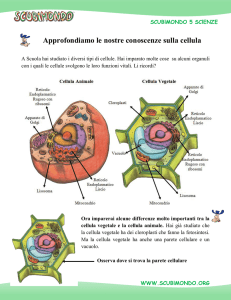
LA PARETE CELLULARE
In tutte le cellule vegetali esiste esternamente alla membrana plasmatica un robusto
rivestimento detto parete cellulare.
Tale struttura presenta diverse funzioni:
- funzione protettiva;
- controbilancia fisicamente la pressione osmotica del succo cellulare e assicura
conveniente solidità;
- è interessata per diversi aspetti al movimento di acqua e soluti nella pianta; il flusso
interno ed esterno di acqua e ioni passa attraverso di essa, e quando l’acqua viene
eliminata dalla pianta attraverso l’evapotraspirazione, essa evapora direttamente dalla
parete della cellula.
•Distingue le cellule vegetali da quelle animali ed è responsabile di molte
caratteristiche degli organismi vegetali.
•È rigida, limita perciò le dimensioni della cellula e impedisce la sua rottura
quando aumenta di volume, in seguito all’assorbimento di acqua da parte
del vacuolo (pressione di parete).
•Determina le dimensioni e la forma della cellula, la trama del tessuto e la
struttura degli organi.
•Difende la cellula da batteri e funghi patogeni con la produzione di
fitoalessine, antibiotici tossici per i patogeni o con la sintesi e la
deposizione della lignina che agisce da barriera contro l’invasione.
•Una volta veniva considerata come una struttura inattiva, mentre oggi è
dimostrato che svolge funzioni specifiche ed essenziali. Può contenere una
grande varietà di enzimi e svolgere un ruolo importante nell’assorbimento,
nel trasporto, nella secrezione di sostanze e nel riconoscimento di segnali
esterni (presenta recettori).
La parete cellulare presenta una struttura complessa: dall’esterno lamella mediana,
parete primaria e parete secondaria. La sua costruzione intorno al protoplasto è
progressiva e può essere scandita in diverse fasi.
A - cellule meristematiche: successivamente alla divisione di una cellula meristematica
si formano due cellule figlie, e questo avviene mediante la formazione di un setto detto
setto di separazione. La porzione centrale del setto di separazione costituisce la
lamella mediana.
B - Fase giovanile di accrescimento: in questa fase la cellula costituisce a ridosso
della lamella mediana con le sue forze una sottile parete pectocellulosica alla quale si
da il nome di parete primaria. In questa fase poi la cellula subisce normalmente
notevoli modificazioni: si accresce per distensione. Qui la parete primaria aumenta
proporzionalmente la sua superficie, mantenendo costante il suo spessore.
C - Inizio della fase adulta: una volta terminato l’accrescimento per distensione la
cellula provvede ad irrobustire la sua parete esterna; si forma così uno strato interno alla
parete primaria che va a costituire la parete secondaria.
da: “Biologia” (N.A. Campbell, J.B. Reece)
Le pareti delle cellule vegetali hanno spessore variabile a seconda
della funzione e dell’età. I primi strati costituiscono la parete
primaria sottile ed elastica (1-3 μm), a cui si aggiungono verso
l’interno altri strati che costituiscono la parete secondaria, più
rigida e spessa (5-10 μm). La lamella mediana è lo strato ricco di
pectine che cementa le pareti primarie di cellule adiacenti.
Come abbiamo già accennato e come vedremo, la parete cellulare presenta uno
spessore ed una qualità differenti a seconda dei tessuti di cui la cellula farà parte.
Nelle cellule che presentano il maggior ispessimento (che si concentra maggiormente
nella parete secondaria) a volte si viene a formare anche una parete terziaria.
Lamella mediana
Non supera mai 1/10 di micron, ed è sempre interposta tra le pareti primarie di due
cellule adiacenti. Essa si forma all’atto della separazione di due cellule figlie, nei tessuti
meristematici primari e secondari.
Successivamente alla formazione del fuso mitotico, la cellula vegetale presenta alcuni
caratteri nel meccanismo di divisione che la differenzia dalla cellula animale.
