04/05/17
MITOSI E MEIOSI
GAMETOGENESI
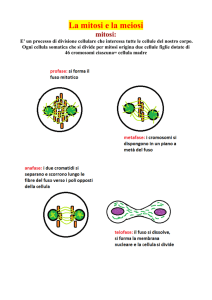
Nella meiosi I
si separano i
cromosomi
omologhi
Nella meiosi II
si separano i
cromatidi
fratelli
Cellula uovo
E l unica cellula animale che ha
tutto l occorrente per generare un
nuovo individuo. Essa é di forma
sferica o ovoidale con un diametro di
100 µm nell uomo, di 1-2 mm nelle
rane e nei pesci, diversi cm negli
uccelli e nei rettili. Una tipica cellula
somatica ha invece un diametro di
circa 20 µm.
La gametogenesi é quel processo per
mezzo del quale, nell embrione in fase di
sviluppo, le cellule germinali capostipiti
migrano nelle gonadi per moltiplicarsi e
diventare gameti maturi: ovuli e
spermatozoi. La fusione della cellula uovo
e dello spermatozoo dopo la fecondazione
e la formazione dello ZIGOTE avvia
l embriogenesi.
STRUTTURA DELLA
CELLULA UOVO
Il citoplasma contiene il vitello o
tuorlo, ricco di sostanze nutritive,
soprattutto lipidi e proteine,
contenute nei granuli vitellini. Una
caratteristica molto importante
della cellula uovo è il rivestimento
esterno che nei non-mammiferi si
chiama membrana vitellina e nei
mammiferi zona pellucida; la
funzione è quella di proteggere
l'uovo da danni meccanici. Alcune
cellule uovo contengono i granuli
corticali che liberano il loro
contenuto durante la fecondazione.
1
04/05/17
OV O G E N E S I
Le cellule germinali capostipiti migrano verso le gonadi per
diventare ovogoni. Dopo alcune divisioni mitotiche diventano oociti
primari. Gli ovociti primari cominciano a formarsi verso il 3° mese
dello sviluppo fetale. Alla nascita essi sono ca. 2 milioni. Queste
cellule subiscono la I divisione meiotica come ovocita primario e
nei mammiferi rimangono nella profase della I divisione meiotica
per diversi anni fino a che non si raggiunge la maturità sessuale.
Alla pubertà gli ovociti primari sono circa 200.000 (da 1-2 milioni
che erano alla nascita) e sotto l'influenza ormonale, maturano uno
ogni 28 giorni (ciclo mestruale) per tutto il periodo che va dalla
pubertà alla menopausa (45-50 anni). Di questi circa 400 arrivano
alla completa maturazione. Il resto degli ovociti continuano a
degenerare per tutta la vita riproduttiva.
L oocita secondario abbandona l ovaio
diretto verso l ovidotto, fermo allo stadio
della metafase II.
Solo in seguito alla (eventuale)
fecondazione si completa la II
divisione meiotica e si forma anche
il II globulo polare.
Questa divisione ineguale ha il vantaggio
di far passare quasi tutto il materiale di
riserva accumulato nell'ovocita alla
cellula uovo matura.
La I divisione meiotica dell'ovocita primario produce
un ovocita secondario e un globulo polare.
Appena un ovocita primario é pronto a completare la
Meiosi si dissolve la membrana nucleare e i
cromosomi si dirigono verso la membrana cellulare.
Il nucleo si divide e il citoplasma forma una
protuberanza. Metà dei cromosomi si dirige verso
questa protuberanza che darà origine al primo
globulo polare e il resto della cellula costituisce
l'ovocita secondario.
La I divisione meiotica si completa poche ore prima
dell'ovulazione (cioé della liberazione dell'ovocita
dalle ovaie).
L ovocita secondario va incontro alla II divisione
meiotica e si ferma in metafase II fino alla
fecondazione.
OVOGENESI
Per ovogenesi s intende lo
sviluppo della cellula-uovo
nell'apparato genitale
femminile, che é costituito, fra
l'altro, dalle ovaie (nelle quali
vengono prodotti gli ovociti) e
dall'utero rivestito
internamente dall'endometrio
nel quale s'impianta l'uovo
fecondato. L ovulo durante lo
sviluppo è detto ovocita. Esso
si trasforma in cellula-uovo,
appunto, nel processo di
ovogenesi che comprende
delle mitosi e la meiosi con la I
divisione meiotica e la II
divisione meiotica.
