IDONEITA’ ALLA CLASSE 5
SCIENZA DELLA NATURA
INDICE
O.G.M.
ATMOSFERA TERRESTRE
OZONOSERA
LA SISMOGRAFIA
I MOVIMENTI DELLA TERRA
FONTI ENERGETICHE ALTERNATIVE
LA TERRA
IL BIG BANG
LA TEORIA DELL’EVOLUZIONE
LA CELLULA ANIMALE E VEGETALE
LA FOTOSINTESI CLOROFILLIANA
LA DIVISIONE CELLULARE
MITOSI E MEIOSI
IL METABOLISMO CELLULARE
LA TRASMISSIONE DEI CARATTERI EREDITARI
TESSUTI
LE TRE LEGGI DI KEPLERO
GLI OGM
Gli
organismi geneticamente
modificati
(OGM),
sono
organismi
caratterizzati da un patrimonio genetico (genoma) alterato rispetto a
quello tipico della propria specie, per l’introduzione artificiale di uno o
più geni provenienti da altri organismi
Per ottenere organismi
transgenici
si
utilizzano
le
tecniche
dell’ingegneria genetica. Il frammento di DNA in cui si trova il gene da
inserire viene iniettato in una cellula batterica, o in una cellula uovo (che
verrà successivamente fecondata) o in un embrione. Per potere essere
attivo, il frammento di DNA deve essere associato a un vettore
d’espressione, ossia a un’altra porzione di DNA specifica che controlla le
modalità di espressione del gene da trasferire; ad esempio, esso permette
che il gene si esprima (cioè svolga la propria attività) soltanto in
determinati tessuti.
Il DNA estraneo viene inoculato per microiniezione nella cellula
ricevente; dopo l’inoculazione, il nuovo gene si integra con il DNA di
questa, e può di conseguenza venire trasmesso a tutte le cellule che
derivano per successive mitosi dalla cellula ricevente.
Nel caso si utilizzino embrioni, i frammenti di DNA contenenti i geni
possono essere anche inseriti tramite un virus-vettore, ossia tramite un
virus infettivo nel quale, a sua volta, è stato inoculato il frammento di
DNA. Si calcola che la percentuale di successo di questa tecnica, che si
traduce con il numero di organismi transgenici vitali e nei quali i geni
estranei sono funzionanti, sia dell’1%.
Il controllo dell’avvenuta integrazione del gene nel patrimonio genetico
dell’organismo ricevente può essere fatto prelevando da alcune cellule
transgeniche campioni di DNA ed esaminandoli, in genere mediante la
tecnica nota come reazione a catena della polimerasi (PCR).
L’IMPIEGO DEGLI ORGANISMI TRANSGENICI
Nella ricerca biologica e genetica, l’impiego di organismi transgenici è
rilevante nell’ambito degli studi sulla funzione di geni specifici; infatti,
l’immissione di un gene estraneo in un organismo determina l’insorgenza
in questo di particolari caratteristiche (come la resistenza a un erbicida o
la capacità di sintetizzare una data proteina) che, confrontate con quelle
degli individui della stessa specie, permettono la comprensione del ruolo
di quel gene.
A scopo di ricerca, sono impiegati anche particolari tipi di organismi
transgenici, i cosiddetti knock-out, in cui un gene dell’organismo viene
eliminato o inattivato; alcuni topi così modificati, ad esempio, sono stati
utilizzati per studiare il ruolo funzionale di alcuni geni specifici nello
sviluppo embrionale. Disattivando in animali da laboratorio il gene
corrispondente a un gene non funzionale nei pazienti affetti da una
particolare malattia, si possono creare modelli utili a fini diagnostici e
terapeutici.
UNA QUESTIONE CONTROVERSA
L’impiego di organismi geneticamente modificati è uno dei più dibattuti
temi della bioetica. Infatti, già da tempo la creazione di nuove culture
vegetali o di microrganismi modificati può essere siglata da brevetto; la
possibilità di estendere questa pratica anche a organismi più complessi, e
ai procedimenti industriali che ne permettono l’ottenimento, suscita
attualmente atteggiamenti diversi: da un lato entusiasmo, per le nuove
prospettive economiche e scientifiche che potrebbero derivarne;
dall’altro, preoccupazione, per tutte le implicazioni, soprattutto etiche e
sociali.
Si ritiene che la questione dell’impiego delle specie transgeniche non
debba limitarsi a un’analisi dei costi e dei benefici economici, e che le
attuali leggi sui brevetti, relative a strumentazioni, non possano essere
semplicemente estese a organismi viventi. Sono inoltre oggetto di
discussione le possibili conseguenze sulla biodiversità e sugli equilibri
degli ecosistemi dell’immissione nell’ambiente di organismi modificati,
con caratteri che potrebbero venire trasmessi alla discendenza; inoltre,
suscitano perplessità i possibili effetti a lungo termine sulla salute umana
del consumo di prodotti derivanti da organismi geneticamente modificati
L'ATMOSFERA TERRESTE
Tra tutti i pianeti del Sistema Solare la Terra è l'unico a possedere
un'atmosfera ricca di ossigeno e di azoto, elementi fondamentali per
consentire la presenza della vita in tutte le sue forme, animali e vegetali
(almeno come la concepiamo noi). L'atmosfera svolge anche un ruolo
essenziale per garantire la protezione della vita: essa costituisce infatti
uno schermo estremamente efficiente per assorbire le radiazioni
ultraviolette e per il flusso di particelle provenienti dal Sole, che
altrimenti la distruggerebbero quasi immediatamente.
L'atmosfera protegge inoltre la superficie terrestre dall'impatto delle
meteoriti che, a eccezione di alcune di dimensioni particolarmente
rilevanti, si disintegrano per l'attrito con gli strati superiori.
L'atmosfera svolge anche un ruolo molto rilevante nella definizione
della morfologia della superficie terrestre: i moti, i fenomeni
meteorologici, le reazioni chimiche che hanno luogo nella bassa
atmosfera costituiscono una delle cause più importanti delle continue
trasformazioni della litosfera e della idrosfera. L'atmosfera interviene
infatti in modo quasi esclusivo nei processi di erosione e di
sedimentazione oltreché nel ciclo dell'acqua.
Composizione dell'atmosfera
Lo strato di atmosfera che circonda la Terra è estremamente sottile.
Se si considera solo la parte più densa che giunge sino a circa 60 km al di
sopra della superficie terrestre, si ha uno spessore che è pari a solo un
centesimo del raggio terrestre all'equatore, che è di 6.378 km.
In realtà, non si può definire un vero e proprio limite superiore
dell'atmosfera, ma solo una regione di transizione in cui essa si confonde
con lo spazio interplanetario.
La composizione chimica e le caratteristiche fisiche dell'atmosfera
variano secondo la quota. La composizione dell'atmosfera tra il suolo e
10-12 km può però considerarsi pressoché costante e formata da un
miscuglio di gas tra i quali dominano nettamente l'azoto e l' ossigeno.
Pressione atmosferica
Peso esercitato sull'unità di superficie dalla colonna d'aria al di sopra
del punto che si considera.
La pressione atmosferica viene generalmente misurata in millimetri di
mercurio o millibar. Un millibar (mb) è un millesimo di bar. Un bar è la
pressione esercitata da 750,06 mm di mercurio (mmHg) alla temperatura
di 0°C (760 mm di mercurio corrispondono a 1.013 millibar). La
pressione atmosferica è un importante fattore che influenza l'andamento
del tempo meteorologico e che svolge pure un ruolo nella sua previsione.
Umidità atmosferica
Ai costituenti dell'atmosfera va aggiunta l'acqua che, sotto forma di
vapore e di nubi, ne rappresenta circa lo 0,33% della massa totale e
determina la cosiddetta umidità atmosferica.
La presenza dell'acqua è l'elemento determinante ai fini della
considerazione dei fenomeni meteorologici.
Anche l'umidità atmosferica varia con la quota. Al livello del suolo e in
prossimità di esso la quantità di vapore acqueo dipende dalle condizioni
climatiche e dalla posizione geografica e può raggiungere una percentuale
in volume pari al 4%. Nei primi 8 km di atmosfera l'umidità varia, nelle
zone temperate, da 6,8 a 0,1 g/m³.
La diminuzione continua sino a circa 15 km, dove si ha uno strato molto
secco. Da 15 a 30 km si ha un certo aumento con la presenza di un tipo
particolare di nubi, le nubi madreperlacee, che si formano tra i 25 e i 30
km.
Una certa quantità di umidità è presente anche a un'altezza di circa 80
km, dove si osservano talvolta le cosiddette "nubi nottilucenti".
Elettricità atmosferica
L'atmosfera è sede di un campo elettrico, cioé è una zona di spazio
dove si risente l'azione di forze elettriche. Tale campo è prodotto dalle
cariche elettriche negative accumulate sulla Terra e dalle cariche,
costituite soprattutto da ioni positivi, presenti nell'aria.
La struttura del campo elettrico atmosferico è soggetta a notevoli e
brusche variazioni per effetto delle pereturbazioni atmosferiche.
La capacità dell'aria di poter condurre la corrente elettrica varia poi
notevolmente con l'altezza. Tale conducibilità è bassa in prossimità del
suolo, ma aumenta rapidamente con l'altezza per effetto della
ionizzazione dell'aria prodotta dalle radiazioni cosmiche.
Negli strati superiori è addirittura presente una zona in cui i gas sono
ionizzati e che quindi ha un'altissima conducibilità elettrica.
Temperatura atmosferica
Il calore dell'atmosfera è dovuto all'irraggiamento solare. Il 30% della
radiazione solare viene riflesso e diffuso nello spazio esterno dalle nubi;
il restante viene assorbito dall'aria e dalla superficie terrestre. L'atmosfera
ha anche un'azione selettiva sulle radiazioni solari.
L'assorbimento dei raggi ultravioletti avviene nell'alta atmosfera e vi
provoca un tipico riscaldamento. La radiazione infrarossa fortemente
calorifica, viene invece assorbita nella zona più bassa dell'atmosfera
dall'anidride carbonica e dal vapore acqueo presenti vicino alla superficie
terrestre.
Queste due sostanze si comportano come il tetto di vetro di una serra e
favorisce l'immagazzinamento di calore in prossimità del suolo.
Questo effetto, noto come "effetto serra", contribuisce notevolmente a
determinare la temperatura diurna sulla superficie terrestre, che risulta in
media di circa 20 °C, mentre senza di esso dovrebbe essere di circa -23
°C.
La temperatura dell'aria è variabile fortemente con l'altezza; fino a una
quota di 10-12 km diminuisce regolarmente di 0,5-0,7 °C ogni 100 m
sino a raggiungere -55 °C circa; da 12 km a 50 km aumenta sino a
raggiungere 0 °C circa. Da questo punto la temperatura riprende
nuovamente a diminuire sino a giungere a valori tra -70 e -100 °C a 80-90
km di quota.
Ha allora inizio un rapido aumento che raggiunge negli strati atmosferici
più alti i 1.000-2.000 °C circa. Data però l'estrema rarefazione delle
molecole a queste altezze, il termine temperatura non ha riferimento
alcuno con sensazioni fisiologiche, me è correlato unicamente all'energia
di agitazione termica delle singole molecole. Il modo in cui varia la
temperatura con l'altezza viene usato per definire i diversi strati
dell'atmosfera stessa.
OZONOSFERA
Ozonosfera Strato dell'atmosfera terrestre compreso tra i 20 e i 50 km di
quota, caratterizzato da una concentrazione di ozono relativamente alta,
che può raggiungere le 10 ppm (parti per milione). A queste quote,
l'ozono si forma naturalmente per effetto dell'interazione delle molecole
di ossigeno presenti nell'atmosfera con le radiazioni ultraviolette
provenienti dal Sole. La concentrazione naturale di ozono rimane
pressoché costante grazie all’equilibrio tra il processo di produzione e
quello di distruzione operato da alcuni composti dell'azoto, anch’essi
presenti in atmosfera. L’assorbimento della radiazione ultravioletta
nell’ozonosfera produce un’inversione nell’andamento della temperatura
in funzione della quota: mentre nella troposfera la temperatura diminuisce
al crescere della quota, nell’ambito dell’ozonosfera essa aumenta al
crescere della distanza dalla superficie terrestre.
Importanza dell’ozonosfera per gli esseri viventi
A livello della stratosfera, lo strato di ozono crea una sorta di schermo
protettivo che assorbe le dannose radiazioni ultraviolette provenienti dal
Sole, consentendo la vita sulla Terra. Questo tipo di radiazioni, infatti,
altamente energetiche e penetranti, possono alterare e danneggiare il
DNA degli esseri viventi. Nella troposfera, invece, la formazione
dell’ozono è correlata soprattutto alle emissioni inquinanti degli
autoveicoli e delle industrie, che rilasciano nell’aria anidride solforosa
(SO2), ossidi di azoto (NOx) e composti organici volatili. Questo ozono
ha un notevole potere ossidante e risulta nocivo per la salute degli
organismi, uomo compreso.
Il “buco nell’ozono”
Negli anni Settanta del Novecento alcuni ricercatori rilevarono che il
naturale assottigliamento stagionale dello strato di ozono, che
periodicamente si osserva al di sopra del continente antartico nei mesi di
settembre e ottobre (la cosiddetta “primavera australe”), stava assumendo
dimensioni
allarmanti.
Il
fenomeno,
chiamato
deplezione
ma
comunemente noto come “buco nell'ozono”, avviene naturalmente e ha
un’origine ancora non chiara, e può perdurare anche per parecchi mesi;
tuttavia, nel 1985, i rilevamenti compiuti dalla stazione scientifica inglese
Antarctica Survey evidenziarono una diminuzione del 65% della
concentrazione dell'ozono, localizzata per il 95% negli strati atmosferici
compresi tra 13 e 22 km di distanza dalla superficie del pianeta.
L'estensione e la durata di questa variazione stanno assumendo
dimensioni sempre più ampie, come hanno confermato i rilevamenti
eseguiti con palloni aerostatici e satelliti meteorologici; la concentrazione
complessiva dell'ozono nell'ozonosfera è in costante diminuzione e non
solo al di sopra del continente antartico, ma anche in corrispondenza delle
regioni artiche.
Quali responsabili dell’alterazione della molecola dell’ozono sono stati
ritenuti i clorofluorocarburi o CFC (ampiamente impiegati come
propellenti nelle bombolette spray, come fluidi refrigeranti nei frigoriferi
e come agenti schiumogeni) e un gruppo di altre sostanze chiamate
genericamente ODS (Ozone-Depleting Substances). Tra queste vi sono:
gli HCFC (idroclorofluorocarburi); i cosiddetti halons (composti
estinguenti come il bromoclorodifluorometano, bromotrifluorometano,
dibromotetrafluoroetano); il metilbromuro; il tetracloruro di carbonio; il
metilcloroformio. Queste molecole complesse sono in grado di
raggiungere l'ozonosfera e di decomporre le molecole di ozono. Sotto
l'azione dei raggi ultravioletti, infatti, le molecole dei CFC si
decompongono in atomi di cloro e in altri derivati clorurati, che, a loro
volta, reagiscono con l'ozono e lo convertono in ossigeno biatomico,
liberando monossido di cloro che va a degradare altre molecole di ozono.
Gli ODS sono molto stabili nella troposfera e si degradano solo per
effetto degli intensi UV della stratosfera.
La necessità di affrontare il fenomeno considerandone tutti gli aspetti e le
ripercussioni su scala globale ha spinto i rappresentanti delle comunità
scientifica, politica ed economica a confrontarsi in periodici incontri, che
hanno l’obiettivo di stabilire strategie comuni di intervento. Il primo
incontro riguardante il buco nell’ozono fu la Conferenza di Vienna,
tenutasi nel 1985. Nel 1987 il Protocollo di Montréal segnò la messa al
bando dei CFC; la persistenza di questi composti in atmosfera, d’altra
parte, fa sì che eventuali effetti positivi dei provvedimenti in favore
dell’atmosfera si manifestino dopo numerosi anni (Vedi Clima).
