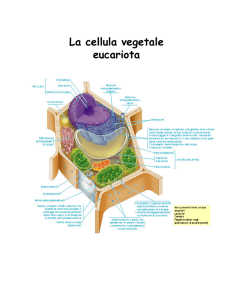
Le cellule vegetali non sono molto differenti da quelle animali. Le differenze risiedono nella
presenza o meno di alcuni elementi nell'uno o nell'altro tipo di cellula.
La cellula vegetale possiede: parete cellulare, membrana cellulare, nucleo e nucleolo, molte
molecole di DNA, ribosomi, reticolo endoplasmatico, apparato di Golgi, un unico lisosoma,
mitocondri, cloroplasti, plastidi diversi dai cloroplasti, vacuolo centrale, citoscheletro.
La cellula vegetale non possiede: lisosomi, centrioli (nella maggior parte di esse).
La cellula animale possiede: membrana cellulare, nucleo e nucleolo, molte molecole di
DNA, ribosomi, reticolo endoplasmatico, apparato di Golgi, lisosomi, mitocondri, centrioli.
La cellula animale non possiede: parete cellulare, cloroplasti, aminoplasti, plastidi diversi
dai cloroplasti, vacuolo centrale.
PLASTIDI
I plastidi sono organuli esclusivi della cellula vegetale, con una zona centrale (stroma) delimitata da
doppia membrana, specializzati in funzioni differenti, situati nel citoplasma; i plastidi
maggiormente specializzati sono i cloroplasti. Questi hanno caratteri comuni con i mitocondri
(doppia membrana, sviluppo eccezionale della superficie interna di membrana, ribosomi 70S, DNA
e RNA, capacità di dividersi per scissione e di essere trasmessi al momento della divisione) e pure
per essi è stata ipotizzata un’origine simbiontica.
Esistono diversi tipi di plastidi per forma e funzioni, tuttavia essi derivano tutti dai proplastidi e in
differenti condizioni possono modificarsi da un tipo all’altro. Essi possono essere classificati nel
modo seguente:
• proplastidi (plastidi immaturi)
• plastidi pigmentati
• fotosintetizzanti
• cloroplasti
• rodoplasti
• feoplasti
• non fotosintetizzanti
• cromoplasti
• plastidi non pigmentati (leucoplasti)
• amiloplasti
• elaioplasti
• proteoplasti
• ezioplasti (plastidi anomali)
Esistono rapporti strettissimi fra i diversi tipi di plastidi; tutti possono formare pigmenti ed amido.
Le modificazioni da una classe all’altra sono determinate da fattori genetici, da fattori ambientali (in
particolare presenza o meno di luce, disponibilità di sali), da variazioni interne all’organismo
(maturazione dei frutti, caduta delle foglie, ecc.). Il tipo di plastidio è collegato anche alla
localizzazione nell’ambito della pianta: in parti superficiali esposte alla luce (foglie) prevalgono i
cloroplasti, in parti più interne (radici, fusto) prevalgono gli amiloplasti. L’insieme dei plastidi di
una pianta è definito plastidioma.
14
15
I proplastidi sono piccoli organuli scarsamente differenziati di 0,5-1 μm, presenti nelle
cellule meristematiche; rappresentano i precursori di tutti i plastidi. Sono già presenti nei gameti,
ma in particolare, nelle piante più evolute con gameti maschili ridotti, sono trasmessi solo dai
gameti femminili,
che hanno dimensioni atte a contenerli. Lo stroma è privo di strutture lamellari, contengono
protoclorofilla (precursore della clorofilla), DNA, RNA e ribosomi 70S; sono in grado di svolgere
la sintesi proteica necessaria ad evolvere nei plastidi più evoluti.
IL VACUOLO
Il vacuolo non è un vero e proprio organulo, ma un compartimento della cellula vegetale,
rappresentato da una cisterna tondeggiante ripiena di un succo vacuolare e delimitata da una
membrana, chiamata tonoplasto. Nelle cellule giovani i vacuoli sono piccoli e numerosi, mentre nel
corso del differenziamento il numero diminuisce e nella cellula adulta vi è, nella maggior parte dei
casi, un unico grosso vacuolo, che occupa 80-90% del volume cellulare. Il complesso dei vacuoli
costituisce il vacuoma.