3°mese
vita fetale
≈ 2.000.000
alla nascita bloccati
in PROFASE1
200.000
Alla pubertà
Poche ore prima
dell ovulazione
Ovocita secondario
rimane in METAFASE II
fino alla fecondazione
Solo 400 arrivano a maturazione in tutta la vita
OOGENESI
1
2
Una cellula somatica (10-20 µm) impiega 24 ore
per crescere e prepararsi alla divisione cellulare.
In proporzione una cellula uovo del diametro di
1000 µm e più dovrebbe impiegare un tempo
molto maggiore di quello effettivamente
impiegato.
Se ciò non avviene é perché altre cellule
cooperano alla crescita e sono le cellule
nutrici (presenti in alcuni invertebrati) e le
cellule follicolari (o cellule della
granulosa). LE CELLULE DELLA GRANULOSA
1
2
PRODUCONO ESTROGENI (estradiolo
principalmente) E UN LIQUIDO che crea intorno
all oocita in maturazione una cavità: ANTRO.
2
04/05/17
In molti vertebrati la maturazione
dell ovocita avviene ad opera di
ormoni liberati dall ipofisi: le
gonadotropine.
Ogni ovocita si sviluppa all'interno del
follicolo primario (ovocita circondato
da un singolo strato di cellule
follicolari) I follicoli primari si trovano
vicino alla superficie dell'ovaio e
possono crescere e trasformarsi in
follicoli in via di sviluppo fino a
diventare i cosiddetti follicoli antrali,
sotto l azione di determinati ormoni
(soprattutto l ormone stimolante il
follicolo, FSH ed estrogeni).
La zona pellucida (membrana di natura glicoproteica)
separa l oocita
in sviluppo dalle cellule follicolari
I follicoli in via di sviluppo continuando a crescere o
degenerano o liberano l ovocita secondario con
l ovulazione.
Una volta al mese, verso la metà del ciclo mestruale, viene
secreto l ormone luteinizzante (LH) che accelera la
crescita di 15-20 follicoli e di questi uno solo maturerà e
libererà l ovocita secondario che potrà essere fecondato
mentre gli altri regrediscono.
Dopo che l'ovocita secondario é stato liberato dal
follicolo (ovulazione), le restanti cellule del follicolo
formano il corpo luteo che secerne progesterone ed
altri ormoni.
Se la cellula uovo matura non é fecondata il corpo
luteo viene riassorbito in circa 2 settimane. Se invece
la cellula uovo viene fecondata il corpo luteo rimane
attivo (continua cioé a produrre progesterone ed altri
ormoni) per circa 3 mesi.
Abbandonato il follicolo l'ovocita inizia il suo viaggio
nell'ovidotto e ciò avviene una volta al mese.
Nell'ovidotto l'ovocita rimane circa 3 giorni e la sua
capacità di essere fecondato nell'ovidotto é di circa 36
ore. Se viene fecondato esso s'impianta
nell'endometrio 3-4 giorni dopo aver raggiunto l'utero
e 6-7 giorni dopo essere stato fecondato.
Il percorso compiuto dall ovocita
nell ovidotto è illustrato nella seguente
figura:
60h
36h
6g dopo la
fecondazione
3
04/05/17
La fecondazione comporta:
· una modificazione della superficie della
membrana esterna per impedire l'ingresso di altri
spermatozoi;
· l'attivazione metabolica dell'uovo;
·
l'introduzione del materiale genetico dello
spermatozoo;
· la segmentazione.
Se la cellula uovo non viene fecondata
muore e viene espulsa insieme con il
rivestimento dell'endometrio con le
mestruazioni.
Dopo 36 ore dalla fecondazione, l'uovo si divide in
due
- dopo 60 ore in quattro
- dopo tre giorni in otto cellule e così via e intanto
continua ad avanzare sempre nell'ovidotto.
SPERMATOGENESI
Lo spermatozoo é una cellula molto
allungata costituita da
una testa,
una regione intermedia e una coda.
Il DNA del nucleo é fortemente addensato,
inattivo e privo di istoni sostituiti da altre
proteine molto più basiche (ricche soprattutto di
arginina) dette protammine.
TESTA
Quasi tutta la sua massa é occupata dal
nucleo aploide e una parte abbastanza
limitata dalla vescicola acrosomiale, ricca di
enzimi idrolitici (tra cui la ialuronidasi e
l'acrosina).