Per monitorare costantemente il fenomeno del buco nell’ozono, nel 1991
la NASA lanciò in orbita un satellite artificiale di 7 tonnellate di peso
(l'Upper Atmosphere Research Satellite, satellite per la ricerca sull'alta
atmosfera). Da una quota di 600 km, il satellite continua a inviare a terra
dati sulle variazioni della concentrazione di ozono ad altitudini differenti,
oltre ad altri dati che hanno consentito di tracciare una mappa completa
della composizione chimica degli strati più alti dell'atmosfera. La
riduzione dello strato di ozono viene osservata ogni anno nel periodo
compreso tra settembre e ottobre. Un preoccupante fenomeno è stato
registrato nel settembre 2002: il buco antartico si è suddiviso in due parti,
assumendo una forma “a otto”, ciascuna delle quali si è estesa
allontanandosi dalla zona occupata originariamente. L’anomalia è stata
osservata per la prima volta da quando è iniziato il monitoraggio della
deplezione dell’ozono, e sembra causata dalle intense perturbazioni
dell’atmosfera verificatesi nei mesi precedenti. Solo qualche mese prima,
le rilevazioni del CSIRO australiano (Commonwealth Scientific and
Industrial Research Organisation) avevano indicato che il buco si era
ristretto, probabilmente a seguito di un periodo prolungato di temperature
atmosferiche insolitamente elevate.
SISMOGRAFIA
Il fenomeno dei terremoti è strettamente legato al processo globale che
è responsabile degli impercettibili mutamenti che ininterrottamente
modificano la struttura interna ed esterna della terra. Questa continua
trasformazione si realizza attraverso il movimento relativo di ampie
zone della superficie terrestre (placche) che scivolano l’una sull’altra
lungo le “faglie”, grandi e profonde spaccature della superficie
terrestre, sottoponendo in questo modo gli strati interni delle rocce a
sforzi enormi.
Quando tali sforzi raggiungono la limite della resistenza offerta dalle
rocce, si produce un’improvvisa frattura che libera energia elastica
accumulata nella fase di compressione. Parte dell’energia rilasciata
viene trasformata in onde di pressione, dette onde sismiche che, a
partire dal punto di frattura interno alla terra detto “ipocentro”, si
propagano lungo tutte le possibili direzioni.
I terremoti sono provocati dalle onde sismiche che, giungendo sulla
superficie terrestre, determinano scosse e bruschi movimenti del suolo.
L’area maggiormente coinvolta dall’evento sismico è detta “epicentro
del terremoto” e corrisponde al punto sulla superficie più vicino
all’ipocentro.
Esistono tre tipi di onde sismiche: le onde P o primarie, che giungono
per prime avendo velocità maggiore, le onde S o secondarie, che
giungono per seconde e le onde di superficie che sono generate dalle
onde P e S quando queste incontrano la superficie terrestre.
I sismologi impiegano opportuni strumenti, detti sismografi, per
misurare il momento di arrivo e l’intensità delle varie componenti (fasi)
delle onde sismiche. Queste misure vengono impiegate per determinare
le caratteristiche fondamentali di un terremoto quali ad esempio la
posizione dell’ipocentro, dell’epicentro e la sua intensità.
La registrazione dello onde sismiche per mezzo di sismografi è detta
sismogramma. In figura si mostra un esempio molto semplificato di
sismogramma nel quale è possibile individuare l’arrivo dei fronti
d’onda delle componenti P ed S.
E’ importante osservare che, poiché le onde P viaggiano più
velocemente delle onde S, l’intervallo temporale tra l’arrivo dei due
treni d’onda diventa sempre più grande man mano che aumenta la
distanza tra l’epicentro del terremoto e il punto di misurazione. In altri
termini conoscendo l’intervallo temporale e la velocità di propagazione
delle due onde è possibile determinare la distanza trala stazione di
osservazione e l’epicentro del terremoto (distanza epicentrale) .
Tuttavia questo metodo non permette di stabilire la direzione di
provenienza dell'onda per cui l’epicentro del terremoto può trovarsi in
un punto qualsiasi sulla circonferenza centrata attorno alla stazione di
osservazione e con raggio pari alla distanza epicentrale.
Per determinare le coordinate dell’epicentro di un terremoto è
necessario conosce almeno tre distanze epicentrali calcolate in
altrettante stazioni sismografiche. La posizione dell’epicentro viene
quindi ottenuta con il metodo della “triangolazione” e coincide con il
punto in cui, con buona approssimazione, si incrociano le circonferenze
centrate attorno alle tre diverse stazioni di osservazione.
I MOVIMENTI DELLA TERRA
IL MOVIMENTO DI ROTAZIONE
La Terra ruota su se stessa intorno ad un asse immaginario detto ASSE DI
ROTAZIONE che passa per i poli; il periodo di rotazione è detto GIORNO
SIDEREO; è un moto periodico che avviene da ovest verso est, osservato dal
polo nord celeste ed in senso antiorario.
La VELOCITA’ ANGOLARE di rotazione è uguale in ogni luogo della
Terra (360° in un giorno). Ciascun punto della superficie percorre in un
giorno un circonferenza più o meno lunga a seconda della latitudine e
dell’altitudine; quindi varia la VELOCITA’ LINEARE DI ROTAZIONE;
essa aumenta man mano che ci si allontana dall’asse di rotazione ed aumenta
con l’altitudine. Il periodo di rotazione attualmente ha una durata di 23h 56
min 4sec, ma sta rallentando progressivamente di millesimi di secondo ogni
secolo.
LE CONSEGUENZE DELLA ROTAZIONE TERRESTRE:
• L’ALTERNARSI DEL GIORNO E DELLA NOTTE: la terra ha
forma pressoché sferica, così in ogni istante solo metà della sua
superficie riceve luce e calore dal sole; la linea che separa le due zone
è detta CIRCOLO DI ILLUMINAZIONE. Ogni punto della terra
passa continuamente da una zona all’altra e vede alternarsi ogni
giorno il dì e la notte. Per la presenza dell’atmosfera che provoca
fenomeni di diffusione il passaggio è graduale. Per questo il circolo di
illuminazione è una fascia in cui il dì e la notte sono divisi da un
debole chiarore detto CREPUSCOLO. A causa della rifrazione il sole
è visibile un po’ prima dell’alba ed un po’ dopo il tramonto. La durata
del crepuscolo aumenta con la latitudine. Il piano del circolo di
illuminazione è in ogni istante perpendicolare ai raggi solari.
• L’APPARENTE MOVIMENTO GIORNALIERO DELLA TERRA:
ogni giorno la sfera celeste sembra ruotare da est verso ovest intorno
al prolungamento dell’asse terrestre. Questo movimento è apparente
ed è provocato dal fatto che la Terra ruota intorno al suo asse nel
verso opposto.
• LA FORZA CENTRIFUGA: è una forza fittizia ovvero apparente,
del cui movimento risentono solo i corpi solidali con il sistema di
rotazione. Agisce in direzione perpendicolare all’asse di rotazione ed è
diretta verso l’esterno. Il suo valore aumenta con l’altitudine e con
l’allontanarsi dall’asse. A livello del mare la forza centrifuga ha
intensità diverse a seconda della latitudine. E’ nulla ai poli e massima
all’equatore.
Essa provoca due importanti effetti:
1. ha contribuito a formare lo schiacciamento polare ed il
rigonfiamento equatoriale;
2. contrasta parzialmente la forza gravitazionale, riducendo il
valore dell’accelerazione di gravità.
Essa si oppone alla forza di gravità perfettamente solo all’equatore
essendo diretta dall’asse e non dal centro verso l’esterno.
• LA FORZA DI CORIOLIS: è una forza fittizia che
interessa i corpi che non sono vincolati alla superficie
terrestre; ogni corpo che si muove dall’equatore ai poli
viene deviato verso destra nell’emisfero settentrionale e
verso sinistra nell’emisfero meridionale. La forza si fa
sentire anche lungo i paralleli, ma è nulla all’equatore; il
suo valore dipende dalla latitudine e dalla velocità del
corpo. Ha una notevole influenza sulla direzione della
circolazione atmosferica e dei grandi circuiti delle
correnti oceaniche.
IL MOVIMENTO DI RIVOLUZIONE
La terra descrive un’orbita ellittica intorno al sole, che ne occupa uno dei
due fuochi; il piano dell’orbita della Terra intorno al sole è detto PINO
DELL’ECLITTICA.
ANNO SIDEREO: periodo di 365 d 6 h 9 min 9,5 s che la terra impiega per
la rivoluzione.
AFELIO: punto in cui la Terra si trova più distante dal Sole.
PERIELIO: punto il cui la terra è più vicina al Sole.
La linea che idealmente li congiunge è la LINEA DEGLI APSIDI. La
VELOCITA’ DI RIVOLUZIONE non è costante, è massima al perielio e
minima all’afelio.
L’asse di rotazione terrestre è inclinato rispetto al piano dell’eclittica di 66°
33’ e durante il moto di rivoluzione si può considerare sempre parallelo a se
stesso. Conseguenza è che il piano dell’equatore è inclinato rispetto al piano
dell’eclittica di 23°27’. Durante l’anno cambia la direzione della Terra
rispetto alla direzione dei raggi solari, quindi si modifica la posizione del
circolo di illuminazione rispetto all’asse terrestre.
EQUINOZIO DI PRIMAVERA: 21 marzo
EQUINOZIO D’AUTUNNO: 23 settembre
In questi giorni la durata del dì e della notte sono uguali. Il sole è allo
ZENIT(raggi perpendicolari al piano dell’osservatore) a mezzogiorno
sull’equatore. In questi giorni il circolo di illuminazione è tangente ai poli.
SOLSTIZIO D’ESTATE: 21 giugno; il sole è allo zenit a mezzogiorno sul
TROPICO DEL CANCRO.
SOLSTIZIO D’INVERNO: 22 dicembre; il sole è allo Zenit a mezzogiorno
sul TROPICO DEL CAPRICORNO.
In queste due posizioni il circolo di illuminazione è alla massima distanza
dai poli; si ha la massima differenza tra la durata del dì e della notte; il
circolo di illuminazione è tangente al circolo polare artico e al circolo polare
antartico.
LE CONSEGUENZE DEL MOTO DI RIVOLUZIONE:
• GIORNO SOLARE E GIORNO SIDEREO: Il GIORNO
SIDEREO è l’intervallo di tempo che intercorre tra due
passaggi successivi della medesima stella sul meridiano
del luogo; 23h 56m 4s.
Il GIORNO SOLARE è l’intervallo di tempo tra due culminazioni( punto
di massima altezza raggiunto da un corpo celeste sull’orizzonte)
consecutive del sole sullo stesso meridiano: 24h.
Quando la Terra ha compiuto un’intera rotazione, l’osservatore non si
trova più nella stessa posizione del giorno precedente rispetto al Sole, dal
momento che la terra si è mossa lungo la sua orbita di rivoluzione. Poiché
la terra impiega 4 min per ruotare di 1° il girono solare risulta più lungo
del giorno sidereo di 4 minuti.
Il giorno solare modifica la sua durata perché la velocità di rivoluzione
non è costante.
• IL MOVIMENTO APPARENTE DEL SOLE SULLO
SFONDO DELLO ZODIACO: Il Sole (movimento
apparente) ritarda ogni giorno di 4 minuti rispetto allo
sfondo delle stelle. Ogni notte troveremo in opposizione
al sole stelle diverse; se ogni giorno fissiamo la posizione
del sole a mezzogiorno sullo sfondo celeste, osserveremo
che in un anno descrive in cielo una linea chiusa che
giace su un piano (eclittica) inclinato di 23°27’ rispetto
all’0equatore.
Lo sfondo di stelle si chiama ZODIACO. Quando il sole si trova in una
costellazione, questa non è visibile.
Poiché l’eclittica è inclinata rispetto all’equatore, la declinazione del Sole,
cioè la sua distanza angolare dall’equatore celeste, cambia durante l’anno.
Il sole per sei mesi si trova nell’emisfero celeste boreale e per sei mesi in
quello astrale
FONTI ENERGETICHE ALTERNATIVE
In tutti i Paesi industrializzati vengono svolte ricerche per trovare fonti
energetiche alternative, che possano sostituire quelle dei combustibili
fossili usati fino ad oggi. Oggi, alcuni scienziati, però, hanno capito che si
possono utilizzare anche altre risorse, ad esempio:
-L’ ENERGIA EOLICA: i venti, che sono enormi spostamenti di masse
d’aria provocati dal riscaldamento solare, sono una fonte che l’ uomo ha
sfruttato anche in passato, nella navigazione a vela e nei mulini. Oggi si è
scoperto che l’energia eolica serve anche nelle centrali, infatti l’ energia
elettrica verrebbe prodotta da alternatori mossi dalla forza del vento. Ma
per essere conveniente, queste centrali richiedono un’ istallazione in
luoghi esposti al vento. In alcune zone in cui il vento è molto forte, si
stanno costruendo centrali di elevata potenza che ormai sono in fase di
sperimentazione o addirittura in esercizio. In Italia, invece, non essendoci
zone appropriate per la costruzione di queste centrali, l’ ENEL ha in
programma di costruire in Sardegna una centrale eolica costituita da dieci
aeromotori, come fonte alternativa per le utilizzazioni agricole, industriali
e civili.
-L’ ENERGIA DELLE MAREE: a causa dell’ attrazione del sole e della
luna, le acque degli oceani si sollevano formando onde di maree che si
ripetono due volte a giorno. Il sollevamento delle maree, racchiude una
grande quantità di energia che può essere anche sfruttata, ad esempio sulle
coste inglesi sono nati dei mulini che erano azionati dalla corrente delle
maree. In Francia, invece, è stata costruita una centrale mareomotrice. Il
funzionamento è molto semplice. Una diga chiude l’ estuario di un fiume,
quindi si forma un bacino che si riempie durante l’ alta marea. Durante la
bassa marea, invece, l’ acqua accumulata si riversa nel mare, quindi si
mettono in movimento le turbine ad elica alloggiate in apposite camere
alla base della diga. Molti paesi si stanno preoccupando di sfruttare l’
energia delle maree non solo come gli impianti francesi, ma anche
attraverso dispositivi galleggianti, atti a sfruttare il moto ondoso.
-L’ ENERGIA DELLE BIOMASSE: per biomasse si intendono tutti i rifiuti
organici, urbani, industriali e agricoli. Da queste sostanze si possono
ricavare nuovi combustibili, cioè attraverso alcuni procedimenti
termochimici e con alcuni procedimenti biochimici. Con i procedimenti
termochimici possiamo trasformare le biomasse attraverso la combustione
diretta dei rifiuti. Tra i processi biochimici, invece, il più importante è la
digestione anaerobica, che consiste nel far fermentare i rifiuti in assenza
d’ ossigeno, così le sostanze organiche si attaccano ai batteri anaerobici
trasformandole in biogas.
Questo particolare trattamento si rivela anche un metodo economico
anche in agricoltura, infatti sono stati sperimentati impianti in cui i rifiuti
organici delle stalle vengono convogliati in un apposito digestore, dove
subisce l’azione dei batteri anaerobici. I residui del processo di digestione
sono utilizzati come fertilizzante per i terreni e anche come alimento per
la piscicoltura.