Le ipotesi sulla genesi del vacuolo sono diverse: dalla semplice idratazione di zone del citoplasma
alladegenerazione di organuli, ad invaginazioni del plasmalemma. Le ipotesi più accreditate
attualmente sono due e consistono (1) nell’ingrandimento e successivo distacco di vescicole del
reticolo endoplasmatico o dell’apparato di Golgi, (2) nella formazione ed evoluzione di strutture
particolari, chiamate GERL (Golgi- Endoplasmic Reticulum Layer), derivanti dalla fusione di RE e
apparato di Golgi. Dai GERL deriverebbero provacuoli che ingabbiandosi, avvolgerebbero porzioni
di citoplasma; successivamente si formerebbe un vacuolo lisosomiale nel quale la membrana interna
e il citoplasma ingabbiato sarebbero digeriti; infine i piccoli vacuoli derivati si fonderebbero fra loro.
Nei semi esiste una relazione tra vacuolo e corpi proteici; pare che la smobilitazione delle riserve
proteiche porti all’utilizzo delle membrane dei corpi proteici per la formazione del vacuolo, mentre,
al contrario, durante la formazione del seme, il vacuolo si suddivide in numerosi corpi proteici.
Le funzioni del vacuolo sono molteplici:
ridurre il volume metabolicamente attivo rendendo più favorevole alla vita della cellula il rapporto
superficie/volume.
• risolvere il problema della estrusione di metaboliti
• segregare prodotti secondari o di rifiuto
• risolvere problemi meccanici, contribuendo al turgore cellulare
• accantonare sostanze di riserva e nutrienti
Il contributo del vacuolo al turgore cellulare deriva principalmente dalle proprietà osmotiche delle
sostanze contenute nel succo vacuolare che richiamando acqua favoriscono un aumento della
pressione rivolta verso l’esterno, bilanciata dalla resistenza opposta dalla parete. Nel caso in cui la
soluzione esterna alla cellula abbia concentrazioni di soluti maggiori di quelle del vacuolo
(ambiente ipertonico), l’acqua, anziché entrare, esce. Ciò provoca la diminuzione della pressione di
turgore esercitata verso l’esterno, il collassamento del vacuolo e il distacco del citoplasma dalla
parete: questo fenomeno si chiama plasmolisi ed entro certi limiti può essere reversibile
(appassimento), ma oltre una soglia è irreversibile (avvizzimento) e porta alla morte della cellula.
La variazione delle condizioni di turgore, determinate dal tenore idrico del vacuolo, è importante
anche in diversi movimenti delle foglie e nella regolazione degli stomi.
La presenza di enzimi idrolitici nel vacuolo permette la decomposizione di macromolecole
garantendo il turnover continuo dei componenti cellulari.
L’accumulo dei prodotti di riserva nel vacuolo è generalmente reversibile e spesso si alternano fasi
di accumulo e fasi di utilizzo.
15
16
Il tonoplasto presenta la struttura lipoproteica tipica delle membrane, con una ATPasi che pompa
protoni nella fase acquosa del vacuolo e determina l’abbassamento del pH del succo vacuolare
intorno a 4-5.
Il succo vacuolare varia nella sua composizione in modo considerevole, in rapporto alla specie, al
tipo di cellula e della sua localizzazione ed in base allo stadio di sviluppo e al momento rispetto al
ritmo circadiano della pianta.
Le sostanze in esso contenute possono essere suddivise in tre categorie:
a) inclusi idrofili
• acidi organici (es.:, acido citrico, acido malico; che conferiscono il sapore acido)
• sali minerali (per lo più dissociati in ioni; es.: cloruri, solfati, fosfati, nitrati di Na, K, Ca, Mg)
• glucidi (es.: fruttosio in diversi frutti, glucosio nell’uva, saccarosio nella barbabietola e nella canna
da zucchero, ecc.)
• aminoacidi (soprattutto asparagina e glutammina)
• alcaloidi (con funzione di difesa contro erbirvori o parassiti, rifiuto o riserva di azoto; es.: caffeina,
nicotina, atropina, papaverina, ecc.)