Il segmento intermedio é costituito da una
guaina di mitocondri fortemente modificati
(ad es. negli spermi di lepidotteri é difficile
identificarli) e da nessuno, uno o due
centrioli, da uno solo dei quali, chiamato
corpo basale, si originano i microtubuli della
coda.
Nella coda i microtubuli assumono il tipico
sistema 9 + 2 dei flagelli (2 singoli al centro e
9 doppi in circolo).
4
04/05/17
La spermatogenesi o spermiogenesi
avviene nell'apparato riproduttore maschile
(testicoli).
I testicoli sono costituiti da un gran numero di
tubuli seminiferi
dove avviene la
spermatogenesi.
Gli spermatozoi derivano da cellule
g ermin ali immatu re c h i a m a t e
spermatogoni che si trovano nel
perimetro esterno dei tubuli
seminiferi.
Alcuni di essi si differenziano in
spermatociti primari che entrano
nella prima profase meiotica dove i
cromosomi appaiati subiscono il
crossing-over e vanno poi alla I
divisione meiotica per dare due
spermatociti secondari contenente
ciascuno 22 cromosomi più un
cromosoma X o Y (con 2 cromatidi
fratelli ognuno prima della II
divisione meiotica). Alla II divisione
meiotica ognuno dà
luogo a
quattro spermatidi con un corredo
cromosomico aploide. Essi passano
nell epididimo e maturano
ulteriormente.
SPERMATOGENESI
2
1
1
2
Differenze meiosi tra
spermatogenesi e ovogenesi
I
risultati sono molto diversi
perché mentre nel maschio da ogni
spermatocita primario si formano
quattro spermatidi che si
differenzieranno a spermatozoi,
nella femmina la meiosi porterà
alla formazione di una sola cellula
uovo e di due-tre globuli polari
(che degenerano).
5
04/05/17
Determinazione del sesso
Determinazione del sesso
XXoXY
CROMOSOMAX:grande,
con@eneil6%delDNAtotale.300geni
CROMOSOMAY:piùpiccolo,dimensionivariabilia
secondadellaquan@tà[email protected]
• Ilsessomaschileèdeterminatodallapresenza
delcromosomaY
temperature-dependentsexdetermina8on(TSD),andgene8csexdetermina8on
(GSD)
Il Cromosoma X
IlcromosomaXcon@enecen@naiadigenichenon
hannoachefaredireNamenteconilsesso.
L'ereditàdiques@geniseguespecificheregole,
InfaR:
• ImaschihannoununicocromosomaX
• QuasituRigenidell Xnonhannoungene
corrispondentenell Y
• TuRigenisull’X,ancheserecessivinelle
femmine,sarannoespressineimaschi.
Mammiferi:
Inattivazione Cromosoma X
Nel 1961 Mary Lyon propose che nei
mammiferi, la dose di prodotti genici sia
identica nei maschi e nelle femmine
inattivando casualmente uno dei due
cromosomi X nelle femmine. Il cromosoma X
inattivo è il corpo di Barr.
Questo meccanismo di compensazione
genica è spesso chiamato l’ipotesi di Mary
Lyon.
Mammiferi:
Inattivazione Cromosoma X
• Sisonoevolu@meccanismipercompensareladifferenza
didosaggiogenicodelcromosomaX,presentein2copie
nellefemmineein1copianeimaschi
Imammiferirealizzanolacompensazionedidosedel
cromosomaaNraversoilmeccanismodiinaRvazionedell X,
chiamatoanchelyonizazzione.
MaryLyonèstatalaprimaasuggerirequestomeccanismo
Mammiferi:
Inattivazione Cromosoma X
Il numero di corpi di Barr varia con il
numero di cromosomi X
XX
Un corpo
di Barr
XXX
Due corpi
di Barr
6
04/05/17
Mammiferi:
Inattivazione Cromosoma X
2 cromosomi X nelle donne,
1 solo negli uomini?