-IMPIEGO DELL’ IDROGENO: l’ idrogeno è un gas che brucia
facilmente, sviluppando una notevole quantità di calore senza produrre
residui inquinanti. In natura, l’ idrogeno non si trova in nessun luogo e
quindi per essere utilizzato, deve essere ricavato decomponendo le
sostanze che lo contengono(ad esempio l’ acqua).
Però ci sono molti problemi per la produzione, il trasporto, l’
immagazzinamento e l’ utilizzazione. La produzione potrebbe avvenire
scomponendo l’ acqua del mare nei suoi componenti attraverso processi
elettrochimici. Così si otterrebbero grandi quantità di ossigeno e di
idrogeno.
L’ idrogeno deve poi essere trasportato tramite gasdotti interrati, nei
luoghi di consumo. Per quanto riguarda l’ impiego dell’ idrogeno, si
possono prendere due vie: o si utilizza come combustibile, oppure si
utilizza inserendolo in speciali pile che producono elettricità
dalla
combustione fredda tra idrogeno e ossigeno. Ma l’ idrogeno oggi risulta
un gas molto interessante, quindi si è cercato di impiegarlo anche nel
funzionamento delle macchine, infatti sono già state realizzate macchine
con motore ad idrogeno.
Per l’impiego automobilistico è indispensabile risolvere il problema dell’
immagazzinamento che deve essere effettuato necessariamente a bordo
del veicolo. Scartate le bombole perché sono un metodo molto pericoloso,
si è sperimentato che l’ idrogeno viene messo all’ interno di serbatoi
contenenti leghe metalliche spugnose. Per effetto dell’ elevata pressione,
la massa spugnosa assorbe il gas fino a saturarsi; conseguenza di una
reazione chimica reversibile che si svolge durante la fase di assorbimento
del gas. Quindi, scaldando la lega spugnosa, l’ idrogeno torna allo stato
gassoso e può essere impiegato per alimentare il motore di un veicolo.
FUSIONE NUCLEARE: nel corso degli ultimi anni, gli scienziati stanno
cercando di utilizzare le reazioni atomiche di fusione al fine di produrre
energia. Fino ad oggi, essa è stata impiegata solo nelle bombe all’
idrogeno, mentre nelle applicazioni a scopo pacifico sono rimaste a livello
sperimentale, perché ancora non si è trovato il metodo per utilizzare il
calore sviluppato dalla reazione che è identica a quella che si verifica all’
interno del Sole.
Attualmente, gli studi sono orientati sulla progettazione di impianti
chiamati di tipo “TOKAMAK” nei quali si cerca di confinare il plasma,
stabilizzandolo per mezzo di elevatissime correnti prodotte dal plasma
stesso. Nella fusione vengono risposte molte speranze, perché essa
potrebbe risolvere il problema energetico, utilizzando l’ idrogeno, che è
presente in grandi quantità di acqua.
LA TERRA
Terra Terzo pianeta del sistema solare in ordine di distanza dal Sole;
unico pianeta, allo stato attuale delle conoscenze, che ospiti la vita. Ha
una composizione prevalentemente rocciosa e una forma irregolare,
riconducibile in prima approssimazione a un ellissoide. Presenta una
struttura a strati, con un nucleo pesante, un mantello intermedio e una
crosta più leggera, ed è all’origine di una magnetosfera. Oltre che dal
Sole, attinge energia per le sue complesse dinamiche da una riserva di
calore immagazzinata al suo interno. Compie un complicato sistema di
moti periodici nello spazio, i più importanti dei quali sono la rotazione
intorno al proprio asse e la rivoluzione intorno al Sole.
Forma della Terra
Calcoli recenti basati sullo studio delle irregolarità orbitali di satelliti
artificiali hanno permesso di appurare che la
Terra presenta
effettivamente una forma di ellissoide, ma lievemente deformata “a
pera”: la differenza tra il raggio minimo equatoriale e il raggio polare
(distanza tra il centro della Terra e il Polo Nord) è di circa 21 km, inoltre
il Polo Nord “sporge” rispetto all’ellissoide regolare di circa 10 m,
mentre il Polo Sud è “schiacciato” di 31 m. Lo studio della forma della
Terra è oggetto di una disciplina che prende il nome di geodesia.
Moti della Terra
La posizione della Terra nello spazio non è stazionaria ma è il risultato di
una complessa composizione di moti con caratteristiche e periodicità
differenti. Insieme al suo satellite naturale, la Luna, il pianeta Terra orbita
intorno al Sole, a una distanza media di 149.503.000 km e con una
velocità media di 29,8 km/s, compiendo una rivoluzione completa in 365
giorni, 6 ore 9 minuti e 10 secondi (il periodo di rivoluzione è detto “anno
sidereo”). La traiettoria di quest’orbita è un’ellisse lievemente eccentrica,
ovvero pressoché circolare, con una lunghezza pari a circa 938.900.000
km. La Terra è inoltre in rotazione intorno al proprio asse; tale rotazione
avviene in senso inverso rispetto all’apparente moto del Sole e della sfera
celeste, vale a dire da occidente a oriente, e ha un periodo di 23 ore, 56
minuti e 4,1 secondi (giorno sidereo).
La Terra segue il moto dell’intero sistema solare e si muove nello spazio
a una velocità di circa 20,1 km/s nella direzione della costellazione di
Ercole; inoltre partecipa al moto di recessione della galassia, e insieme
alla Via Lattea si sposta verso la costellazione del Leone.
Oltre che dai moti principali, la Terra è interessata dal moto di
precessione degli equinozi e dalle nutazioni. Queste ultime sono
variazioni periodiche dell’inclinazione dell’asse terrestre, dovute alla
combinazione delle due forze di attrazione gravitazionale esercitate su di
essa dal Sole e dalla Luna.
Composizione della Terra
Anteprima della sezione
La Terra può essere schematicamente suddivisa, procedendo dall’esterno
verso l’interno, in cinque porzioni: l’atmosfera (gassosa), l’idrosfera
(liquida), la litosfera (solida), il mantello e il nucleo, in parte solidi.
L’atmosfera, costituita prevalentemente da azoto (N2) e ossigeno (O2), è
l’involucro gassoso che circonda il corpo del pianeta: ha uno spessore di
oltre 1100 km, ma data la rarefazione progressiva all’aumentare della
quota, circa la metà della sua massa è concentrata nei primi 5600 metri.
Età e origine della Terra
I metodi di datazione basati sullo studio dei radioisotopi hanno consentito
agli scienziati di stimare l’età della Terra in 4,65 miliardi di anni. Benché
le più vecchie rocce terrestri datate in questo modo non raggiungano i 4
miliardi di anni, alcune meteoriti, che sono simili geologicamente al
nucleo del nostro pianeta, risalgono a circa 4,5 miliardi di anni fa e si
ritiene che la loro cristallizzazione sia avvenuta approssimativamente 150
milioni di anni dopo la formazione della Terra e del sistema solare.
Il nostro pianeta, subito dopo la sua formazione (avvenuta probabilmente
per aggregazione gravitativa di materia libera nello spazio), doveva
essere un corpo quasi omogeneo e relativamente freddo. La contrazione
gravitazionale provocata dal progressivo accrescimento della sua massa
produsse un aumento di temperatura, al quale contribuì senza dubbio il
decadimento radioattivo di alcuni isotopi. L’aumento di temperatura
giunse a un livello tale da innescare un processo di parziale fusione del
pianeta e la conseguente riorganizzazione dei suoi componenti in strati
concentrici – crosta, mantello e nucleo: i silicati, più leggeri, risalirono
verso la superficie della massa fluida, formando il mantello e la crosta,
mentre gli elementi pesanti, soprattutto ferro e nichel, affondarono
perlopiù verso il centro. Al tempo stesso, tramite le eruzioni vulcaniche,
gran parte dei gas leggeri furono espulsi dal mantello e dalla crosta.
Alcuni di questi gas, in particolar modo l’anidride carbonica e l’azoto,
andarono a costituire l’atmosfera primordiale, mentre il vapore acqueo
condensava, dando origine ai primi oceani.
Magnetismo terrestre
La Terra nel suo insieme si comporta come un enorme magnete. Il campo
magnetico terrestre, infatti, è molto simile a quello che si osserverebbe
collocando al centro del pianeta una barra magnetica con l’asse inclinato
di circa 11° rispetto all’asse di rotazione terrestre. Benché gli effetti del
geomagnetismo siano noti e sfruttati da molte centinaia di anni (ad
esempio con la bussola), i primi studi scientifici su questa proprietà del
nostro pianeta furono compiuti intorno al 1600 dal fisico e filosofo
britannico William Gilbert.
Poli magnetici
Il fatto che l’asse del campo magnetico terrestre non coincida con l’asse
di rotazione fa sì che anche i poli magnetici siano distinti da quelli
geografici. Il polo nord magnetico attualmente si trova al largo delle coste
occidentali delle isole Bathurst, nei Territori del Nord-Ovest canadesi,
quasi 1290 km a nord-ovest della baia di Hudson. Il polo sud magnetico
si trova invece sul bordo del continente antartico, nella zona di Terra
Adelia, circa 1930 km a nord-est di Little America.
La posizione dei poli magnetici non è fissa, ma muta in modo sensibile da
un anno all’altro. Il campo magnetico terrestre, infatti, varia in direzione
con una periodicità di circa 960 anni, e inoltre compie piccole variazioni
su scala giornaliera. Recenti studi effettuati sulla magnetizzazione fossile
dei sedimenti marini hanno rilevato un’ulteriore periodicità nelle
variazioni del campo geomagnetico, di 100.000 anni. Essa, secondo gli
scienziati, potrebbe essere legata alla variazione di eccentricità dell’orbita
terrestre, che avviene appunto secondo un ciclo di 100.000 anni.
I dati raccolti dai satelliti rivelano che per il campo magnetico terrestre è
in corso da circa 150 anni un lento processo di indebolimento destinato a
risolversi con un’inversione di polarità. In sostanza, al termine di tale
processo, che dovrebbe durare circa due millenni, il Nord magnetico non
coinciderà più con il Nord geografico, ma con il Sud. Un campo
magnetico meno intenso, nel frattempo, potrebbe significare una
maggiore esposizione alle tempeste magnetiche provenienti dal Sole,
difficoltà nella navigazione dei satelliti e, in campo biologico, difficoltà
di orientamento per tutti gli animali che nelle migrazioni si affidano al
magnetismo – uccelli, farfalle, balene e molti altri.
Studi recenti del magnetismo residuo nelle rocce e delle anomalie
magnetiche dei fondi oceanici dimostrano inoltre come, negli ultimi 100
milioni di anni, si siano verificate almeno 170 inversioni di polarità del
campo magnetico terrestre. La conoscenza di queste inversioni, che
possono essere datate per mezzo degli isotopi radioattivi contenuti nelle
rocce, ha una grossa influenza sulle teorie della tettonica globale.
Elettricità terrestre
Sulla Terra e nell’atmosfera si manifestano fenomeni elettrici prodotti da
processi naturali. L’elettricità atmosferica, eccetto quella associata alle
cariche nelle nubi che genera i fulmini, deriva dalla ionizzazione prodotta
dalla radiazione solare e dal movimento di nubi di ioni trasportate dalle
maree atmosferiche; queste ultime sono prodotte, come le maree marine,
dall’attrazione gravitazionale del Sole e della Luna sull’atmosfera della
Terra. La ionizzazione (e quindi la conduttività elettrica) dell’atmosfera
in prossimità della superficie terrestre è bassa, ma aumenta rapidamente
con l’altitudine: tra i 40 e i 400 km la ionosfera forma un involucro
sferico quasi perfettamente conduttore che riflette le onde radio
permettendone la trasmissione a lunga distanza. La ionizzazione
dell’atmosfera varia molto anche a seconda dell’ora e della latitudine.
EVOLUZIONE
In biologia, l'evoluzione delle specie è il fenomeno del cambiamento, non
necessariamente migliorativo, del fenotipo, espressione visibile e diretta
del genotipo (cioè del patrimonio genetico) degli individui di una specie.
La teoria dell'evoluzione delle specie è uno dei pilastri della biologia
moderna. Nelle sue linee essenziali, essa è riconducibile all'opera di
Charles Darwin (che vide nella selezione naturale il motore fondamentale
dell'evoluzione della vita sulla Terra) e alla genetica.
Se i princìpi generali della teoria dell'evoluzione sono consolidati presso
la comunità scientifica, aspetti secondari della teoria sono tutt'oggi
ampiamente dibattuti, e costituiscono un campo di ricerca estremamente
vitale.
La definizione del concetto di evoluzione ha costituito una vera e propria
rivoluzione nel pensiero scientifico in biologia, e ha ispirato numerose
teorie e modelli in altri settori della conoscenza.
Uno dei primi disegni di Darwin
Sin da prima che Charles Darwin, il "padre" del moderno concetto di
evoluzione biologica, pubblicasse la prima edizione de L'origine delle
specie, le posizioni degli studiosi erano divise in due grandi correnti di
pensiero che vedevano, da un lato, una natura dinamica ed in continuo
cambiamento, dall'altro una natura sostanzialmente immutabile (la Scala
Naturae di Linneo). Della prima corrente facevano parte scienziati e
filosofi vicini all'Illuminismo francese, come Maupertuis, Buffon, La
Mettrie, che rielaboravano il meccanismo di eliminazione dei viventi
malformati proposto da Lucrezio nel De rerum natura ed ipotizzavano
una derivazione delle specie le une dalle altre. Tuttavia, l'interpretazione
di tali teorie come veri e proprî preannunci di evoluzionismo è
discussa.[1]
In ogni modo, ancora alla fine del 1700 la teoria predominante era quella
"fissista" dello scienziato Linneo, che definiva le varie specie come entità
create una volta per tutte e incapaci di modificarsi o capaci entro ben
determinati limiti.
Su questo tema oggi il mondo scientifico non è più diviso: le scoperte di
Mendel e Morgan nel campo della genetica, i progressi della
paleontologia e della biogeografia hanno conferito validità scientifica alla
teoria dell'evoluzione delle specie.
Il dibattito si è così spostato su un altro tema: ci si interroga sulle
modalità e le dinamiche dell'evoluzione e quindi sulle teorie che la
possono spiegare.
Oggi sappiamo che l'evoluzione delle specie è avvenuta in seguito a
trasformazioni, selezionate poi dall'ambiente; per arrivare a questa
affermazione ci sono voluti molti anni.
All'inizio del XIX secolo iniziarono a sorgere, negli studiosi di Scienze
Naturali i primi dubbi concreti: negli strati rocciosi più antichi infatti
mancano totalmente tracce (fossili) degli esseri attualmente viventi e se
ne rinvengono altre appartenenti ad organismi attualmente non esistenti.
Nel 1809, il naturalista Lamarck presentò per primo una teoria
evoluzionista (detta lamarckismo) secondo cui gli organismi viventi si
modificherebbero gradualmente nel tempo adattandosi all'ambiente: l'uso
o il non uso di determinati organi porterebbe con il tempo ad un loro
potenziamento o ad un'atrofia. Tale ipotesi implica quello che oggi viene
considerato l'errore di fondo: l'ereditabilità dei caratteri acquisiti
(esempio: un culturista non avrà necessariamente figli muscolosi; la
muscolosità del culturista è infatti una manifestazione fenotipica, cioè
morfologica, derivante dall'interazione dello sportivo con l'ambiente, il
continuo sollevare pesi; ma il particolare sviluppo muscolare non è
dettato dal suo patrimonio genetico, il genotipo).
Lamarck trovò opposizione in Georges L. Chretien Cuvier, il quale aveva
elaborato la 'teoria delle catastrofi naturali' secondo la quale la maggior
parte degli organismi viventi nel passato sarebbero stati spazzati via da
numerosi cataclismi e il mondo infatti sarebbe stato ripopolato dalle
specie sopravvissute.