Qui il primo atto della citodieresi è rappresentato dalla formazione del fragmoplasto,
l’apparato con cui prende avvio la formazione del setto di separazione e quindi della
parete cellulare. Tale formazione si rende visibile solo alla fine dell’anafase (profase,
metafase, anafase e telofase). Il fragmoplasto compare come una zona più densa di
citoplasma ricca di fibrille (microtubuli). Mano a mano che si forma il setto, (che procede
dal centro verso le zone periferiche, il fragmoplasto scompare.
In realtà il fragmoplasto sembra essere l’edificio di orientamento per la formazione del
setto di separazione, in quanto alla formazione reale di esso interviene tutta la cellula
con il suo metabolismo interno (Apparato del Golgi e reticolo endoplasmatico).
I dittiosomi dell’Apparato del Golgi producono infatti un’enorme quantità di vescicole che
si portano nella zona dove si formerà il setto. Qui il loro contenuto (proteine e
polisaccaridi non cellulosici) si fondono a costituire l’impalcature del setto di
separazione.
Il setto di separazione sin dai
primi
stadi
della
sua
formazione
si
rivela
birifrangente:
sono
infatti
visibili esternamente due strati
brillanti, separati al centro da
uno strato isotropo. Tale zona
centrale costituisce la lamella
mediana, mentre i due strati
esterni rappresentano le zone
che
in
seguito
si
completeranno come le pareti
primarie delle due cellule figlie.
La lamella mediana ha la funzione di saldare assieme due cellule figlie: essa è infatti
costituita da sostanze pectiche, a forte azione cementante. Si usano infatti particolari
enzimi per poter separare due cellule (pectinasi)
Il setto oltre a separare due cellule deve anche provvedere ad una continuità
citoplasmatica, in modo da permettere degli scambi di metabolici. Ciò avviene attraverso
la formazione di cordoni o canali di connessione detti plasmodesmi.
Numerose sono le teorie rispetto alla formazione dei plasmodesmi. Si pensa ad
esempio a frammenti del reticolo endoplasmatico che rimangono intrappolati nella
costituenda parete cellulare. Sono internamente rivestiti da una membrana lipoproteica
in continuità con quella plasmatica delle due cellule fra loro connesse. Nella sua parte
centrale è presente un microtubulo, molto simile a quelli citoplasmatici detto
desmotubulo.
Su 100 micron quadri,
si possono trovare da
100
a
5000
plasmodesmi. Quando
la
cellula
effettua
l’accrescimento
per
distensione essi non
aumentano di numero,
e quindi diminuisce la
loro densità.
Oltre alle pectine (che sono macromolecole derivate dalla polimerizzazione dell’acido
galatturonico, sovente indicate come acido peptico), la lamella mediana è costituita da
sostanze proteiche: proteine strutturali ed enzimatiche e quindi si hanno in essa attività
metaboliche. E’ assente la cellulosa.
Formano anche la
lamella mediana,
lo
strato
di
materiale
intercellulare
che
tiene unite le pareti
di cellule vegetali
adiacenti.
Le pectine sono polisaccaridi molto idrofili e
grazie all’acqua che essi introducono,
conferiscono alla parete plasticità e flessibilità,
condizione necessaria per la sua distensione.
Parete Primaria
Si forma a ridosso della lamella mediana, e raramente è più spessa di 1 micron. Come
abbiamo visto la prima rudimentale impalcatura della parete primaria si forma
contemporaneamente alla costruzione del setto di separazione (natura tri-stratificata).
Possiamo dire che la lamella mediana e la parete primaria iniziano
contemporaneamente la loro formazione, solo che la parete primaria completa più tardi
la sua architettura, sotto la direzione del protoplasto delle due cellule figlie, mentre la
mediana si può ritenere tutta costruita dall’attività del protoplasto della cellula madre.
La parete primaria è formata da una sostanza fondamentale o matrice, e da una
sostanza fibrillare in essa disperso.