LeregioniPARpresen8suicromosomisessuali
contengonogenichenonsonoinaGva8,
perchéildoppiodosaggioèassicurato
comunque
1. ilcromosomaX
raddoppial’espressione
dituRigenicontenu@,
cioèsiproduce2volte
piùRNA
2. nellefemmineunodei
duecromosomiXè
inaRvatocasualmentein
ciascunacellulaallo
stadiodiblastocis@
PAR1 ha 24 geni, PAR2 ha solo 4 geni
ERRORI DELLA MEIOSI
trisomia 21 Down
Il fenomeno della non-disgiunzione
porta a gameti con un cromosoma in
più (es. sindrome di Down)
Incidenza: circa 1/1000 nati vivi
anomaliecromosomicheriscontrate
13
12
11.2
p
11.1
11.2
2
altre t
3
t21;22
21
17
t21;21
5
t15;21
q
6
t13;21
22.1
15
t14;21
925
free T21
1000
100
10
1
Numero
DSCR
Anomalia
11.1
27
mosaicism
D21S17
22.2
ETS2
22.3
MX1
HC21
7
04/05/17
Trisomia 21 (Sindrome di Down)
40.000 casi in Italia
◆
Neurologici:
– Ritardomentale
100%
– Alzheimerdopoi35anni 100%
◆
Ipotoniamuscolare
◆
Bassastatura
100%
70%
Sindrome di Klinefelter (47,XXY)
1:900-1:600 maschi
•
•
•
•
Il50%dellegravidanzegiungeatermine
Feno@pomaschile
CaraNeris@cheprincipali:
Staturaalta
– Ipogonadismo,bassilivelliditestosterone,
mancataproduzionedispermatozoi(azoospermia)
equindisterilità
– Ginecomas@a
– Sial intelligenzasial aNesadivitasonoquasi
normali
LeregioniPARpresen8suicromosomisessuali
contengonogenichenonsonoinaGva8,
perchéildoppiodosaggioèassicurato
comunque
• Sindrome di Klinefelter (47,XXY)
Il Klinefelter?
1:900-1:600 maschi
1. NelKlinefelter(XXY)uno
deiduecromosomiXè
inaRvatocasualmentein
ciascunacellulaallo
stadiodiblastocis@
2. Quindiildosaggio
sarebbemantenuto
IlgeneSHOX
ShortstatureHOmeoboX-containing
• MutazioniodelezionidelgeneSHOXnella
regionePAR1causaritardodicrescitaebassa
statura.
• Labassastaturadidonneconsindromedi
TurnerSyndrome(X0)èilrisultatodiunasola
copiadiSHOX(maancheilquartometacarpo
corto)
• LamaggiorestaturanelKlinefelter(XXY)e
nellatriplaX(XXX)potrebbeessereilrisultato
di3copiediSHOX
PAR1 ha 24 geni, PAR2 ha solo 4 geni
8
04/05/17
Trisomia X (47,XXX)
1:1.200
• Il70%dellegravidanzegiungeatermine
• Errorenelladisgiunzionematernae
correlaconl etàmaterna
• CaraNeris@cheprincipali:
– Staturaalta
– Fer@litànormale,irregolaritàciclo
– Sial intelligenzasial aNesadivitasono
normali
Maschio (47,XYY)
1:1.000 maschi
• Feno@pomaschile
• CaraNeris@cheprincipali:
– Staturaalta
– Fer@litànormale
– Nonviècorrelazioneconl etàpaterna
– Sial intelligenzasial aNesadivitasono
perfeNamentenormali
MonosomiaX(45,X0)Turner
MonosomiaX(45,X0)Turner
Èl unicamonosomiacompa@bileconlavita,mail
98%dituRife@monosomiciTSvaincontroad
abortospontaneo
Prendeilnomedall endocrinologoHenryTurnerche
ladescrissenel1938
L incidenzanegliabor@ècircail7-10%,mentrealla
nascitaè1/2500femmine.