Dopo cinquant'anni Darwin formulò una nuova teoria evoluzionista; il
noto naturalista, durante il suo viaggio giovanile sul brigantino Beagle, fu
colpito dalla variabilità delle forme viventi che aveva avuto modo di
osservare nei loro ambienti naturali intorno al mondo. Riflettendo sugli
appunti di viaggio e traendo spunto dagli scritti dell'economista Thomas
Malthus, Darwin si convinse che la “lotta per la vita” fosse uno dei
motori principali dell'evoluzione intuendo il ruolo selettivo dell'ambiente
sulle specie viventi. L'ambiente, infatti, non può essere la causa primaria
nel processo di evoluzione (come invece sostenuto nella teoria di
Lamarck) in quanto tale ruolo è giocato dalle mutazioni genetiche, in
gran parte casuali. L'ambiente entra in azione in un secondo momento,
nella determinazione del vantaggio o svantaggio riproduttivo che quelle
mutazioni danno alla specie mutata, in poche parole, al loro migliore o
peggiore adattamento (fitness in inglese).
I principali meccanismi che partecipano in queste situazioni sono:
•
meccanismi genetici
•
meccanismi ecologici
Neodarwinismo: la sintesi moderna
La moderna teoria dell'evoluzione (detta anche sintesi moderna o
neodarwinismo) è basata sulla teoria di Charles Darwin, che postulava
l'evoluzione delle specie attraverso la selezione naturale, combinata con
la teoria di Gregor Mendel sulla ereditarietà biologica. Altre personalità
che hanno contribuito in modo importante allo sviluppo della sintesi
moderna sono: Ronald Fisher, Theodosius Dobzhansky, J.B.S. Haldane,
Sewall Wright, Julian Sorell Huxley, Ernst Mayr,George Gaylord
Simpson e Motoo Kimura.
La maggior parte dei biologi sostiene la tesi della discendenza comune:
cioè che tutta la vita presente sulla Terra discenda da un comune
antenato. Questa conclusione si basa sul fatto che molte caratteristiche
degli organismi viventi, come il codice genetico, in apparenza arbitrari,
sono invece condivisi da tutti gli organismi anche se qualcuno ha
ipotizzato origini multiple della vita.
I rapporti di discendenza comune tra specie o gruppi di ordine superiore
si dicono rapporti filogenetici, e il processo di differenziazione della vita
si chiama filogenesi. La paleontologia dà prove consistenti di tali
processi.
Organi con strutture interne radicalmente diverse possono avere una
somiglianza superficiale e avere funzioni simili: si dicono allora analoghi.
Esempi di organi analoghi sono le ali degli insetti e degli uccelli. Gli
organi analoghi dimostrano che esistono molteplici modi per risolvere
problemi di funzionalità. Nello stesso tempo esistono organi con struttura
interna simile ma che servono a funzioni radicalmente diverse (organi
omologhi).
Confrontando organi omologhi di organismi dello stesso phylum, ad
esempio gli arti di diversi Tetrapodi, si nota che presentano una struttura
di base comune anche quando svolgono funzioni diverse, come la mano
umana, l'ala di un uccello e la zampa anteriore di una lucertola. Poiché la
somiglianza strutturale non risponde a necessità funzionali, la spiegazione
più ragionevole è che tali strutture derivino da quella del comune
progenitore. Inoltre, considerando gli organi vestigiali, risulta difficile
ammettere che siano comparsi fin dall'inizio come organi inutili, mentre
se si ammette che avessero una funzione in una specie progenitrice la loro
esistenza risulta comprensibile.
La mutazione (termine introdotto all'inizio del Novecento) consiste nella
comparsa improvvisa, casuale ed ereditabile nelle future generazioni, di
caratteristiche non possedute da antenati degli individui che le
presentano. La ricombinazione genetica, che permette di creare nuove
combinazioni di caratteristiche ereditarie, può aver luogo sia durante la
meiosi (riproduzione sessuata) sia per trasferimento di materiale genetico
da una cellula all'altra (coniugazione o trasformazione batterica).
Con cladismo si intende la ramificazione evolutiva già figurata da Darwin
nell'Origine della specie del 1859. Attualmente fonda la classificazione
sulla prospettiva filogenetica. La paleontologia aiuta a comprendere con
numerosi esempi come una specie madre possa dare origine a due o più
specie figlie, per ramificazione dicotomica, utilizzando la distinzione fra
caratteri primitivi e innovativi.
Sopravvivenza differenziata delle caratteristiche [modifica]
Con questo termine si intende quali caratteristiche sono presenti in una
popolazione e se la frequenza di presenza aumenta o diminuisce (anche
fino alla totale scomparsa). Due processi fondamentali determinano la
sopravvivenza di caratteristiche: la selezione naturale e la deriva genetica.
La selezione naturale è il fenomeno per cui organismi della stessa specie
con caratteristiche differenti ottengono, in un dato ambiente, un diverso
successo riproduttivo; di conseguenza, le caratteristiche che tendono ad
avvantaggiare la riproduzione diventano più frequenti di generazione in
generazione. Si ha selezione perché gli individui hanno diversa capacità
di utilizzare le risorse dell'ambiente e di sfuggire a pericoli presenti (come
predatori e avversità climatiche); infatti le risorse a disposizione sono
limitate, e ogni popolazione tende ad incrementare la sua consistenza in
progressione geometrica, per cui i cospecifici competono per le risorse
(non solo alimentari).
È importante notare che mutazione e selezione, prese singolarmente, non
possono produrre un'evoluzione significativa.
La prima, infatti, non farebbe che rendere le popolazioni sempre più
eterogenee. Inoltre, per il suo carattere casuale, nella maggior parte dei
casi essa è neutrale, oppure nociva, per la capacità dell'individuo che la
esibisce di sopravvivere e/o riprodursi.
La selezione, dal canto suo, non può introdurre nella popolazione nessuna
nuova caratteristica: tende anzi ad uniformare le proprietà della specie.
Solo grazie a sempre nuove mutazioni la selezione ha la possibilità di
eliminare quelle dannose e propagare quelle (poche) vantaggiose.
L'evoluzione è quindi il risultato dell'azione della selezione naturale sulla
variabilità genetica creata dalle mutazioni (casuali, ovvero indipendenti
dalle caratteristiche ambientali). L'azione della selezione naturale e delle
mutazioni viene analizzata quantitativamente dalla genetica delle
popolazioni.
È anche importante sottolineare che la selezione è controllata
dall'ambiente, che varia nello spazio e nel tempo e comprende anche gli
altri organismi.
Le mutazioni forniscono perciò il meccanismo che permette alla vita di
perpetuarsi. Infatti gli ambienti sono in continuo cambiamento e le specie
scomparirebbero se non fossero in grado di sviluppare adattamenti che
permettono di sopravvivere e riprodursi nell'ambiente mutato.
Deriva genetica
La deriva genetica è la variazione, dovuta al caso, delle frequenze
geniche in una piccola popolazione. Nelle piccole popolazioni derivanti
da una più vasta è anche importante l'"effetto del fondatore", per cui esse
possono avere casualmente frequenze geniche significativamente diverse
da quelle della popolazione originaria.
Grazie a questi due fenomeni piccole popolazioni possono "sperimentare"
combinazioni genetiche improbabili in quelle grandi.
Affinché specie oggi distinte possano discendere da un progenitore
comune è necessario che le specie in qualche modo "si riproducano". Ciò
richiede che una parte della specie subisca un'evoluzione divergente dal
resto, in modo che ad un certo punto si siano accumulate tante variazioni
da poterla considerare una specie distinta.
Ogni specie (a meno che non sia in via di estinzione o residuale) è
formata da più popolazioni mendeliane. Esse non coincidono con le
popolazioni ecologiche e sono definite come parti della specie al cui
interno si ha un'ampia possibilità di incrocio. La speciazione è possibile
quando tra popolazioni o gruppi di popolazioni si instaura un isolamento
riproduttivo, ossia vi è uno scambio genetico pressoché nullo.
Se si realizza l'isolamento per un tempo abbastanza lungo, è impossibile
che per puro caso si abbia la stessa evoluzione nelle due parti della
specie. La divergenza evolutiva è ancor più marcata se i due gruppi
vivono in ambienti diversi poiché la selezione agisce su di loro in modo
diverso
Speciazione allopatrica
La speciazione allopatrica avviene quando l'evoluzione di parti diverse
della specie madre avviene in territori diversi. È necessario che l'areale
della specie sia discontinuo, ossia che sia diviso in porzioni disgiunte,
separate da zone in cui la specie non può vivere. Si ha quindi un
isolamento geografico.
Più che l'isolamento geografico, il meccanismo di speciazione allopatrica
sembra principalmente legato all'isolamento periferico: in seno ad una
piccola subpopolazione, vivente ai margini dell'areale della specie in
condizioni non ottimali, avviene la rapida differenziazione evolutiva e
segregazione di una nuova specie in seguito al limitato scambio genetico
con la popolazione principale.
Speciazione simpatrica
Si ha speciazione simpatrica quando due popolazioni si evolvono
separatamente
pur vivendo
nello
stesso
territorio.
L'isolamento
riproduttivo senza separazione geografica si può avere in due modi.
•
L'isolamento ecologico è dovuto al fatto che le popolazioni
occupano nicchie ecologiche differenti. Un esempio classico sono i
fringuelli delle Galápagos, che han dato origine a specie diverse
per alimentazione. Questo esempio non è ritenuto corretto dalla
totalità degli ambienti scientifici, infatti, si potrebbe obiettare che
la distanza tra le isole è una sorta di separazione geografica (quindi
rientrerebbe nella categoria della speciazione allopatrica).
•
L'isolamento genetico è causato da riarrangiamenti cromosomici
stabilizzatisi in un piccolo gruppo, che non si può più incrociare
con i cospecifici pur avendo inizialmente lo stesso fenotipo
(criptospecie).
Prove
Oggi l'evoluzione è considerata, dalla stragrande maggioranza dei
biologi, un "fatto" supportato da una mole impressionante di prove di
varia natura.
Si tratta, perlomeno sino ad oggi, della migliore spiegazione scientifica
(quindi falsificabile) della diversità dei viventi.
Prove paleontologiche
Esempio di successione evolutiva
La successione degli ammoniti Hildoceratidi del Lias superiore
(Giurassico) nell'Appennino umbro-marchigiano, mostra continue
variazioni verticali (ossia nel tempo) con graduali modificazioni nella
morfologia delle ammonite presenti, variazioni che sono state
interpretate come evolutivi passaggi tra genere e genere.
Qui, all'interno dell'unità litostratigrafica del Rosso Ammonitico, è
presente una serie di ammoniti, ben conservate, raccogliendone
sistematicamente varie centinaia di campioni, strato per strato, si e'
osservato, muovendosi verso i termini più recenti, un adattamento
funzionale verso una sempre maggiore idrodinamicità, interpretata con
l'idea darwiniana della evoluzione gradualista per selezione naturale.
Trattasi di una microevoluzione simpatrica in quanto queste specie sono
presenti esclusivamente nell'area mediterranea della Tetide.
I dati della paleontologia mostrano non solo che gli organismi fossili
erano diversi da quelli attuali, ma anche che man mano che andiamo
indietro nel tempo le differenze con gli organismi viventi sono maggiori.
Ad esempio, fossili abbastanza recenti possono essere attribuiti
generalmente a generi attuali, mentre quelli man mano più antichi sono
sempre più diversi e sono attribuibili ad altri generi; permangono talora
caratteristiche di base, per cui possono essere spesso attribuiti agli stessi
gruppi tassonomici di ordine elevato attuali.
Ciò si accorda bene con l'ipotesi generale, che, arretrando nel tempo, ci si
avvicina alla radice dell'albero filogenetico.
La paleontologia fornisce prove concrete dell'evoluzione, quando i fossili
sono trovati nelle successioni stratigrafiche sedimentarie in abbondanza,
laddove è rispettato il principio fondamentale geologico della
sovrapposizione. I fossili dentro le rocce sedimentarie marine sono diffusi
in tutte le parte del mondo e permettono indagini stratigrafiche molto
dettagliate.
Prove biogeografiche
La distribuzione geografica delle specie viventi, anche alla luce delle
conoscenze sulla deriva dei continenti, ben si accorda con l'evoluzione
organica. L'enorme varietà di adattamenti dei marsupiali australiani, ad
esempio, può essere spiegata col fatto che la separazione dell'Australia
dagli altri continenti precede la comparsa degli euteri, per cui i marsupiali
terrestri australiani hanno potuto adattarsi a nicchie ecologiche per cui
non dovevano competere con altri ordini di mammiferi.
Anche la presenza di grossi uccelli non volatori in grandi isole porta alle
medesime conclusioni. Infatti, visto che esse erano già separate dai
continenti alla comparsa degli omeotermi, solo gli uccelli hanno potuto
raggiungerle ed occupare nicchie terrestri solitamente occupate da
mammiferi.
Alle
prove
biogeografiche
si
possono
aggiungere
quelle
paleobiogeografiche. La paleobiogeografia si occupa della posizione
paleogeografica dei fossili, a partire da quella geografica attuale.
L'argomento ha enorme importanza quando i fossili sono molto antichi
(per es. quelli del Paleozoico e del Mesozoico), e talora danno indizi di
speciazione allopatrica per migrazione. Tali studi, ancora poco sviluppati,
devono essere eseguiti con il concorso della biostratigrafia; in tal caso
possono dare risultati eccezionali. Un caso diverso è quello della presenza
degli stessi fossili in aree oggi separate; Sudamerica e Africa infatti
presentano in successioni rocciose simili, di origine continentale, fossili
di rettili sinapsidi simili del Permiano, 250 milioni di anni fa, a
testimoniare che i due continenti erano uniti nel supercontinente
Gondwana in quel lontano periodo.
Prove matematico/informatiche
Gli algoritmi genetici sono delle metaeuristiche per la ricerca della
soluzione ottimale di un problema basate sulla logica del modello
evoluzionistico. Studiando questo metodo si è visto come, partendo dalle
ipotesi del modello evoluzionistico, si può arrivare all'evoluzione di più
specie.
Sono stati realizzati molti programmi per computer che simulano un
ecosistema per diversi scopi (divertimento, studio dei meccanismi
evolutivi naturali, studio degli algoritmi genetici). Anche questi hanno
dimostrato la plausibilità del modello evoluzionistico. Inoltre, gli
algoritmi genetici sono stati applicati in campi lontani dalla biologia,
come i problemi di ottimizzazione di funzioni matematiche, in cui le
soluzioni vengono fatte "competere" e "incrociare" tra di loro con
particolari metodi.
Evoluzione osservabile
Uno dei pochi fenomeni di evoluzione osservabili, per via della estrema
brevità dei cicli vitali in gioco e quindi della rapidità con cui è possibile
osservare la successione delle generazioni, è quello relativo alla
progressiva resistenza agli antibiotici da parte dei batteri. È necessario
utilizzare sempre nuovi antibiotici per assicurare trattamenti efficaci e ciò
è dovuto al fatto che i batteri, come tutte le specie, mutano, e in un
ambiente a loro ostile come un corpo umano in terapia antibiotica,
sopravvivono
semplicemente
quegli
individui
le
cui
mutazioni
determinano una maggiore resistenza a quello specifico antibiotico. L'uso
diffuso degli antibiotici (sia sugli uomini che sugli animali) non fa che
selezionare i ceppi batterici più resistenti, con drammatica diminuzione
dell'efficacia. L'introduzione di un nuovo e più potente antibiotico non
farà che riproporre lo schema già descritto: tra le infinite mutazioni ve ne
saranno sempre alcune che daranno un vantaggio riproduttivo (che
renderanno cioè più "adatti") agli individui che le hanno subite.