La matrice è costituita da acqua, emicellulose, sotanze pectiche, proteine e lipidi. La
matrice è la parte più abbondante e l’acqua rappresenta il suo 60%, con abbondante
presenza di emicellulose (polisaccaridi a catena ramificata di xilosio o arabinosio, dei
pentosi, e di glucosio ed altri esosi). Sono quindi polisaccaridi complessi non molto simili
alla cellulosa.
Le proteine presenti sono
per lo più proteine
strutturali
(come
glicoproteine),
ovvero
strettamente
connesse
con la crescita per
distensione della cellula
(estensina). Oltre a ciò
sono
anche
presenti
proteine
enzimatiche
(ATPasi,
proteasi,
galtturonidasi).
Il sistema fibrillare è dato dalla cellulosa, legami idrogeno beta 1-4. Nei funghi si ha la
chitina, un polisaccaride contente azoto, mentre nei lieviti torva posto il poliglucano,
che differisce in parte dalla cellulosa.
La cellulosa è la molecola
organica più abbondante sulla Terra),biologica
costituita da monomeri
di β-glucosio uniti
da
più abbondante
nella
legami 1,4, le cui lunghe molecole sono
unite in microfibrille
del diametro
di 10-25dei
nm
biosfera,
costituendo
il 50%
grazie alla formazione di legami idrogeno tra i gruppi –OH (C3-C6) di molecole
composti organici naturali.
parallele. Ogni microfibrilla è costituita da un centinaio di catene di cellulosa.
Il componente principale della parete è un polisaccaride, la cellulosa (la molecola
parallele. Ogni microfibrilla è costituita da un centinaio di catene di cellulosa.
Il grado di polimerizzazione della cellulosa varia da 2000-6000 unità di
β-glucosio nella parete primaria a >13000 unità di β-glucosio nella
parete secondaria.
Le microfibrille si uniscono avvolgendosi tra di loro per
formare le macrofibrille del diametro 0,5 μm. Questa
particolare struttura conferisce alla parete una resistenza pari
ad una lamina di acciaio dello stesso spessore.
Alcune regioni delle microfibrille, le micelle, presentano
proprietà cristalline per la disposizione ordinata delle molecole
di cellulosa.
La cellulosa forma un’impalcatura compenetrata da una
matrice costituita da emicellulose, pectine, proteine
strutturali (glicoproteine) ed enzimi, che si legano ad essa
trasversalmente.
Le pareti cellulari sono state
paragonate a cemento armato;
in esso sono immersi dei
tondini
di
acciaio,
(le
microfibrille di cellulosa), che
servono a rinforzare il cemento
(la matrice).
xiloglucani
Le emicellulose sono polisaccaridi eterogenei (es. xiloglucani
nelle Dicotiledoni e xilani nelle Monocotiledoni) che sono
legati alle microfibrille di cellulosa tramite legami idrogeno
(stabilizzano la parete cellulare limitandone l’estensibilità e
l’aumento di dimensioni della cellula).
Nella parete primaria la cellulosa raggiunge soltanto il 10-15% del peso fresco della
parete e di conseguenza le microfibrille hanno molta occasione di disporsi a caso
nell’abbondante matrice, e quindi all’inizio della formazione della parete primaria
(prima dell’accrescimento per distensione), si parla di due tipologie di tessitura
dispersa:
- Fogliata: quando le microfibrille non solo sono sovrapposte ma anche intrecciate
tra loro; lo si riscontra in cellule isodiametriche che rimangono tali anche dopo la fase di
accrescimento per distensione.
- Tubolare: qui le microfibrille sono disperse orientate però più o meno
perpendicolarmente il futuro asse della cellula; si riscontra in cellule che accrescendosi
assumono forma cilindrica.
Nelle pareti primarie che hanno già subito l’accrescimento per distensione si
riscontra una tessitura di tipo:
- Fibroide: la disposizione delle fibrille segue la direzione dell’asse principale della
cellula; si forma nelle pareti primarie che hanno già subito l’allungamento per
distensione.
Parete secondaria
Si appone a ridosso internamente alla parete primaria. Si forma dopo che la cellula ha
completato l’accrescimento per distensione.
Costituisce la maggior parte di tutta la parete cellulare e può raggiungere uno spessore
di 3-5 micron.