LasindromediTurner(TS)definisceuncomplesso
feno@poumanofemminile,dovutoacompletao
parzialeassenzadelsecondocromosomasessuale
Nonèchiaroperchéilcario@po45,X0sialetalein
uteroedinvececompa@bileconlasopravvivenza
postnatale
Dipendedaunerrorenellaspermatogenesinell 80%
deicasienoncorrelaconl etàdeigenitori
LaveramonosomiadelcromosomaXèresponsabile
del45%deicasiTS;glialtrihannomosaicismo(45,
X0/46,XX)e/ounanormalecromosomaXoY
UnprecedentefiglioconTSnonaumentailrischio
riproduRvoprevistoperunacoppiadiparietà
Unbassolivellodimosaicismosoma@coTurner,
inferioreal2%,èdinormaleriscontronella
popolazione
triploidia
MonosomiaX(45,X0)Turner
lamenopausaprecedeilmenarca Leovaiesonoallungateeformatedatessuto
stromaleprivodifollicoli:gliooci@sonospesso
anda@inapoptosiprimadei2annidivita
L insufficienzaovaricaprepuberaleportaad
amenorreaprimaria,sterilitàecarenzadi
estrogeni
Inmenodel10%deicasi,lapubertàpuò
verificarsiesonopossibiligravidanzeconun
aumentatorischiodiperditafetale
Ancheinrapportoall eterogeneitàdel
geno@po,ilfeno@posimanifestainmodo
moltovariabile
Frequenzaallanascita=1/10.000
Frequenzanegliabor7=1/14
Cario@po69,XXY57%
Cario@po69,XXX40%
Cario@po69,XYY3%
9
04/05/17
Mosaicismo, quando il cambiamento
avviene dopo che si è formato lo
zigote
47,XXY/46,XY
IDIVERSITIPIDIMUTAZIONI
Mutazioni geniche (o
puntiformi)
MutazioninellastruTuradei
cromosomi
Esistono 4 tipi principali di mutazione della
struttura dei cromosomi
• delezione
Mutazioni cromosomiche
§ Mutazioni nel numero dei cromosomi
• duplicazione
§ Mutazioni nella struttura
• inversione
• traslocazione
Delezione.
Parte del cromosoma (e
quindi delle basi del DNA) viene perduta.
Tutti i geni contenuti nella parte
interessata dalla delezione vanno
perduti e quindi non possono esprimersi nel
fenotipo
1)
2)
abcdefghilmnopqrstuvz
abcdefghilmnopqrsvz
tu
10
04/05/17
1)
abcdefghilmnopqrstuvz
2)
abcdefgefghilmnopqrstuvz
Inversione.
In seguito ad una rottura, una
parte del cromosoma viene riunita in posizione invertita.
A parte i geni che si possono trovare nel punto di
rottura, non ci sono gravi conseguenze per la cellula.
Tuttavia non può avvenire crossing over in questa
regione in quanto essa non si appaia con quella del
cromosoma omologo
1)
abcdefghilmnopqrstuvz
2)
abcdefghilmno
3)
abcdefghilmnorqpstuvz
stuvz
r
Parte del cromosoma
viene duplicata. I geni contenuti nella parte
interessata sono duplicati. Perciò la cellula
può contenere tre o quattro copie di ciascun
gene. In qualche caso ciò può danneggiare
la cellula in quanto i sistemi di
regolazione possono essere alterati
pq
Duplicazione.
Leucemia mieloide cronica
Traslocazione.
In seguito ad una rottura, una
parte del cromosoma (es n° 6) si stacca e viene
inserito in un cromosoma diverso (es n° 12). Le
t ra s l o c a z i o n i p o s s o n o r e n d e r e c o m p l i c a t o
l appaiamento dei cromosomi durante la meiosi
abcdefghilmnopqrstuvz
1)
2)
ABCDEFGHILMNOPQRSTUVZ
abcdefghilmnopqrs
A B C D E F G H I L M N O P Q R S T U V Z tuvz
Presenza di
cromosoma anomalo,
il cosiddetto
un
cromosoma Filadelfia
o cromosoma Ph1,
o r i g i n a d a l l a
traslocazione reciproca i
cromosomi 9 e 22. Sul
cromosoma 9 si trova un
oncogene abl, che svolge
ruolo importante nella
crescita delle cellule
normali.
Leucemia mieloide cronica
il gene abl viene rotto in due
pezzi:
• uno rimane sul cromosoma 9
• l'altro si trasferisce sul
cromosoma 22 e si fonde con un
frammento di un altro gene
chiamato bcr
In conseguenza di questo scambio
genetico si viene a creare un
nuovo gene, che prima non
esisteva, chiamato bcr/abl.
Il nuovo gene di fusione bcr/abl
produce una proteina anomala,
molto più potente nello stimolare
la crescita delle cellule. Single Nucleotide Polimorfism (SNP)
bp alternativi presenti ad alta frequenza
nella popolazione (non meno dell 1%)
• Diversità genetica tra gli individui
• Interne a regioni non codificanti o a geni
• Suscettibilità a malattie
• Terapia farmacologica personalizzata
(farmacogenetica)
11
04/05/17
Traslocazioni robertsoniane (rob)
rob
• coinvolgonoicromosomiacrocentrici13,14,15,
21e22
• nessunaregionecromosomicaèassente,perché
ques@contengonounbracciocortoprivodigeni
chepuòrisultareperdutoconlafusionedei
bracciqdiduecromosomiacrocentrici
• LapiùfrequentetraslocazioneRobertsonianaèla
rob(13q14q)cherappresentail75%dituNelerob
• seguepoilarob(14q21q)elarob(21q21q)
• siformanoingeneredurantelameiosifemminile
ecomportanoinfer@litàmaschileoabor@vità
ripetuta.