Anche i virus mutano rapidamente, producendo sempre nuovi ceppi, cosa
che rende ancor più difficile cercare di contrastarli. Per questo motivo è
difficile riuscire a produrre vaccini definitivamente efficaci contro
l'influenza, visto che i tempi di mutazione del virus sono paragonabili ai
tempi necessari per mettere in commercio un vaccino.
L'impatto culturale del moderno concetto di evoluzione
L'evoluzionismo filosofico
Il concetto di evoluzione definito in biologia da Darwin è andato
estendendosi, nel tempo, come paradigma di intelligibilità applicabile a
tutta la storia dell'universo (vedi per esempio, in astrofisica, il concetto di
evoluzione stellare).
Anche le discipline umanistiche come la filosofia hanno recepito il
modello interpretativo evoluzionistico, così accanto alla versione
filosofica dell'evoluzione di tipo materialistico (quella di Herbert
Spencer), il concetto di evoluzione in filosofia portò anche a
reinterpretare le manifestazioni spirituali in senso evoluzionistico (due
esempi emblematici: il pensiero del filosofo e premio Nobel Henri
Bergson e del teologo gesuita, nonché paleoantropologo, Teilhard de
Chardin, i quali hanno utilizzato la teoria dell'evoluzione come uno
strumento utile a descrivere il ruolo del divino negli accadimenti della
storia).
Il nuovo approccio evoluzionistico nelle scienze umane
Anche Karl Marx dedicò Il capitale a Darwin, ritenendo il proprio studio
dell'economia in qualche modo corrispondente al darwinismo in biologia.
Anche l'antropologia culturale nello studio dell'evoluzione dei gruppi
umani e delle organizzazioni sociali trovò molto produttivo adottare il
punto di vista evoluzionista come è il caso dell'antropologia americana
con Lewis Henry Morgan e la sua scuola.
Critiche e alternative
Antievoluzionismo
Il concetto di Evoluzionismo ha ricevuto critiche sia per motivi
strettamente religiosi (Creazionismo biblico, professato dall'Ebraismo
ortodosso e da alcune Chiese protestanti americane, ma non più dalla
Chiesa cattolica, la quale non ha tuttavia una posizione unitaria e definita
sul darwinismo [v. Evoluzione e chiesa cattolica]), ma anche per opinioni
riguardanti l'adeguatezza del meccanismo esplicativo neodarwiniano, o
riguardanti la presunta insufficienza di prove della teoria di Darwin.
Malgrado queste opinioni siano decisamente minoritarie nella comunità
scientifica, e spesso non tengano conto di quanto la teoria originale di
Darwin sia evoluta nel tempo, è giusto ricordare alcune di queste teorie
alternative. Tra esse vi sono il Disegno intelligente e il Devoluzionismo
del biologo italiano Giuseppe Sermonti. La teoria di Darwin è anche
avversata dal fisico italiano Antonino Zichichi, che ne nega la solidità
matematica e ne attacca le prove biologiche e paleontologiche. D'altra
parte Zichichi ha criticato la teoria di Darwin solo su pubblicazioni
divulgative e non su riviste scientifiche sottostanti al meccanismo del
peer review.
Evoluzione e casualità
Il biologo Jacques Monod nel suo libro "Il caso e la necessità", asserì che
la teoria scientifica evoluzionistica andava intesa come una teoria che
concepiva l'evoluzione come una somma di eventi casuali, selezionati
dalle necessità ambientali, che nulla avrebbe quindi a che fare con
qualunque concezione finalistica sia riguardo all'uomo sia riguardo al
mondo.
La casualità evolutiva, che deriva dalla casualità delle modifiche naturali
del patrimonio genetico, che sono responsabili della differenziazione dei
diversi individui entro la singola specie, viene rigettata, con
considerazioni diverse dai sostenitori di una prospettiva finalistica
dell'evoluzione.
Evoluzione dell'uomo
Per l'evoluzione che dagli insettivori ha condotto all'homo sapiens
sapiens, e dall'H.s.s. in poi ha proseguito con la storia, vedere le voci
Evoluzione umana e Storia dell'uomo.
Evoluzione umana
L'evoluzione della vita sulla Terra, a quanto attualmente noto, parte circa
4 miliardi di anni fa.
Prima dell'uomo
Circa 70 milioni di anni fa (mya), proseguendo per un albero filogenetico
che affonda le radici alle origini della vita sulla terra, da esponenti
insettivori appartenenti alla classe dei mammiferi ebbe origine il ramo dei
primati, ordine di cui fanno parte con l'uomo tutte le scimmie. Nel
Miocene, da appartenenti a questa classe, 18 mya, (con Proconsul, un
arboricolo e frugivoro candidato ad entrare nella biforcazione evolutiva)
si diramarono le attuali scimmie antropomorfe, (gibbone 18 mya, orango
14 mya, gorilla 7 mya, scimpanzé e bonobo 3-5 mya), attualmente riuniti
con l'uomo in un'unica famiglia. Ardipithecus ramidus e Ardipithecus
kadabba
paiono
essere
anelli
importanti
nella
transizione
ad
australopiteco, mentre Kenyanthropus platyops sembra fondamentale per
spiegare la successiva transizione ad Homo.
Secondo un recente studio [1] l’andatura bipede è molto più antica di
quanto si pensasse. Alcuni fossili di Morotopithecus bishopi, un primate
arboricolo vissuto circa 21 milioni di anni fa nell’attuale Uganda,
presentano nella struttura dello scheletro e delle vertebre forti analogie
con le caratteristiche che nell’essere umano consentono di assumere la
posizione eretta. Queste analogie potrebbero essere dovute a convergenza
evolutiva, giacché lo stato attuale delle conoscenze (anche a causa della
frammentarietà dei resti fossili) non permette di chiarire questo dubbio.
Circa 20-15 milioni di anni fa, gli ominidi iniziarono a vagare per le
savane in cerca di cibo: qui la pressione selettiva favorì quegli individui
capaci di ergersi sugli arti posteriori potendo così, ad esempio, avvistare
in anticipo un predatore. Iniziò così l'evoluzione fisiologica e culturale di
questi primati: impararono infatti ad afferrare, trasportare, scegliere
piante e cibo ed osservare la natura.
Scala in migliaia di anni
Le età degli spazi * sono stimati. - I tratti verticali rappresentano le
possibili separazioni (ipotesi "splitter", dell'origine unica)
(1) o Homo sapiens arcaico antico - (2) o Homo sapiens arcaico recente
•
Pierolapithecus catalaunicus (13 milioni di anni fa) [specie ancora
in fase di studio]
o
•
"Pau"
Oreopithecus bambolii (8,5 milioni di anni fa) [anche se la sua
appartenenza al ramo umano è controversa]
o
•
"Proto" e "Sandrone"
Sahelanthropus tchadensis (fra 7 e 6 milioni di anni fa)
o
"Toumaï"
•
Orrorin tugenensis (6 milioni di anni fa)
•
Ardipithecus kadabba (fra 6 e 5,5 milioni di anni fa)
•
Ardipithecus ramidus (4,5 milioni di anni fa)
La specie più antica che conosciamo è quella dell'australopiteco, cioè le
scimmie dell'emisfero australe, che quasi sicuramente vissero in Tanzania
ed in Etiopia per almeno 3 milioni di anni, finché non si estinsero circa 1
milione di anni fa. L'australopiteco non era capace di costruire utensili,
ma utilizzava ciottoli per scopi semplici come spezzare o percuotere;
inoltre faceva vita di gruppo, dava la caccia ad animali di piccola stazza e
raccoglieva uova e semi.
•
Australopithecus anamensis (4 milioni di anni fa)
•
Kenyanthropus platyops (3,5 milioni di anni fa)
•
Australopithecus afarensis (fra 4 e 3 milioni di anni fa)
o
"Lucy"
•
Australopithecus bahrelghazali (fra 3,5 e 3 milioni di anni fa)
•
Australopithecus africanus (fra 3 e 2 milioni di anni fa)
•
Australopithecus garhi (2,5 milioni di anni fa)
•
Australopithecus aethiopicus - Parantropo (2.5 milioni di anni fa)
•
Australopithecus boisei - Parantropo (fra 1.7 ed 1.4 milioni di anni
fa)
•
Australopithecus robustus - Parantropo (fra 2 ed 1.5 milioni di anni
fa)
Distribuzione temporale e geografica delle popolazioni di ominidi, basata
sui fossili rinvenuti
La prima specie del genere homo conosciuta è l' Homo habilis (ca 2
milioni di anni fa). Molto simile all'australopiteco, l' Homo habilis viene
già ritenuto uomo per le sue abilità manuali: utilizzava infatti strumenti
rudimentali per la caccia.
Un'evoluzione arriva con l' Homo erectus (ca 1 - 1,5 milioni di anni fa).
L' erectus ha posizione eretta e una maggior capacità intellettiva,
testimoniata anche dal maggior sviluppo della tecnologia rispetto
all'homo habilis.
•
Homo habilis (fra 2,5 ed 1 milione di anni fa)
•
Homo rudolfensis (2 milioni di anni fa)
•
Homo ergaster (fra 2 milioni e 600.000 anni fa)
•
Homo erectus (fra 2 milioni e 300.000 anni fa)
o
"Argil", o Uomo di Ceprano
•
Homo antecessor (800.000 anni fa)
•
Homo heidelbergensis (fra 600.000 e 200.000 anni fa)
o
"Ciampate del Diavolo"
•
Homo neanderthalensis (fra 250.000 e 30.000 anni fa)
•
Homo floresiensis (da 95.000 a 18.000 anni fa)
o
•
"Ebu", o Uomo di Flores
Homo sapiens (da 200.000 anni fa ad oggi)
Tabella comparativa delle specie Homo
I nomi delle specie in grassetto indicano l'esistenza di numerosi fossili.
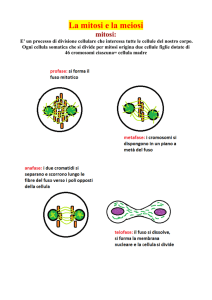
LA CELLULA ANIMALE E VEGETALE
La cellula (dal latino, piccola camera) è l'unità fondamentale di tutti gli
organismi viventi[1], la più piccola struttura ad essere classificabile come
vivente.
Alcuni organismi, come ad esempio i batteri acidoplastici o i protozoi,
possono consistere di una singola cellula ed essere definiti unicellulari.
Altri organismi, come l'uomo (formato da circa 100 mila miliardi di
cellule), sono invece pluricellulari. I principali organismi pluricellulari
appartengono tipicamente ai regni animale, vegetale e dei funghi. Le
cellule degli organismi unicellulari presentano caratteri morfologici
solitamente uniformi. Con l'aumentare del numero di cellule di un
organismo, invece, le cellule che lo compongono si differenziano in
forma, grandezza, rapporti e funzioni specializzate, fino alla costituzione
di tessuti ed organi.
Il termine cellula è legato all'analogia che Robert Hooke immaginò tra le
microstrutture che osservò nel sughero, utilizzando un microscopio di sua
invenzione, e le piccole camere che caratterizzano molti monasteri.
Ogni cellula può esser definita come un'entità chiusa ed autosufficiente:
essa è infatti in grado di assumere nutrienti, di convertirli in energia, di
svolgere funzioni specializzate e di riprodursi se necessario. Per fare ciò,
ogni cellula contiene al suo interno tutte le informazioni necessarie.
Tutte le cellule mostrano alcune caratteristiche comuni:Fotosintesi
clorofilliana
FOTOSINTESI CLOROFILLIANA
La fotosintesi clorofilliana è l’insieme delle reazioni durante le quali le
piante verdi producono sostanze organiche a partire da CO2 e dall’acqua,
in presenza di luce.
Mediante la clorofilla, l'energia solare (luce) viene trasformata in uno
zucchero definito glucosio fondamentale per la vita della pianta la cui
formula chimica è:C6H12O6,ovvero 6 atomi di carbonio,12 di idrogeno e 6
di ossigeno.Inoltre alla pianta(detta autotrofa)rimangono 6 atomi di
ossigeno atmosferico di cui si libera grazie agli stomi delle sue foglie.
Oggi questo processo è quello nettamente dominante sulla Terra, per la
produzione
di
composti
organici
da
sostanze
inorganiche
e,
probabilmente, rappresenta la prima forma di processo anabolico
sviluppato dagli organismi viventi. Inoltre, la fotosintesi è l'unico
processo biologicamente importante in grado di raccogliere l'energia solare,
da cui, fondamentalmente, dipende la vita sulla Terra.
Reazione complessiva
Il prodotto organico della fotosintesi ossigenica è il glucosio (C6H12O6),
il carboidrato monosaccaride più diffuso sul nostro pianeta. In seguito da
questo sono assemblate varie altre macromolecole, quali l'amido (la
forma di accumulo del carbonio nelle piante) e il saccarosio (la forma di
trasporto principale del carbonio nelle piante). Il carbonio e l'idrogeno
solfurato e fluorescente da convertire in sostanza organica sono forniti
rispettivamente
dall'anidride
carbonica
(CO2)
atmosferica
e
dall'acqua(H20). La quasi totalità della fotosintesi ossigenica è compiuta
da piante e alghe che ricavano l'idrogeno dall'acqua (H2O). In questo
caso l'equazione chimica che riassume il processo è:
6 CO2 + 6 H2O + 686 Kilocalorie/mole → C6H12O6 + 6 O2
Forme di fotosintesi
Esistono, soprattutto fra gli organismi procarioti autotrofi, varie forme di
fotosintesi, oltre alla fotosintesi clorofilliana ossigenica descritta qui. In
alcune specie di batteri autotrofi, l'idrogeno proviene non dall'acqua ma
dall'acido solfidrico, che nella fotosintesi viene ossidato a zolfo
elementare (S8)
6 CO2 + 12 H2S → C6H12O6 + 12 S + 6 H2O
Si noti che questi batteri sono anaerobi obbligati. Le forme di fotosintesi
clorofilliana che vengono effettuate con lo zolfo (o in alcuni casi anche
con l'azoto) vengono dette fotosintesi anossigeniche.
Anche fra le piante si riscontrano vari tipi di fotosintesi clorofilliana. Le
piante sono suddivise, in base alla forma di fotosintesi clorofilliana da
esse compiuta, in tre gruppi principali, che hanno diverse caratteristiche:
le piante C3, C4 e CAM. Vi è anche una forma di fotosintesi, la
chemiosintesi, in cui l'energia chimica è data dalla demolizione di
molecole organiche anziché dalla radiazione elettromagnetica.
Fasi della fotosintesi
La fotosintesi clorofilliana avviene per tappe riunibili in due fasi: la fase
luminosa (o fase luce-dipendente), dipendente dalla luce; la fase di
fissazione del carbonio (o fase oscura, indipendente dalla luce) di cui fa
parte il ciclo di Calvin.
La seconda fase viene anche definita fase al buio; il termine, tuttavia,
potrebbe essere fuorviante, in quanto non si riferisce all'assenza della luce
dato che alcuni enzimi coinvolti in questa fase sono direttamente attivati
proprio dalla luce, tanto che avviene contemporaneamente alla fase
luminosa e non di notte. Infatti in assenza di luce si ha scarsità di ATP e
NADPH, che si formano durante la fase luminosa e gli stomi si chiudono,
dunque non vi è accesso di CO2; inoltre si verifica anche l'inattività di
alcuni enzimi che sono luce-dipendenti (RuBisCO, 3-PGA deidrogenasi,
fosfatasi e ribulosio 1,5 bis-fosfato chinasi).
Fase luminosa
Le reazioni della fase luce dipendente della fotosintesi clorofilliana.