Essa si accresce centripetamente, per la deposizione di strati concentrici; a volte il suo
ispessimento è tale che il lume della cellula è notevolmente ridotto.
Anch’essa è costituita da una matrice e da un sistema fibrillare formato da cellulosa.
Qui la porzione di matrice però è molto scarsa, mentre prevalente è il contributo del
sistema fibrillare. In alcune cellule come le fibre in genere o i pappi ed i peli del cotone
la porzione di cellulosa può arrivare al 95%.
Considerando una tale abbondanza di microfibrille, esse debbono essere ben serrate
tra loro e disposte parallelamente in quanto solo così possono sostenere una così
elevata concentrazione di cellulosa. La deposizione delle fibrille segue una
organizzazione a strati.
L’orientamento rispetto all’asse longitudinale della cellula è piuttosto vario, ma il più delle
volte obliquo.
La tessitura parallela all’asse maggiore è piuttosto rara ed è presente solo su cellule
fibrose, e la tessitura stessa è detta fibrosa.
Più comunemente la disposizione delle fibre è obliqua rispetto all’asse; essa è tipica
delle cellule meccaniche e delle tracheiti, ed è detta tessitura spirale.
In alcuni casi, come nelle tracheiti a ispessimento anulato, la disposizione delle fibrille
è perpendicolare alla direzione dell’asse principale della cellula, la tessitura è detta
anulare.
La parete secondaria, che nei vasi e nelle cellule meccaniche è piuttosto spessa, mostra
sempre una struttura stratificata. Ciascun strato è costituito da più lamelle, e nelle
tracheidi delle conifere e nelle fibre extraxilari si possono evidenziare tre strati: S1, S2,
S3. Lo strato S3 è quello più interno e delimita il lume cellulare.
Lo strato S2 è il più spesso della parete e da solo condiziona quasi la totalità dello
spessore. Nelle sue lamelle le fibrille sono orientate in modo da formare un piccolo
angolo rispetto all’asse longitudinale, e l’orientamento dell’angolo varia
alternativamente nelle successive lamelle. Tale tipologia di struttura irrobustisce
meccanicamente la cellula. Lo strato S3 è di scarsa rilevanza e la disposizione delle
fibrille è tale da creare un grosso angolo rispetto all’asse principale della cellula
Lo strato S3 comunque non sempre esiste (nelle fibre xilari delle Angiosperme infatti non
è presente).
Prima di morire il protoplasto depone un leggero strato verrucoso sulla superficie
interna di S3. Nel legno di reazione, spesso nella porzione interna del lume cellulare si
appone uno strato gelatinoso (sulla superficie interna dell’S3).
Da quanto detto è facile distinguere la parete primaria da quella secondaria? Non
sempre:
- infatti il tipo di tessitura a volte può essere piuttosto simile (anche nella primaria si
possono sviluppare bande a tessitura parallela);
- a volte non è possibile distinguerle nemmeno considerando le due diverse fasi: prima
e dopo l’accrescimento; infatti vi sono casi (cellule del collenchima, fibre
sclerenchimatiche, ecc.) in cui le pareti si accrescono in superficie anche dopo la
deposizione di diversi strati di ispessimento.
La più significativa differenza tra le due pareti sta nel diverso comportamento delle
fibrille durante le fasi di accrescimento: nella parete primaria le fibrille vengono
spostate individualmente le une rispetto alle altre mentre nella parete secondaria le
lamelle fibrillari subiscono delle lacerazioni ma le fibrille all’interno delle lamelle
mantengono sempre il loro orientamento parallelo.
Incrostazioni della parete
Per la presenza di un complicato sistema di interstizi, la parete cellulare si presta ad
essere stabilizzata, una volta terminato il suo accrescimento, attraverso delle
incrostazioni di lignina.
Questo è uno dei processi più importanti nella vita degli organismi vegetali
superiori. La lignina si va depositare sulle maglie del reticolo cellulosico. La lignina è
facilmente colorabile e quindi sono ben evidenziabili anche le pareti di tutte le
cellule lignificate (rosso).