• glucosidi (es.: digitossina)
• tannini (derivati dall’acido gallico o della pirocatechina, con funzione antiputrefattiva, di difesa
contro attacchi da microrganismi)
• pigmenti antocianici e flavonici (che conferiscono colore ai fiori e alle foglie)
b) inclusi idrofobi
• lipidi (trigliceridi soprattutto, come in molti semi e frutti)
• oli essenziali (derivati isoprenici, molto volatili, con funzioni diverse)
• resine, balsami e latici (soluzioni oleose varie di prodotti terpenici, soprattutto con funzione
antiputrefattiva e di difesa)
c) inclusi insolubili
• ossalato di calcio (in forma cristallina con cristalli isodiametrici –druse- o aghiformi –rafidi-,
probabilmente rappresentano un mezzo per precipitare l’acido ossalico, inibitore di diversi
complessi enzimatici)
• silice (per esempio in alcune graminacee, in aggiunta alla silice che mineralizza la parete)
• gesso (solfato di calcio idrato, per esempio in alcune alghe)
• aleurone (corpi con matrice proteica e una massa cristallina –cristalloide- ed una o più particelle
globose –globoidi- contenenti fitina, cioè derivato dell’acido fitico salificato contemporaneamente
con fosforo, calcio e magnesio)
16
17
I cloroplasti sono specializzati per la fotosintesi; forma, dimensioni e numero sono molto
variabili a seconda del gruppo sistematico. Le alghe verdi ne hanno pochi o addirittura uno solo per
cellula, il tessuto a palizzata delle piante superiori anche un centinaio. Nei vegetali inferiori, quando
il cloroplasto è unico, esso è particolarmente grande (100 μm); nelle piante superiori la media è 2-3
x 4-6 μm. Forma e dimensioni sono pressoché costanti ed ereditarie; nelle cellule poliploidi i
cloroplasti hanno dimensioni maggiori. Il numero dei cloroplasti in una pianta è notevole; per
esempio è stato calcolato che in una foglia di ricino sono presenti circa 400.000 cloroplasti per mm 3.
La forma dei cloroplasti nelle alghe può variare da quella a stella, a coppa, a spirale, a nastro, a
reticolo, ecc.; nelle piante superiori è a disco o lenticolare. La localizzazione all’interno della cellula
non è casuale: sono per lo più addossati alla parete nei punti dove vi sono spazi intercellulari che
facilitano scambi gassosi; correnti citoplasmatiche movimentano i cloroplasti che possono essere
orientati rispetto alla fonte di luce, grazie al citoscheletro.
La struttura dei cloroplasti è complessa: la duplice membrana, che forma uno spazio intermembrana
di circa 3 nm, circonda lo stroma particolarmente ricco d’enzimi, nel quale si evidenzia un sistema
lamellare costituito da membrane, chiamate tilacoidi. La membrana esterna ha una componente
lipidica maggiore ed è relativamente permeabile, mentre quella interna svolge un particolare ruolo
selettivo e presenta proteine di trasporto; le due membrane sono tra le più ricche di lipidi in assoluto
ed in particolare abbondano di glicolipidi. Questo involucro esterno dei cloroplasti presenta anche
enzimi, come la nitrato reduttasi che partecipazione al metabolismo dell’azoto.
Lo stroma ha aspetto granulare, con granuli di amido primario, plastoglobuli (goccioline lipidiche),
enzimi deputati allo svolgimento della fase oscura della fotosintesi (ma anche per la sintesi di acidi
grassi, acidi nucleici, riduzione del solfato, ecc.), DNA, RNA, ribosomi 70S.
Il sistema lamellare è probabilmente derivato dalla membrana interna per invaginazioni che hanno
dato origine a sacculi o cisterne chiuse appiattite, i tilacoidi, in parte impilate le une sulle altre; le
pile di cisterne o sacculi sono definite grana (ogni pila è un granum), sono formate da tilacoidi
granari e collegate fra loro da tilacoidi intergrana o stromatici. Ogni granum è costituito da 2 a 200
tilacoidi impilati ed ha un diametro di 0,3-1 μm, ma si tratta di un sistema dinamico con variazioni
continue di formazione e disaggregazione dei grana. Esistono alcune eccezioni (certe piante
tropicali C4) nelle quali mancano i grana. Le membrane tilacoidali amplificano enormemente la
superficie di scambio e delimitano uno spazio interno, il lumen tilacoidale o spazio intratilacoidale.