Il processo fotosintetico si svolge all'interno dei cloroplasti. All'interno di
questi si trova un sistema di membrane che formano pile di sacchetti
appiattiti (tilacoidi), dette grani, e delle lamelle di collegamento dei grani
(lamelle intergrana). All'interno di queste membrane troviamo delle
molecole di clorofilla, aggregate a formare i cosiddetti fotosistemi. Si
possono distinguere il fotosistema I e il fotosistema II. I fotosistemi sono
un insieme di molecole di pigmenti in cui l’energia viene convogliata
verso una molecola di clorofilla "a" trappola. Nel fotosistema I la
molecola trappola viene eccitata da una lunghezza d’onda di 700 nm, il
fotosistema II da 680 nm.
Il fotosistema I è formato da un LHC (complesso che cattura la luce)
costituito da circa 70 molecole di clorofilla a e b e da 13 diversi tipi di
catene polipeptidiche, e da un centro di reazione che comprende circa 130
molecole di clorofilla a e P700, un particolare tipo di clorofilla che ha il
massimo assorbimento della luce a 700nm.
Il fotosistema II è anch'esso composto da un LHC, formato da circa 200
molecole di clorofilla a e b, nonché da diverse catene polipeptidiche, e da
un centro di reazione formato da circa 50 molecole di clorofilla a e di
P680, che ha il massimo assorbimento della luce solare a 680nm.
Tutte queste molecole sono in grado di catturare l'energia luminosa, ma
solo quelle di clorofilla a sono in grado di passare ad uno stato eccitato
che attiva la reazione fotosintetica. Le molecole che hanno solo funzione
di captazione sono dette molecole antenna; quelle che attivano il processo
fotosintetico sono definite centri di reazione. La "fase luminosa" è
dominata dalla clorofilla a, le cui molecole assorbono selettivamente luce
nelle porzioni rossa e blu-violetta dello spettro visibile, attraverso una
serie di altri pigmenti coadiuvanti. L'energia catturata dalle molecole di
clorofilla consente la promozione di elettroni da orbitali atomici a energia
minore ad orbitali ad energia maggiore. Questi vengono subito sostituiti
mediante idrolisi di molecole d'acqua (che, da H2O, si scinde in due
protoni, due elettroni ed un ossigeno grazie alla fotolisi, operata dai due
fotosistemi). Gli elettroni liberati dalla clorofilla del fotosistema II
vengono immessi in una catena di trasporto costituita dal citocromo B6f,
durante la quale perdono energia, passando ad un livello energetico
inferiore. L'energia persa viene utilizzata per pompare protoni dallo
stroma all'interno dello spazio del tilacoide, creando un gradiente
protonico. Infine gli elettroni giungono al fotosistema I, che a sua volta,
per effetto della luce, ha perso altri elettroni. Gli elettroni persi dal
fotosistema I vengono trasferiti alle ferredossina, che riduce NADP+ in
NADPH. Tramite la proteina di membrana ATP-sintetasi situata sulla
membrana del tilacoide (strati membranosi interni al cloroplasto o, nel
caso dei batteri autotrofi, distribuiti nel citoplasma), gli ioni H+ liberatisi
dall'idrolisi dell'acqua passano dallo spazio del tilacoide allo stroma, cioè
verso gradiente, sintetizzando ATP a partire da gruppi liberi di fosfato e
ADP. Si può formare una molecola di ATP ogni due elettroni persi dai
fotosistemi.
Fase di fissazione del carbonio
La fase di fissazione del carbonio o ciclo di Calvin (chiamata anche fase
al buio o fase luce indipendente) comporta l'organicazione della CO2,
ossia la sua incorporazione in composti organici e la riduzione del
composto ottenuto grazie al ATP ricavato dalla fase luminosa.
In questo ciclo è presente un composto organico fisso, il ribulosiobifosfato, o RuBP, che viene trasformato durante la reazione fino a
tornare al suo stato iniziale. Le 12 molecole di ribulosio bifosfato presenti
nel ciclo di Calvin reagiscono con l'acqua e l'anidride carbonica subendo
una serie di trasformazioni ad opera dell'enzima ribulosio-bifosfato
carbossilasi o rubisco. Alla fine del processo, oltre alle 12 RuBP
nuovamente sintetizzate, si originano 2 molecole di gliceraldeide 3fosfato, che vengono espulse dal ciclo come prodotto netto della
fissazione. Per essere attivato, il ciclo di Calvin necessita di energia
chimica e supporto mediante l'idrolisi di 18 ATP in ADP e
dell'ossidazione di 12 NADPH in NADP+ e ioni liberi di idrogeno H+
(che sono protoni). L'ATP e la NADPH consumate durante il ciclo di
Calvin vengono prelevate da quelle prodotte durante la fase luminosa, e,
una volta ossidate, tornano a far parte del pool disponibile per la
riduzione. Complessivamente, nel ciclo di Calvin vengono consumate 6
molecole di CO2, 6 di acqua, 18 di ATP e 12 di NADPH per formare 2
gliceraldeide 3-fosfato (abbreviato in G3P), 18 gruppi liberi di fosfato, 18
ADP, 12 protoni, 12 NADP+.
Sintesi di glucosio
Le due molecole di gliceraldeide 3-fosfato formatesi durante il ciclo di
Calvin vengono utilizzate per sintetizzare glucosio, in un processo
perfettamente inverso alla glicolisi, o per formare lipidi quali acidi grassi
oppure amminoacidi (aggiungendo un gruppo amminico nella struttura). I
prodotti finali della fotosintesi, quindi, svolgono un ruolo di
fondamentale importanza nei processi dell'anabolismo degli organismi
autotrofi].
La divisione cellulare
La divisione cellulare è un processo importantissimo per la vita, in quanto
esso permette ad una cellula genitore (in inglese parent cell) si divida in
due o più cellule figlie (in inglese daughter cells).
Esistono fondamentalmente due tipi di divisione cellulare:
•
Divisione asessuata o per via vegetativa
o
fissione binaria ovvero la forma di riproduzione usata dagli
organismi procarioti. Questo processo esita nella formazione
di due parti, ognuna delle quali ha il potenziale di crescere
fino alle dimensioni della cellula originale. Ed ognuna delle
parti è un organismo completo.
•
o
divisione multipla
o
gemmazione
o
frammentazione
o
rigenerazione
o
clonazione
o
poliembrionia
o
sporulazione
Divisione sessuata
o
mitosi: ovvero divisione del nucleo genitore (in inglese
parent nucleo) in due nuclei figli (inglese daughter nucleus),
ognuno dei quali contiene un genoma identico al genoma
genitoriale.
o
meiosi: ovvero la divisione del nucleo in cellule sessuali, le
quali riducono il numero diploide di cromosomi ad un
numero aploide.
Nelle cellule eucariote multicellulari la mitosi permette la crescita e la
riparazione cellulare.
Il ciclo cellulare, o ciclo di divisione cellulare (CDC), è la serie di eventi
che avvengono in una cellula eucariote tra una divisione cellulare e quella
successiva. La durata del ciclo cellulare varia col variare della specie, del
tipo di cellula e delle condizioni di crescita.
Cenni generali
Il ciclo cellulare è un processo geneticamente controllato, costituito da
una serie di eventi coordinati e dipendenti tra loro, dai quali dipende la
corretta proliferazione delle cellule eucariotiche. Gli eventi molecolari
che controllano il ciclo cellulare sono ordinati e direzionali: ogni
processo è la diretta conseguenza dell'evento precedente ed è la causa di
quello successivo.
Molti geni coinvolti nella progressione del ciclo cellulare sono stati
individuati agli inizi degli anni settanta grazie ad uno studio condotto da
Lee Hartwell e collaboratori sul lievito Saccharomyces cerevisiae, un
microrganismo eucariotico unicellulare che si presta molto bene alle
analisi genetiche; grazie a questo lavoro furono isolati e caratterizzati
mutanti che presentavano alterazioni nelle diverse fasi del ciclo cellulare
(Hartwell, 1974).
Nelle cellule eucariotiche la progressione attraverso le varie fasi del ciclo
cellulare risulta essere finemente regolata dalle chinasi ciclina-dipendenti
o CDK (Cyclin-dependent Kinases) una famiglia di protein chinasi la cui
attività dipende dalla loro associazione con delle subunità proteiche
regolative dette cicline; queste ultime sono proteine instabili, sintetizzate
e degradate periodicamente, che si accumulano in fasi del ciclo specifiche
e che non solo attivano le CDK, ma ne determinano anche la specificità
di substrato.
Leland H. Hartwell, R. Timothy Hunt e Paul M. Nurse hanno vinto il
Premio Nobel per la Fisiologia e la Medicina nel 2001 per la loro
scoperta del ruolo centrale di queste molecole nel ciclo cellulare. Le
scoperte sono state ottenute studiando il ciclo cellulare rispettivamente
nel lievito gemmante Saccharomyces cerevisiae, nelle uova del riccio di
mare
Sphaerechinus
granularis
ed
nel
lievito
a
fissione
Schizosaccharomyces pombe
Negli eucarioti multicellulari la necessità di rispondere a una maggiore
quantità di stimoli esterni ed interni ha permesso l’evoluzione di
molteplici e diverse CDK: i vari complessi CDK - ciclina che si formano
durante il ciclo cellulare di tali organismi cambiano sia per quanto
riguarda la subunità regolatoria (ciclina) sia per quanto riguarda la
subunità catalitica (CDK). In ogni periodo del ciclo cellulare è presente
quindi un solo tipo di complesso CDK - ciclina cataliticamente attivo e, a
seconda del complesso formatosi, vengono fosforilate molecole bersaglio
differenti .
Oltre all’azione regolatoria della ciclina, il complesso CDK - ciclina è
anche soggetto all’azione di inibitori in grado di legarsi a tale complesso
e di renderne inattiva la subunità catalitica: questa classe di proteine
prende il nome di CKI (CDK Inhibitors). Inoltre, determinati siti della
subunità catalitica delle CDK risultano essere bersaglio di molte chinasi e
fosfatasi che, determinando lo stato di fosforilazione del complesso, ne
modulano più finemente la sua attività.
Fasi del ciclo cellulare
Il ciclo cellulare è un evento molto importante, per questo motivo è
regolato in tutte le sue dimensioni. Affinché l’informazione genetica
venga correttamente trasmessa dalla cellula madre alle cellule figlie, il
genoma deve essere prima duplicato durante il periodo di tempo
denominato fase S e in seguito i cromosomi devono venire segregati nelle
due cellule figlie durante la fase M. La fase M è a sua volta composta da
due processi, strettamente collegati: la mitosi, durante la quale i
cromosomi della cellula sono divisi tra le due cellule figlie e la
citodieresi, che comporta la divisione fisica del citoplasma della cellula.
Esiste una fase chiamata G0 (g zero) in cui la cellula ferma il suo ciclo
cellulare. Le cellule nervose e quelle muscolari (striate scheletriche)
rimangono in questo stadio per tutta la vita dell'organismo.
I punti di controllo
Il ciclo cellulare è un processo estremamente importante; errori in questo
processo potrebbero compromettere la vitalità cellulare. Per tale motivo,
nel ciclo cellulare, sono presenti dei punti di controllo o checkpoints,
localizzati a livello delle transizioni G1/S e G2/M. Infatti, tra le fasi S ed
M ci sono normalmente due periodi di tempo detti "gap": G1 fra la fine
della mitosi e l’inizio della fase S e G2 fra il termine della fase S e
l’inizio della fase M. In questi periodi di tempo si ha la maggior parte
della sintesi proteica con conseguente aumento della massa cellulare e la
realizzazione dei controlli che impediscono l’inizio della fase successiva
se non è stata completata quella precedente. Le fasi G1 e G2 sono quelle
che possono subire la maggior variabilità di durata e in alcuni casi
particolari possono anche essere eliminate, contrariamente alle fasi S e M
che sono essenziali e che rappresentano due eventi chiave del ciclo
cellulare. L'insieme delle fasi G1, S e G2 è collettivamente identificato
come interfase. Si dice che le cellule che hanno smesso di dividersi, in
modo temporaneo o irreversibile, sono in uno stato di quiescenza (fase
G0); le cellule che non vanno più incontro a divisione in modo
permanente, in seguito ad invecchiamento o a danni al DNA sono
chiamate senescenti. È da osservare che la mitosi produce sempre due
cellule geneticamente identiche alle cellula madre e che la maggior parte
degli organuli citoplasmatici si distribuisce a caso nelle cellule figlie.
Meccanismi molecolari comuni
Uno fra i cicli cellulari più semplice è quello del lievito Saccharomyces
cerevisiae, nel quale è presente una sola chinasi ciclina dipendente (CdK)
chiamata Cdc28 e due sole classi di cicline: G1 (Cln) e B (Clb). Nella
fase G1 del ciclo cellulare è attiva la trascrizione del gene che codifica
per una particolare ciclina di fase G1 della Cln3, che sembra agire da
sensore della massa cellulare. Infatti quando la cellula raggiunge la sua
massa critica, la concentrazione di questa ciclina aumenta, ed
associandosi con Cdc28 attiva un complesso programma trascrizionale
che comprende fra gli altri i geni codificanti per le cicline Cln1, Cln2,
Clb5 e Clb6. Ciò porta alla formazione dei complessi Cln1/2-Cdc28
responsabili della formazione della gemma (la cellula figlia) e della
duplicazione del corpo polare del fuso. Anche le cicline Clb5/6 si
associano alla CdK formando il complesso Clb5/6-Cdc28 la cui attività
chinasica è però bloccata attraversi il legame dell'inibitore Sic1. Quando
la concentrazione del complesso Cln1/2-Cdc28 ha raggiunto una soglia
critica, questo è in grado di fosforilare Sic1, indirizzandolo verso la
degradazione e permettendo l'attivazione dei complessi Clb5/6-Cdc28
sino a quel momento accumulati che a loro volta fosforilano e degradano
Sic1, mantenendone bassi i livelli. Questa attivazione è direttamente
responsabile dell'inizio della replicazione del DNA a livello delle origini
di replicazione, sulle quali è assemblato il complesso pre-replicativo (preRC). In seguito all'utilizzo dell'origine di replicazione, il complesso
Clb5/6-Cdc28 converte il pre-RC nel post-RC, impedendo che quella
stessa origine sia riutilizzata prima della successiva fase S. In tarda fase S
si ha la trascrizione dei geni codificanti per le cicline Clb3 e Clb4 ed alla
loro associazione con Cdc28, necessaria per l'entrata in mitosi e
l'allungamento del fuso mitotico, che in Saccharomyces cerevisiae
avviene immediatamente al termine della fase S. In altri organismi,
invece, a questo punto si ha la fosforilazione della CdK da parte della
chinasi Wee1 (presente anche in lievito), la quale inattiva i complessi
Clb-Cdc28 che si vanno via via accumulando. Quando giunge il segnale
di "via libera" la fosfatasi Cdc25 interviene eliminando il gruppo fosfato
sul complesso Clb-Cdc28, il quale attivandosi porta alla fosforilazione ed
inattivazione di Swe1, in un feedback positivo che produce un rapido
aumento della forma attiva di Clb-Cdc28, portando all'entrata in mitosi. Il
complesso Clb-Cdc28 quindi attiva il complesso promuovente l'anafase
(APC) il quale, grazie all'associazione con Cdc20, va ad indurre la
degradazione di una serie di proteine fra cui la securina (Pds1). La
degradazione di quest'ultima porta alla liberazione della separasi (Esp1)
che permette il taglio delle coesine che mantengono legati i cromatidi
fratelli, permettendone la migrazione ai poli opposti della cellula.