La lignificazione è una modificazione secondaria della parete cellulare, ed il suo
significato funzionale è quello di aumentare la resistenza meccanica delle
cellule. In particolare la lignina aumenta la resistenza alla compressione delle
cellule. La lignina inoltre rende tutti i complessi di polisaccaridi meno attaccabili
dagli agenti patogeni (funghi, e muffe). Mai lignificate sono invece le pareti delle
cellule dei meristemi primari.
LIGNINA
¾Svolge una funzione prevalentemente meccanica o di
sostegno, conferisce rigidità e resistenza alla compressione,
ed è idrofoba all’acqua (si sostituisce all’H2O nella parete).
¾Rende impermeabile la parete cellulare facilitando il
trasporto dell’acqua verso l’alto nelle cellule conduttrici
dello xilema e permettendo a queste di resistere alla
tensione generata dalla corrente di acqua che viene aspirata
fino alle foglie.
¾La “lignina da ferita” che si deposita in risposta agli
attacchi da parte di funghi o alla comparsa di ferite,
protegge la pianta dall’attacco fungino, aumentando la
resistenza delle pareti alla penetrazione del fungo e
proteggendole dall’attività degli enzimi secreti dal fungo.
precursori fenilpropanoidici
da: “Biologia vegetale” (A. Bruni, M. Nicoletti)
La lignina non viene prodotta
all’interno della cellula come
tale
ma
suoi
precursori
sintetizzati
nel
citoplasma,
migrano nella parete ove
polimerizzano. Chimicamente la
lignina è un polimero di
fenilpropanoidi (C6-C3), con
alcuni residui zuccherini, che
può differire in composizione a
seconda
della
specie
considerata.
La lignificazione non è omogenea in tutte le porzioni ed i comparti della parete: a
volte è più abbondante nella lamella mediana e nella parete primaria, in altri casi
negli ispessimenti della secondaria. In generale, siccome la parete primaria è
povera di cellulosa e ricca di matrice, ne deriva che in essa la lignina può
depositarsi in quantità molto elevata.
Altre sostanze impregnanti possono essere fenoli e tannini
(pigmentazione), sostanze minerali (mineralizzazione),
La cutina si appone sulle pareti delle cellule epidermiche
(cutinizzazione) e la suberina in quelle del sughero.
Cutina, suberina e cere sono sostanze grasse che si
trovano nelle pareti delle cellule dei tessuti esterni di
protezione della pianta (foglie, fusti, frutti ecc.) dove
formano vere e proprie barriere capaci di impedire la
perdita di acqua dalla superficie della pianta oppure
fungere da protezione contro agenti patogeni e agenti
atmosferici.
la cutina si trova nelle pareti (lato esterno)
dell’epidermide delle foglie e dei fusti giovani
dove insieme alle cere forma la cuticola.
da: “Biologia delle piante” (P.H. Raven et alii)
la suberina si trova insieme
delle cellule del sughero, un
protezione (periderma),
nell’esoderma.
da: “Biologia delle piante” (P.H. Raven et alii)
alle cere nelle pareti
tessuto secondario di
nell’endoderma e
GOMME E MUCILLAGINI
Le pareti cellulari possono gelificare a seguito
della costituzione di cellule specializzate oppure
per reazione a ferite. L’elevato contenuto in
sostanze pectiche di alcune pareti cellulari viene
utilizzato per fare marmellate o per estrarre
pectina di impiego alimentare. Alcune pareti
cellulari possono essere convertite in gomme e
mucillagini con un processo indotto da stress
(es. incisione del fusto con un coltello); le
gomme mucillaginose che fuoriescono vengono
raccolte ed utilizzate in farmacia.
Punteggiature
Per mantenere gli scambi tra le cellule a livello plasmatico, la parete cellulare è provvista
di perforazioni dette punteggiature, nelle quali si insinuano più plasmodesmi.
Le punteggiature quando si è formata soltanto la parete primaria assumono la forma di
piccoli affossamenti sulla parete. Quando poi si viene a formare la parete secondaria
essa non si va ad apporre sopra le punteggiature, anzi, forma quelli che noi chiamiamo i
porocanali.
Le punteggiature si suddividono poi in semplici ed areolate.