Inversioni
• Leinversionisonorare(menodi1casosu1000)eavolte
difficilidameNereinevidenza
• Possonoesseresempliciquandocomprendonoduepun@di
roNurasudiunsingolocromosoma
• Sonopericentrichequandoilsegmentoinver@tocon@eneil
centromero(es:46,XXinv(3)p25q21)
• Leinversionipericentrichedeicromosomi1,9,16eYsono
eteromorfismicitogene@cidinormaleriscontroinsoggeR
sani
• LeinversionisonodeNeparacentricheseconfinateadunodei
duebracci(es:46,XX.Inv(11)q21q23)
• L eterozigoteperun inversioneèunsoggeNonormale.
Percentualeallanascitadifiglicon
cario8posbilanciatodagenitoricon
traslocazionerobertsoniana
•
•
•
•
t(13;14)M=F1%
t(14;21)F15%M2%
t(21;22)F10%M5%
t(21;21)M=F100%
principalisindromidadelezione
LOCALIZZAZIONE
CROMOSOMICA
SOGGETTI CON
MICRODELEZIONE
Prader Willi/Angelman
15q11.13
70%
Williams
7q11.23
90%
DiGeorge/Velocardiofacciale
22q11.2
75%
Smith-Magenis
17p11.2
95%
Miller-Dieker
17p13.3
90%
SINDROME
L IMPRINTING DURANTE LA GAMETOGENESI
La metiltrasferasi de novo
rinnova l imprinting ad ogni
generazione : quindi, durante
la gametogenesi, l'imprinting
viene cancellato e ripristinato
successivamente in base al
sesso del soggetto
La mancanza di un imprinting genetico corretto che
coinvolge i geni del cromosoma 15 causa
Il gene segue un imprinting paterno
Femmina
entrambi i geni che trasmette
alla progenie NON saranno
imprinted (entrambe le copie
saranno attive)
Maschio
saranno imprinted entrambe le
copie (entrambe NON funzionanti)
SINDROME DI
ANGELMAN
SINDROME DI
PRADER-WILLI
Due sindromi complesse che influenzano lo stato ormonale, il
metabolismo e la capacità di movimento.
12
04/05/17
LA GENETICA DELLE MALATTIE DI
E DI
Le due malattie sono di
solito causate da una
microdelezione che
colpisce il braccio
lungo del cromosoma
15 ma, mentre nella
PWS il cromosoma
colpito è quello di
origine paterna, nella
AS è quello di origine
materna. Il fatto che le
due malattie siano
clinicamente molto
diverse è imputabile al
fatto che i geni
presenti in quella stessa regione genomica sono espressi diversamente nei cromosomi ereditati dall uno o dall altro genitore.
Mentre la AS è causata dalla mancata espressione del solo gene UBE3A, la PWS è
causata dalla mancata espressione di più geni.
Delezioni di certe regioni cromosomiche causano un fenotipo
differente se presenti sul cromosoma paterno o materno.
Sindrome di
Angelman
Cromosoma
materno
Cromosoma
paterno
Sindrome di
Prader-Willi
DiGeorge/velocardiofacciale
LasindromediDiGeorgedel22q11.2èlapiùfrequentesindromeda
microdelezione,conunincidenzadi1su4000—5000na@
Ladelezionecomprende3Mbedalmeno30geni
FECONDAZIONE
La fecondazione é la fusione del gamete maschile con
quello femminile ed avviene normalmente tra organismi
dellastessaspecie.
La fecondazione tra specie diverse, laddove é possibile,
portaallaformazionediunindividuoibridocheésterile
perchéicromosomisessualinonsiappaionoallaMeiosi.
TuNavia, quando un individuo sterile raddoppia il
contenuto dei cromosomi può generare un individuo
fer@le perché i cromosomi sessuali si appaiano alla
Meiosi;quest'ul@moéuncasodipoliploidia.
FECONDAZIONE
N e g l i a n fi b i e n e i p e s c i l a
FECONDAZIONE é ESTERNA, mentre
n e i r e R l i , n e g l i u c c e l l i , n e i
mammiferi monotremi, marsupiali e
placenta@éINTERNA.