L'associazione dell'APC con la proteina Cdh1 induce inoltre la
degradazione di tutte le cicline di tipo B, provocando il crollo dell'attività
di Cdc28, con la conseguente uscita dalla mitosi. La perdita di attività di
Cdc28, privo di cicline, porta anche al riassemblamento dei complessi
pre-RC sulle origini di replicazione, alla possibilità di accumulare
nuovamente l'inibitore Sic1, nonché alla trascrizione delle cicline di fase
G1 (Cln)... ed un altro ciclo può ricominciare
Il ciclo cellulare negli eucarioti superiori [modifica]
Gateway G1/S
In questo punto, interviene il complesso cicline-CDK, il cui compito è
quello di determinare la iperfosforilazione ed attivazione della proteina
p105RB codificata dal gene RB1 (la cui mutazione può causare un
tumore dell'età pediatrica, il retinoblastoma (RB)). Le cicline implicate in
questo evento sono soprattutto le classi D1, D2, D3, della ciclina-D, la
quale viene prodotta durante la fase G1, ma nella fase S non è più
possibile ritrovarla. La ciclina-D, si lega alla CDK4, il complesso
risultante, determina la iperfosforilazione del RB ed il rilascio del fattore
di trascrizione E2F. Successivamente alla sua attivazione, il fattore di
trascrizione E2F viene rilasciato e favorisce l'entrata della cellula nel
ciclo cellulare. Inoltre, alcuni fattori di crescita come il TGF-β e la p53,
favoriscono l'aumento delle concentrazioni di p16, la quale blocca le
CDK ed il ciclo cellulare. Altri fattori di crescita come il PDGF e l'FGF,
favoriscono l'attivazione delle CDK.
Meccanismo d'azione del retinoblastoma
Il gene RB1 è un oncosoppressore localizzato sul cromosoma 13. La
proteina del retinoblastoma p105RB da esso codificata, può essere o nella
forma ipofosforilata (inattiva) o iperfosforilata (attiva). La forma inattiva
è caratterizzata dal fatto che p105RB mantiene complessato nella sua
struttura il fattore di trascrizione E2F, pertanto, il ciclo cellulare è inibito.
Mentre, la forma attiva è caratterizzata dal fatto che non mantiene
complessato nella sua struttura il fattore di trascrizione E2F, pertanto, il
ciclo cellulare è favorito. Nelle forme mutate del RB, il fattore di
trascrizione E2F è sempre libero, pertanto il ciclo cellulare non è
controllato.
Il fattore di trascrizione E2F, precedentemente rilasciato dal RB,
favorisce la formazione del complesso ciclina-A-CDK2 che agisce sia a
livello della mitosi che della profase, nella quale il complesso ciclina-BCDK1 favorisce la degradazione dell'involucro nucleare e consente
l'avvio della mitosi. Inoltre questi ultimi due complessi, comportano la
separazione
dei
centromeri
e
l'addossamento
dei
cromosomi.
Successivamente, la cellula va incontro a divisione.
Controllo della proliferazione cellulare
Negli organismi pluricellulari le cellule, normalmente in uno stato
quiescente, decidono di entrare in un nuovo ciclo cellulare in risposta ad
eventi biochimici fra i quali ci sono ad esempio, la ricezione attraverso il
meccanismo di trasduzione del segnale di fattori di crescita sintetizzati da
altre cellule.
Mitosi
La mitosi (mitòsi) (dal greco mìtos, che significa filo, per l'aspetto
filiforme dei cromosomi durante la metafase) è la riproduzione per
divisione equazionale della cellula eucariote. Il termine viene spesso
utilizzato anche per la riproduzione delle cellule procariote, un processo
molto più semplice e più correttamente chiamato scissione binaria o
amitosi.
La mitosi, nell'uomo, riguarda le cellule somatiche dell'organismo (cioè
tutte le cellule tranne quelle che hanno funzione riproduttiva, i gametociti
primari, che invece vanno incontro alla meiosi) e le cellule germinali
ancora indifferenziate.
Il processo inizia con la condensazione della cromatina che avviene
grazie alla presenza di proteine istoniche che fungono da centri primari di
organizzazione
del riavvolgimento del DNA, primo
ordine
di
spiralizzazione, e della topoisomerasi II che, oltre alla sua funzione
catalitica, agisce come centro di organizzazione del secondo ordine di
spiralizzazione; poi seguono un terzo ordine di cui non si conoscono le
proteine implicate e forse è conseguenza della tensione accumulata dalle
precedenti spiralizzazioni; poi questo grosso superfilamento viene prima
impaccato formando delle anse che poi si riuniscono formando il
cromosoma visibile. La durata media di questo meccanismo di
riproduzione cellulare varia in media, negli organismi superiori, tra le 15
e le 30 ore.
La mitosi nel ciclo cellulare
Il ciclo cellulare si suddivide in 3 parti: l'interfase, in cui la cellula si
prepara alla divisione; la mitosi, periodo di gran lunga più breve in cui la
cellula si divide che viene descritto nei particolari in questa pagina; il
periodo detto G0, più o meno definitivo, in cui la cellula si specializza
nella sua funzione e non è in grado o non è stimolata a riprodursi.
Il processo inizia con la condensazione della cromatina che avviene
grazie alla presenza di proteine istoniche che fungono da centri primari di
organizzazione
del riavvolgimento del DNA, primo
ordine
di
spiralizzazione, e della topoisomerasi II che, oltre alla sua funzione
catalitica, agisce come centro di organizzazione del secondo ordine di
spiralizzazione; poi seguono un terzo ordine di cui non si conoscono le
proteine implicate e forse è conseguenza della tensione accumulata dalle
precedenti spiralizzazioni; poi questo grosso superfilamento viene prima
impaccato formando delle anse che poi si riuniscono formando il
cromosoma visibile. La durata media di questo meccanismo di
riproduzione cellulare varia in media, negli organismi superiori, tra le 15
e le 30 ore.
Eventi che precedono la mitosi
Prima della mitosi avviene l'interfase, momento molto importante nella
vita della cellula, infatti proprio in questa fase, che si suddivide in G1, S e
G2, gli organelli della cellula aumentano e nella fase S il DNA si duplica.
La differenza sostanziale tra la sottofase G1 e la sottofase G2, è che la
sottofase G1 è interposta da dopo la mitosi precedente, e prima della
sottofase S, mentre la sottofase G2 avviene dopo la sottofase S e prima
dell'inizio della successiva mitosi
Profase
(pro- = prima)
I cromosomi ( nel caso della specie umana), si condensano e sono visibili
anche al microscopio ottico sotto forma di doppi bastoncelli basofili,
(infatti "mitosi" dal greco mitos significa "filo") i cromatidi gemelli, che
sono agganciati tra loro in un punto centrale detto centromero grazie ad
un complesso sistema di interazioni tra il DNA e numerose proteine
chiamatofuso
acromatico.
Viene sintetizzato un secondo centrosoma ed entrambi appaiono
circondati da una coltre di microtubuli(Fuso,formato da dimeri di subunita proteiche tubulina alfa e beta dissassemblando il citoscheletro). Il
ciclo si potrebbe interrompere in questo punto se alla coltura si
aggiungesse la tossina falloidina che impedisce la formazione dei
filamenti di microtubuli, questo si fa quando si vogliono visualizzare al
microscopio i cromosomi per studiarne le caratteristiche. L'apparato del
Golgi, il reticolo endoplasmatico in questa fase si scompongono in
piccole vescicolette che si distribuiscono uniformemente in tutto il
citoplasma; anche la membrana nucleare, grazie alla sua struttura a
doppia membrana si scompone similmente agli organelli citati prima.
Metafase
(meta- = durante)
Inizia anzitutto con una fase iniziale, detta prometafase, in cui avviene
l'improvvisa dissoluzione della membrana nucleare, che si frammenta in
tante vescicolette. Tale processo viene innescato dalla fosforilazione,
attraverso delle chinasi, delle proteine dei filamenti intermedi (lamìne)
che costituiscono la làmina nucleare; in conseguenza della fosforilazione
i filamenti si dissociano negli elementi costitutivi.
I due centrosomi si portano ai poli opposti della cellula ed agiscono come
centri di organizzazione microtubulare, catalizzando l'allungamento ed
assicurando il corretto orientamento dei microtubuli che andranno a breve
a legarsi al centromero di uno dei due cromatidi gemelli. In questa fase si
possono verificare degli errori e due microtubuli si possono agganciare
allo stesso cromatidio dando poi una cellula figlia mutilata e non vitale.
Questa fase viene chiamata anche prometafase, che significa prima parte
della metafase.
Le 23 coppie di cromatidi vengono portate nella parte mediana della
cellula, formando la "piastra equatoriale" in cui un piano immaginario,
passante per i centromeri, divide le coppie di DNA. È questo il momento
più favorevole per lo studio dei cromosomi, che sono ora al massimo
della loro spiralizzazione e affiancati ordinatamente lungo la piastra
equatoriale posta al centro della cellula.
Anafase
(ana- = ulteriore)
I cromatidi,migrano verso i due centrosomi ai poli opposti della cellula.
Si riconoscono due momenti detti anafase A e anafase B. Nella prima si
assiste alla separazione dei due cromatidi fratelli ad opera di un enzima
detto separasi con relativa migrazione degli stessi grazie a proteine
motore (tipo dineine citoplasmatiche) presenti a livello del cinetocore.
Nell'anafase B si assiste al reciproco scorrimento dei microtubuli polari
del fuso mitotico con conseguente allontanamento dei due centrosomi
verso direzioni opposte. Pertanto si ottiene il ripristino, per ogni polo, del
numero originario di cromosomi.
Telofase
(telo- = finale)
I cromosomi si despiralizzano. Intorno ai due nuovi complessi
cromosomici ricompaiono le membrane nucleari e gli organelli si
ricompongono. La telofase si conclude con una sottofase: la citodieresi,
in cui si separa il citoplasma in modo equivalente in entrambe le cellule.
La cellula si divide al centro formandone due cellule figlie, esattamente
identiche alla cellula madre ma più piccole. Questo avviene grazie ad un
anello di actina creatosi al centro della cellula madre che contraendosi
stringe la cellula al centro, a questo punto proteine specializzate operano
la fusione e la separazione della membrana in punti specifici e le due
cellule si separano.
In alcune cellule la telofase non avviene e si accumulano all'interno di
uno stesso nucleo di una stessa cellula da due ad alcune decine di corredi
cromosomici. Questo tipo di cellule si chiamano plasmodi e l'esempio
principale sono i protozoi del genere Plasmodium come il P. malariae;
anche cellule umane vanno incontro a questo processo o patologicamente,
come le cellule tumorali, o fisiologicamente come nel megacariocita e
forse (in attesa di una conferma dalla ricerca) in una minoranza di cellule
cardiache ed epatiche, anche i macrofagi iperattivati in un granuloma
vanno incontro a questa "mitosi mutilata".
L’importanza della mitosi consiste nel fatto che tutte le cellule
discendenti da una cellula originaria ereditano lo stesso numero di
cromosomi e risultano geneticamente identiche.
Mitosi come
modalità
di accrescimento
dell’organismo
Negli organismi pluricellulari, durante lo sviluppo embrionale, tutti i
blastomeri (cioè le cellule che derivano per la progressiva suddivisione di
uno zigote, ovvero per segmentazione), ereditano gli stessi cromosomi.
La mitosi costituisce in questi organismi la modalità di crescita
dell’individuo; le cellule che ne derivano subiscono poi un processo di
differenziamento che ne permette la specializzazione in differenti tessuti.
Quando l’individuo ha completato la sua crescita, però, non tutte le sue
cellule perdono la capacità di compiere mitosi e di suddividersi; ciò
permette la possibilità per alcuni tessuti di rinnovarsi rapidamente e di
riparare eventuali lesioni. Si possono distinguere, in tal senso, tre tipi di
cellule: quelle cellule soggette al rinnovamento, o cellule staminali, che
mantengono la capacità mitotica (ad esempio, le cellule dei meristemi
vegetali e le cellule dello strato germinativo dell’epidermide umana);
cellule in espansione, che hanno subito un processo di differenziamento
ma possono riprendere in alcuni casi la capacità mitotica (ad esempio, le
cellule che formano il callo dei tessuti vegetali lesionati, oppure gli
epatociti umani); infine, le cellule statiche, che non possono riprodursi e,
pertanto, se vengono danneggiate non possono essere sostituite (ad
esempio, le cellule del mesofillo fogliare o i neuroni).
MEIOSI
Meiosi Processo caratteristico delle cellule eucarioti, durante il quale da
una cellula si formano quattro cellule figlie, aventi la metà del patrimonio
genetico di quella originaria. In altri termini, la meiosi determina la
ripartizione di ciascuna coppia di cromosomi omologhi (cromosomi su
cui si trovano geni corrispondenti) presenti nelle cellule diploidi. La
meiosi si differenzia da un altro processo di divisione cellulare, la mitosi,
nella quale si formano cellule figlie aventi lo stesso patrimonio genetico
della cellula madre.
La meiosi avviene in una determinata fase del ciclo vitale degli
organismi. Negli organismi aplonti, caratterizzati da un patrimonio
genetico aploide (con un’unica copia di ciascun gene), la meiosi è detta
zigotica, perché interviene nello zigote diploide che si forma dopo la
fusione di due cellule aploidi; in tal modo, la meiosi interviene a formare
cellule aploidi da cui si svilupperanno per mitosi nuovi individui, e quindi
permette di ripristinare il patrimonio genetico della specie.
Negli organismi diplonti, caratterizzati da un patrimonio genetico
diploide (con due copie di ciascun gene), la meiosi è detta gametica,
perché si verifica in particolari cellule dell’organismo deputate alla
formazione di cellule riproduttive aploidi (gameti); i gameti permettono
l’attuazione della riproduzione sessuata e, fondendosi al momento della
fecondazione, formano zigoti diploidi da cui si sviluppano i nuovi
individui. La meiosi in questo caso è compresa nel processo di
gametogenesi, che porta dalla formazione dei gameti aploidi alla loro
completa maturazione. In particolare, i gameti maschili sono detti
spermatozoi e quelli femminili cellule uovo.
Negli organismi aplo-diplonti, la mitosi è detta intermedia: come nei
diplonti, la meiosi avviene in una cellula particolare (cellula gonotoconte
o zeugide) degli organismi della fase diploide (sporofiti) e porta alla
formazione di spore aploidi (meiospore); in questo caso, però, le spore
aploidi si moltiplicano per mitosi e danno origine a individui aplonti
(gametofiti).
Anteprima della sezione
La meiosi avviene secondo due fasi principali, dette rispettivamente
prima e seconda divisione meiotica, o meiosi I e meiosi II.
L’evento fondamentale della prima divisione meiotica è la ripartizione
dei due cromosomi di ciascuna coppia di omologhi nelle due cellule figlie
derivanti dalla cellula originaria: dunque, si ottengono due cellule dotate
ciascuna di un patrimonio genetico aploide, ovvero di un solo cromosoma
per tipo.
La cromatina visibile nel nucleo cellulare, che rappresenta la massa del
DNA quando la cellula svolge le sue normali attività metaboliche, si
condensa, in modo che si formano strutture bastoncellari, i cromosomi.
Ciascun cromosoma appare a forma di X (stadio di tetrade), poiché è
formato da due cromatidi fratelli, uniti in un punto detto centromero. I
cromatidi derivano da un processo di duplicazione del DNA; pertanto,
ciascuno è geneticamente identico all’altro. In questa fase, possono
avvenire scambi di frammenti di cromosoma tra i cromatidi dei due
cromosomi omologhi, fenomeno che prende il nome di crossing-over e di
fondamentale importanza per il mantenimento della variabilità genetica
tra individui della stessa specie. La membrana che avvolge il nucleo si
disgrega. Si forma un fascio di microtubuli proteici (fuso), che si estende
da un polo all’altro della cellula e le cui due estremità fanno capo a due
coppie di organuli, detti centrioli.