Crescita della parete cellulare
Durante la fase di differenziazione, con l’aumento in volume della cellula, anche la
parete è costretta ad accrescersi. Inizialmente, seguendo lo sviluppo della cellula, la
parete si accresce per distensione, poi successivamente in spessore.
Tali forme di accrescimento variano molto nei diversi tipi di cellule. Alcune non cambiano
moltissimo rispetto alla dimensione delle cellule meristematiche, mentre altre si
accrescono anche di 150 volte (vasi).
Nelle cellule parenchimatiche l’accrescimento in spessore è solitamente modesto,
mentre molto più consistente è nelle cellule meccaniche e vascolari.
La crescita per distensione avviene normalmente a carico soltanto della parete
primaria, che come sappiamo presenta una tessitura dispersa, di tipo tubolare.
Durante la crescita per distensione l’orientamento delle fibre viene modificato, e da
una distribuzione intrecciata più o meno obliqua rispetto all’asse principale essa
diventa sempre più parallela all’asse, e quindi di tipo fibroide.
Teoria della multi net growth
Durante la crescita la parete primaria tende a creare anche nuovi strati interni. Questi
inizialmente avranno un tessitura tubolare, poi per l’effetto della distensione della cellula
l’orientamento tenderà a divenire più di tipo fibroide.
Terminata la distensione la parete cellulare aumenta in spessore apponendo al suo
interno la parete secondaria, a tessitura parallela. Quando inizia questa fase
solitamente la parete primaria cessa il suo accrescimento in superficie.
La crescita e lo sviluppo della parete cellulare coinvolge ovviamente tutto il protoplasto.
Infatti nel citoplasma vi è la sintesi di tutti i materiali necessari. Pectine,
emicellulose, cellulose e proteine.
L’apparato del Golgi è l’organello più coinvolto in questa fase di costruzione della parete:
- glicoproteine sono sintetizzate dai ribosomi del reticolo endoplasmatico;
- nel reticolo endoplasmatico la catena polipeptidica è modificata attraverso la
idrossilazione dei residui prolinici presenti (in idrossiprolina);
- tale proteina è poi trasferita all’Apparato del Golgi dove i residui idrossiprolinici
vengono glicosilati con l’arabinosio;
- le proteine così fatte vengono immesse nelle vescicole del Golgi ed inviate verso
l’esterno, dove arrivano a saldarsi con il plasmalemma, in modo tale che i materiali in
esse contenuti possono essere immessi nella parete cellulare.
Oltre alle glicoproteine vengono trasportati polisaccaridi non cellulosici e molte
proteine enzimatiche.
da: “Biologia delle piante” (P.H. Raven et alii)
FORMAZIONE
DELLA PARETE
CELLULARE
Le
microfibrille
di
cellulosa sono sintetizzate
da complessi enzimatici a
rosetta detti cellulosa
sintasi, localizzati nella
membrana plasmatica e
inseriti mediante vescicole
di secrezione provenienti
dal reticolo trans del
Golgi, il cui movimento è
guidato dai microtubuli
corticali.
da: “Biologia vegetale” (A. Bruni, M. Nicoletti)
Ciascuna subunità della rosetta sintetizza 6 catene di glucano (polimero
di β-glucosio) che si assemblano automaticamente a quelle di altre
subunità, per formare una unità microfibrillare. Nelle microfibrille le
catene glucaniche vengono ordinate grazie a legami idrogeno intra- e
intermolecolari tra gli ossidrili dei residui glucosidici, in modo da
formare aggregati parzialmente cristallini.
da: “Biologia delle piante” (P.H. Raven et alii)
I microtubuli corticali giocano un ruolo fondamentale
nell’allineamento delle microfibrille di cellulosa neoformate
nella parete cellulare. Essi corrono paralleli alle microfibrille
depositate al di sopra della membrana plasmatica.
da: “Biologia” (W.K. Purves et alii)
La trama delle microfibrille nella parete
rispecchia l’orientamento dei microtubuli
dello strato corticale della cellula. I
microtubuli fanno in modo che lo
scorrimento dei cellulosa-sintasi avvenga
solo lungo una specifica direzione
attraverso la membrana.