Con la fecondazione il nucleo della
c e l l u l a u o v o e q u e l l o d e l l o
spermatozoosifondonoperformare
ilgenomadiunnuovoorganismo.
13
04/05/17
Iprincipalistudisullafecondazionesono
sta@ condoR sul riccio di mare la cui
femmina possiede 107 cellule uovo e il
maschio 1012 spermatozoi. Inoltre ha
una fecondazione esterna, più facile da
studiare.
Nella maggior parte degli animali la
fecondazionecomprendevaripassaggi:
·l'avvicinamentodellospermatozooallacellulauovo;
· illegamedellospermatozooallamembranadellacellula
uovo;
· lapenetrazionedeirives@men@dell'uovodapartedello
spermatozoo;
·lafusionedellamembranadellospermatozooconquello
dell'uovo e l iniezione del materiale del nucleo dello
spermatozoonelcitoplasmadell'uovo;
· l'aRvazione del citoplasma dell'uovo con rilascio dei
granulicor@caliebloccodell'entratadialtrispermatozoi;
· la trasformazione del nucleo dello spermatozoo in
pronucleomaschile;
·lafusionedelpronucleomaschileconquellofemminile.
FECONDAZIONE-RICONOSCIMENTO
ADISTANZA
CHEMIOTASSI
Avvicinamentoallacellulauovoedaumentodellamo@lità
dellospermatozoo
MECCANISMI
•Interazioneligando-receNore
•Trasduzionedelsegnale
•ARvazionedineina
•Aumentometabolismoossida@vo
La reazione acrosomiale é scatenata dal
legame di una proteina dello spermatozoo ad
un polisaccaride del rivestimento gelatinoso.
Ne consegue un ingresso di Ca++ e
l innalzamento del pH nella testa dello
spermatozoo. Ciò induce una liberazione del
contenuto della vescicola acrosomiale e la
formazione del processo acrosomiale.
Lafecondazioneinizianelmomentoincuila
testa dello spermatozoo viene in contaNo
con il rives@mento gela@noso della cellula
uovoeiniziacosìlareazioneacrosomialecon
liberazione degli enzimi idroli@ci e di altre
proteine, per roNura della vescicola
acrosomiale, seguita da un intensa
polimerizzazione di ac@na (processo
acrosomiale, non presente nei mammiferi);
gli enzimi idroli@ci perforano la membrana
vitellina consentendo al nucleo dello
spermatozoodientrarenellacellulauovo.
Riconoscimentospecifico
spermatozoo-polisaccaride
delrives@mentogela@noso
QuindiingressoCa++nellospermatozoo,
Liberazionecontenutovescicolaacrosomiale
epolimerizzazioneac@na
14
04/05/17
Lo spermatozoo, una volta
superato il rives@mento
g e l a @ n o s o p u ò l e g a r s i
all'involucro vitellino (o alla
zona pellucida) perché una
sua proteina la bindina si
lega ai receNori della
bindina sulla membrana
vitellina dell'uovo. Tale
legame é streNamente
specie-specifico.
Se la fusione della cellula uovo avviene con
piùdiunospermatozoosiparladipolispermia.
Una volta che il nucleo di un solo
spermatozoostapenetrandoall interno,la
cellulauovoaRvailproprioprogrammadi
sviluppo e diventa impenetrabile agli altri
spermatozoievitandola polispermia.
L ATTIVAZIONE DELLA CELLULA UOVO è dovuta alle
seguen@variazioniioniche:
1. aumento della permeabilità al Na+ che provoca
una rapida depolarizzazione della membrana
plasma@cadell'uovo;
2. liberazione del Ca++ intracellulare sequestrato nel
citosol;
3.entratadiNa+eduscitadiH+coninnalzamentodel
pHintracellulare.
ATTIVAZIONEDELL UOVO
•AumentoaRvitàmetaboliche(sintesi
proteica,metabolismoossida@vo)
•Cambiamen@struNurali
•Completamentodellameiosi
LA DEPOLARIZZAZIONE DELLA
MEMBRANA PLASMATICA dell'uovo
probabilmente induce una variazione
conformazionale e INATTIVAZIONE
DI PROTEINE RECETTRICI PER LO
SPERMATOZOO.
Tale reazione é definita BLOCCO
RAPIDO CONTRO LA POLISPERMIA .