Esse partecipano alla fase luminosa e sono fortemente specializzate, con il 50% di lipidi (per la
maggior parte glicolipidi insaturi) e 50% di proteine. Le membrane tilacoidali sono configurate in
un modo peculiare: le porzioni intergrana presentano ATPasi che esporta H+ dal lumen (con pH5)
allo stroma (con pH 8), pigmenti accessori, il complesso proteico del fotosistema I con clorofilla a e
la catena di trasporto del citocromo b6f, mentre sulle porzioni granari sono localizzati pigmenti
accessori e il complesso proteico del fotosistema II con clorofilla a. La disposizione delle molecole
proteiche e dei pigmenti sulle membrane è visibili con tecniche di criodecappaggio. I pigmenti sono
rappresentati prevalentemente da clorofilla a (75%), clorofilla b (25%), carotenoidi e derivati
(xantofille, luteina); nelle alghe rosse i rodoplasti (corrispondenti dei cloroplasti) hanno clorofilla a,
clorofilla d, carotenoidi e ficoeritrina; nelle alghe brune i fenoplasti contengono clorofilla a,
clorofilla c e fucoxantina.
17
18
cloroplasto visto al microscopio
.
modello tridimensionale di
cloroplasto
All’interno dello stroma si trovano le membrane tilacoidali o tilacoidi, dove avvengono le prime
fasi della fotosintesi. Le membrane tilacoidali si distinguono in membrane granali, impacchettate
tra loro, e membrane stromatiche, in contatto con lo stroma. Lo spazio interno ai tilacoidi si
chiama lumen.
Struttura ultramicroscopica:
Sulle membrane dei tilacoidi sono ancorati le clorofille . Qui si svolge la fase luminosa della
fotosintesi, che corrisponde alla cattura e conversione dell'energia luminosa in energia chimica.
Nello stroma, invece, avviene la fase oscura, che coincide con la vera e propria organicazione della
CO2.
18
19
La fotosintesi clorofilliana è l’insieme delle reazioni durante le quali le piante verdi producono
sostanze organiche a partire da CO2 e dall’acqua, in presenza di luce
Mediante la clorofilla, l'energia solare (luce) viene trasformata in uno zucchero definito glucosio
fondamentale per la vita della pianta la cui formula chimica è:C6H12O6,ovvero 6 atomi di
carbonio,12 di idrogeno e 6 di ossigeno. Inoltre alla pianta (detta autotrofa) rimangono 6 atomi di
ossigeno atmosferico di cui si libera grazie agli stomi delle sue foglie
La fotosintesi clorofilliana avviene per tappe riunibili in due fasi: la fase luminosa (o fase lucedipendente), dipendente dalla luce; la fase di fissazione del carbonio (o fase oscura, indipendente
dalla luce) di cui fa parte il ciclo di Calvin.
Oggi questo processo è quello nettamente dominante sulla Terra, per la produzione di composti
organici da sostanze inorganiche . Inoltre, la fotosintesi è l'unico processo biologicamente
importante in grado di raccogliere l'energia solare, da cui, fondamentalmente, dipende la vita sulla
Terra.
. Il bilancio complessivo dei flussi di ossigeno e Co2 da e verso l'ambiente esterno è comunque a
favore della fotosintesi ovvero la pianta si comporta come un 'pozzo' (assorbitore) di
accumulazione di carbonio piuttosto che come una 'sorgente' (emettitore) verso l'ambiente esterno
di carbonio e viceversa una 'sorgente' di ossigeno piuttosto che un 'pozzo' di ossigeno
Questo perché parte del carbonio assorbito e non utilizzato dal ciclo ossidativo della pianta rimane
fissato sotto forma di cellulosa e lignina nelle pareti cellulari delle cellule 'morte' che costituiscono
il legno interno della pianta.
19