Le tetradi omologhe si dispongono simmetricamente lungo una
immaginaria linea equatoriale, trasversale rispetto al fuso. In tal modo,
ciascuna è rivolta verso uno dei due poli della cellula.
Le fibre del fuso prendono contatto con i centromeri; ciascuna tetrade
migra verso un polo della cellula.
Ai due poli della cellula madre si formano due agglomerati di cromosomi
aploidi, in cui è presente un solo cromosoma per ciascun tipo. I
cromosomi sono ancora allo stadio di tetrade. Il citoplasma delle due
cellule si ripartisce e avviene la citodieresi, ossia la vera e propria
divisione della cellula originaria in due cellule figlie distinte (in alcuni
casi, questa ripartizione può essere incompleta). Le fibre del fuso si
disgregano; i cromosomi si despiralizzano.
Nella seconda divisione meiotica, in modo analogo alla mitosi, i
cromatidi di ciascuna tetrade si separano e si ripartiscono in due cellule
figlie. Spesso la seconda divisione meiotica avviene immediatamente
dopo la prima divisione. Le due cellule che ne derivano risultano
anch’esse aploidi.
La cromatina si condensa nuovamente, in modo che si possono osservare
i cromosomi, formati da due cromatidi uniti dal centromero. Si forma
nuovamente il fuso di microtubuli.
I cromosomi si dispongono su una linea equatoriale, trasversale rispetto
alle fibre del fuso, in modo che ciascun cromatidio sia rivolto verso uno
dei due poli della cellula. I centromeri prendono contatto con le fibre.
I cromatidi migrano ciascuno verso un polo della cellula, spostandosi
verso le fibre del fuso. In tal modo, ciascun cromatidio diviene un nuovo
cromosoma.
Ai poli della cellula, si formano due aggregati di cromosomi. Le fibre del
fuso si disgregano, i cromosomi cominciano a decondensarsi, e si forma
infine una membrana nucleare. Il citoplasma della cellula si ripartisce in
due, così da portare alla formazione di due cellule figlie aploidi.
Da un punto di vista genetico, la meiosi assume una grande importanza
perché rappresenta il modo con cui possono formarsi nuove combinazioni
di geni e, quindi, rende possibile la variabilità genetica tra individui di
una stessa specie. Infatti, già con il crossing-over, ovvero con lo scambio
di porzioni di DNA tra cromatidi di due cromosomi omologhi, al
momento
della
profase
I,
avviene
una
prima
modificazione
dell’assortimento di geni rispetto a quello della cellula madre. Inoltre,
occorre considerare che la divisione dei due cromosomi omologhi (allo
stadio di tetrade) durante la fase di anafase I avviene in modo casuale: ciò
significa che non è prestabilito il polo della cellula verso cui migrerà
ciascun cromosoma. Dunque, a partire da una cellula madre, si formano
con la prima divisione meiotica due cellule aploidi che sono
geneticamente differenti tra loro e diverse da qualsiasi altra coppia di
cellule che derivano dalla stessa cellula madre. La diversità genetica
riguarda, precisamente, gli alleli presenti nel patrimonio di un individuo;
in altri termini, ciò che è suscettibile di variazioni non è il numero di
cromosomi o il numero di geni presenti (caratteristiche che devono
restare costanti perché tipiche della specie), ma il tipo di allele che, per
ciascun carattere, ciascuna cellula figlia può ereditare. La variabilità
genetica, assicurata anche dai meccanismi di mutazione spontanea,
assume un ruolo essenziale nei processi evolutivi, secondo il concetto di
selezione naturale.
Il metabolismo cellulare
Con tale termine si indicano i continui processi, sia chimici che fisici, cui
è soggetto il protoplasma e che danno luogo al continuo scambio di
energia e di sostanze tra l'ambiente esterno e la cellula stessa.
Si distingue:
a) anabolismo cellulare, in cui si comprendono tutti i processi per
mezzo dei quali la cellula si arricchisce di sostanze vitali per essa
e immagazzina complesse molecole chimiche fondamentali per la
sua evoluzione e per il suo trofismo;
b) catabolismo cellulare, con cui si intendono tutti i processi
distruttivi
cui
vanno
incontro
le
molecole
chimiche
precedentemente immagazzinate; distruzione che porta alla
formazione di energia con conseguente eliminazione dei rifiuti.
Tutti questi processi si possono raccogliere sotto un comun
denominatore: ricambio cellulare. L'assunzione di particelle solide
avviene per mezzo della fagocitosi. Tale proprietà venne studiata per la
prima volta sui leucociti di un mollusco da Haeckel, nel 1862, e consiste
nella emissione di pseudopodi (prolungamenti, dovuti ad estroflessioni
della membrana cellulare) o di membranelle ondulanti, per cui il
materiale da inglobare viene circoscritto da tali prolungamenti ed infine
inglobato entro il citoplasma.
In base alle proprietà fagocitanti, le cellule sono state distinte in
macrofagi e in microfagi: i primi sono in grado di assimilare cellule
batteriche al completo, i microfagi, invece, solo parti corpuscolate o
residui di cellule. Entrambi i tipi di cellule fagocitanti sono
abbondantemente rappresentati nel corpo umano. Le funzioni a cui tali
elementi sono deputati sono: difesa contro i germi e i microorganismi
patogeni in genere, eliminazione di pulviscolo atmosferico che attraverso
la respirazione perviene negli alveoli polmonari, eliminazione di detriti
da organi in via di decomposizione (come avviene per esempio nella
metamorfosi di alcuni animali) e infine l'assorbimento dei processi
infiammatori. Il meccanismo principale per mezzo del quale la cellula si
nutre avviene attraverso l'assorbimento di particelle liquide. A tale
riguardo, la membrana cellulare svolge una funzione fondamentale. Essa
si comporta infatti, come una membrana semipermeabile, come un filtro,
permettendo il passaggio di determinate sostanze e non di altre, anche se
con notevole affinità chimica. Si è notato inoltre che la disponibilità ad
assorbire della membrana varia a seconda dello stato funzionale in cui
essa si trova: per esempio, se la cellula, in un determinato stato, non avrà
bisogno di lipidi, pur potendoli assorbire, non li introdurrà, in quanto il
suo fabbisogno è per il momento espletato.
Ereditarietà dei caratteri
Le caratteristiche morfologiche, fisiologiche o psichiche di ogni
organismo vivente sono determinate da due fattori: il patrimonio
genetico ricevuto in eredità dai genitori e l’influenza ambientale. Il
primo determina i cosiddetti “caratteri ereditari” ossia da un lato
l’insieme delle caratteristiche generali che definiscono una determinata
specie e dall’altro i numerosi caratteri che differenziano tra loro
individui della stessa specie come ad esempio il colore degli occhi, il
gruppo sanguigno ecc. nell’uomo
La trasmissione dei caratteri ereditari obbedisce alle leggi formulate da
G. Mendel verso la fine del XIX secolo. La conclusione principale a cui
giunse Mendel è che il patrimonio genetico di ogni essere vivente, ossia
il suo “genotipo”, è composto dalla successione di coppie di elementi,
detti “geni”, che l’individuo riceve dai genitori al momento della
fecondazione tramite le cellule seminali. Ogni coppia di geni stabilisce
funzioni e proprietà di un dato organo (ad esempio il colore degli occhi)
mentre ogni gene della coppia è responsabile della manifestazione di un
carattere specifico (ad esempio il colore azzurro degli occhi). L’insieme
dei caratteri mostrati dall’individuo, ossia il suo “fenotipo”, è il risultato
della “competizione” che si stabilisce in ogni coppia tra i due geni
preposti alla medesima funzione. Quando una coppia contiene due geni
identici l’individuo manifesta il carattere trasmesso da entrambi i geni.
Se invece sono presenti geni contrapposti, l’individuo manifesta solo il
carattere del cosiddetto “gene dominate”. L’azione del secondo gene,
detto recessivo, risulta dunque mascherata.
Un carattere recessivo si manifesta quindi solo negli individui in
possesso di un genotipo recessivo puro. Questo spiega perché un
carattere recessivo si manifesta in genere con minor frequenza rispetto
al corrispondente carattere dominate. D’altra parte, due genitori del
fenotipo dominante possono generare figli che mostrano viceversa il
carattere recessivo. Ad esempio incrociando due piante di pisello giallo
si possono ottenere piante di pisello verde, un colore recessivo rispetto
al primo. Questo accade quando entrambi i genitori possiedono un
genotipo misto, avente cioè sia il gene dominante che quello recessivo.
Al momento della fecondazione, ogni genitore trasmette al figlio un
solo gene, in questo caso dominante o recessivo. Ogni gene è trasmesso
con uguale probabilità per cui se il figlio riceve dai entrambi i genitori il
gene recessivo, cosa che accade con una probabilità di uno su quattro,
mostrerà il carattere che nella generazione dei genitori risulta invisibile.
Infine l’osservazione, sempre dovuta a Mendel, che i vari caratteri
ereditari non sono in genere correlati tra loro, ad esempio il carattere
della buccia liscia o rugosa si trasmette in modo indipendente dal colore
di essa, completa il quadro scientifico sull’ereditarietà dei caratteri.
E’ opportuno tuttavia osservare che il quadro precedente è influenzato
da vari fattori che ostacolano in genere la verifica diretta delle leggi di
Mendel. Tra tutti ricordiamo il fenomeno della dominanza incompleta o
lo presenza di caratteri determinati da un numero di geni maggiore di
due. Questo si verifica ad esempio nell gruppo sanguigno dell’uomo
che può presentare i fenotipi A, B, AB o 0 e che viene determinato da
tre geni distinti caratterizzati da definiti rapporti di dominanza o
condominanza (due geni che contribuiscono in modo paritario al
fenotipo).
TESSUTO
Esempio di tessuto animale: tessuto nervoso.
In biologia si definisce tessuto un insieme di cellule simili per struttura e
funzione. Costituisce un livello superiore di organizzazione cellulare,
deputato a svolgere un ruolo determinante all'interno di un organismo, e
presente solo negli Animali e nelle Piante (in forma solo abbozzata nei
Poriferi e nelle Briofite). Negli animali superiori, spesso più tessuti
diversi si associano tra di loro a formare strutture ulteriormente
organizzate, gli organi. Un tessuto, nell'accezione corrente è un solido,
ma può essere ugualmente un fluido. Il sangue e la linfa sono anch'essi,
dal punto di vista anatomico, tessuti.
La scienza che studia i tessuti è chiamata istologia, ed una importante
branca della medicina e della biologia. Lo studio dei tessuti a scopo
prettamente diagnostico, invece, prende il nome di Anatomia patologica.
Lo strumento classicamente più utilizzato per studiare i tessuti è il
microscopio ottico; in tempi recenti, tuttavia, si è andato affermando
sempre di più l'uso del microscopio elettronico, dell'immunochimica
(l'utilizzo di anticorpi appositamente trattati che si legano ai diversi
componenti cellulari, permettendone l'identificazione), e di tecniche di
biologia molecolare e di genetica.
Tessuti animali
Esistono quattro tipi fondamentali di tessuti presenti in tutti gli animali,
dall'uomo ai più semplici invertebrati. Questi tessuti sono a loro volta
suddivisi in sotto-tipi, più specializzati, e, negli animali superiori, vanno a
costituire i diversi organi. I quattro tessuti fondamentali sono:
•
il tessuto epiteliale, o epitelio, costituito da cellule strettamente
ammassate e connesse tra loro, che costituisce il rivestimento di
tutte le superfici esterne ed interne del corpo, dei vasi sanguigni, e
che forma le ghiandole.
•
il tessuto connettivo, costituito da cellule di forma varia,
caratterizzate
dalla
presenza
di
una
abbondante
sostanza
intercellulare (o matrice) tra di esse. Come suggerisce il nome, la
funzione primaria di questo tessuto è quella di connettere, sia
strutturalmente che funzionalmente, gli altri tessuti e gli organi.
Comunque, il tessuto connettivo si differenzia in numerosi sottotipi, che esplicano a loro volta funzioni molto varie. Alcuni esempi
di questi sotto-tipi, oltre al tessuto connettivo propriamente detto,
sono: il tessuto cartilagineo, il tessuto osseo, il tessuto adiposo ed il
sangue.
•
il tessuto muscolare, costituito da cellule conteneti numerosi
filamenti contrattili, capaci di scorrere fisicamente gli uni sugli altri
e di cambiare la forma delle cellule stesse. Il tessuto muscolare
permette
il
movimento
dell'organismo,
e
la
contrazione
involontaria di diversi organi o apparati. Si divide in tre sotto-tipi:
il muscolo striato (o scheletrico), il muscolo liscio ed il muscolo
cardiaco.
•
il tessuto nervoso, costituito sia da cellule ricche di prolungamenti
e facilmente eccitabili (i neuroni), capaci di ricevere e ritrasmettere
gli impulsi nervosi, sia da cellule di più varia forma e funzione, le
cellule
della
glia
(o
nevroglia).
Insieme,
queste
cellule
costituiscono il cervello ed il sistema nervoso.
Tessuti vegetali
I tessuti vegetali si distinguono in tre tipi fondamentali:
•
il tessuto tegumentale, che ha funzioni di rivestimento e di
protezione.
•
il tessuto parenchimatico, che è il più diffuso e svolge sia funzioni
di sostegno che di nutrimento per l'intera pianta.
•
il tessuto vascolare, che permette il trasporto a breve e a lunga
distanza delle soluzioni e dei nutrienti all'interno della pianta, ed è
costituito da due sottotipi: lo xilema ed il floema.
Organo
L'organo
è
l'unità
definibile
morfologicamente,
costituita
dall'associazione di tessuti che svolgono funzioni tra loro integrate.
L'organo fa parte, ed opera, in un apparato o sistema, che riunisce organi
differenti e che permette lo svolgersi in maniera coordinata del complesso
di funzioni di più organi.
Ad una relativa semplicità organizzativa presentata dagli organi presenti
nelle Piante, fa riscontro una maggiore complessità con differenziazione
di numerosi tipi di organi nel regno animale, l'organizzazione di tessuti in
organi si realizza negli organismi pluricellulari a partire dal phylum dei
Platelminti e raggiunge i maggiori livelli di complessità nei vertebrati
superiori.
L'organizzazione di tessuti in organi inizia nel periodo di sviluppo
embrionale al termine della gastrulazione, e continua attraverso il
processo di organogenesi che consiste nell'organizzazione spaziale, nella
moltiplicazione e nella differenziazione di cellule derivanti dai tre diversi
foglietti embrionali.
Il processo di organogenesi assume caratteri specifici per ciascun organo
ed è caratterizzato dall'integrazione fra differenziamento cellulare ed
interazioni fra popolazioni cellulari. Anomalie di tale processo si possono
tradurre in malformazioni con diversa rilevanza funzionale a seconda
dell'organo coinvolto, sia che tali anomalie interessino le popolazioni di
cellule indifferenziate che in via di differenziazione.
Dal punto di vista strutturale, gli organi si distinguono in organi cavi e
organi solidi. I primi sono costituiti da pareti che racchiudono un lume,
idoneo ad accogliere un contenuto, mentre i secondi mancano di una
cavità principale, con i tessuti organizzati in strutture compatte.
LE LEGGI DI KEPLERO
PRIMA LEGGE
L'orbita descritta da un pianeta è un ellisse, di cui il Sole occupa uno dei
due fuochi.
SECONDA LEGGE
Il raggio vettore che unisce il centro del Sole con il centro del pianeta
descrive aree uguali in tempi uguali.
TERZA LEGGE
I quadrati dei periodi di rivoluzione dei pianeti sono proporzionali ai cubi
dei semiassi maggiori delle loro orbite.