15
04/05/17
Controlapolispermiaagisceneimammiferiunsecondomeccanismo
definito blocco lento contro la polispermia o reazione
cor@cale.
Gli enzimi dei granuli cor@cali libera@ tra la membrana plasma@ca e
quella vitellina a causa dell aumento della concentrazione citosolica
diCa++causano:
1) indurimento della membrana vitellina (deNa ora membrana di
fecondazione),allaqualeglispermatozoinonpossonolegarsi;
2)RimozionedeireceNoriperlabindinaedelleproteinecheuniscono
lamembranaplasma@caaquellavitellina;
3)Aumentodisolu@tramembranaplasma@caevitellina,vienequindi
richiamata acqua che allontana ulteriormente la membrana vitellina
daquellaplasma@ca.
In alcuni animali comprenden@ reRli, uccelli e inseR, il
blocco della polispermia non é così efficace come nel
ricciodimare,anzinellacellulauovopenetranoinuclei
didiversispermatozoi,peròunosolodiessisifondecon
ilnucleodellacellulauovo. Nei mammiferi il meccanismo che
impediscelapolispermiaépiùcomplicato
eancoraingranpartesconosciuto.
Ilnucleodellospermatozooinglobatodall'uovo
perde l'involucro nucleare cambiando la sua
struNura e trasformandosi in pronucleo
maschile(nucleoaploide).
PRONUCLEO MASCHILE:
•Dissoluzionedell involucronucleare
•Sos@tuzioneproteineistoniche(protammine)
conquelledell uovo(istoni)
•Riarrangiamentodellacroma@nainformapiù
lassa
•Ricos@tuzionedellamembrananucleare
Il nucleo della cellula uovo forma il pronucleo
femminile terminando la meiosi II con la
liberazione del II globulo polare
I due pronuclei compiono un movimento di
avvicinamento.
Nel riccio di mare essi si fondono completamente
a formare lo ZIGOTE prima della divisione Mitotica
che segue la fecondazione.
16
04/05/17
Durantelafecondazione,sideterminaancheilsesso
dell organismochesaràgenerato.
Glispermatozoidonanoallacellulauovoancheidue
centrioli. Dai centrioli si assembla una struNura a
raggiera di microtubuli (l aster sperma@co) che
guida l avvicinamento e la fusione dei due
pronuclei.
NEI MAMMIFERI
3) Lo spermatozoo si lega ad una glicoproteina
(chiamata nel topo ZP3) della zona pellucida
(nonmembranavitellina);èquestolegameche
determinalafecondazionespecie-specifica
4) Le molecole di riconoscimento sono situate
sulla membrana plasma@ca dello spermatozoo
enonsuquellaacrosomiale
Mentre il nucleo sperma@co si trasforma in pronucleo maschile Nei
mammifericisonoalcunedifferenze:
1) Nonabbiamounacellulauovomaunovocitasecondariochecompleta
lameiosisoloinseguitoallafecondazioneliberandounglobulopolare
2) Glispermatozoinonpossonofecondarel ovocitaseprimanonhanno
subito un processo di maturazione deNo CAPACITAZIONE a carico di
secrezionipresen@nell apparatogenitalefemminile
CAPACITAZIONE(mammiferi)
•Aumentodellamo@litàdellospermatozoo(rimozionedelcolesterolodalla
membranaplasma@ca)
•Acquisizionedellacapacitàdiriconoscerel uovoedieffeNuarelareazione
acrosomiale(esposizionedireceNorichericonosconospecificheproteinenella
zonapellucidadell uovo)
MECCANISMI
Ricezionediunsegnale
Modificazioninellacomposizionedellamembranaplasma@ca
5) In seguito al legame vi è una reazione acrosomiale
ma non un processo acrosomiale
6) Dopo il nucleo anche il resto dello spermatozoo (ad
eccezione della coda) entra nell'uovo il cui citoplasma
ingloba gli organelli citoplasmatici dello spermio.
7) Non vi è un blocco rapido contro la polispermia ( a
causa del numero ridotto di spermatozoi che arriva
all uovo, circa 200) ma solo uno lento (reazione
corticale con disattivazione di ZP3)
8) I centrioli sono donati dalla cellula uovo e non dallo
spermatozoo
9) I due pronuclei avvicinatisi non si fondono finché le
membrane nucleari non si sono dissolte; i pronuclei
entrano in mitosi e si fondono durante la metafase
preparandosi alla prima divisione cellulare dello zigote.
17