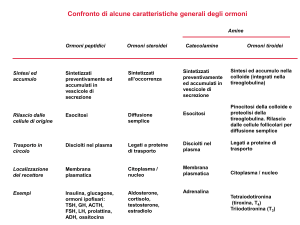
ORMONI
ETILENE
Questo è il mutante etr1 di
Arabidopsis insensibile
all’etilene
ORMONI
ETILENE
la tripla risposta all’etilene:
- accrescimento diageotropico
-inibizione dell’allungamento
dell’epicotile
- accrescimento in spessore
ORMONI
ETILENE: BIOSINTESI
ORMONI
ETILENE: BIOSINTESI
ACC + O2 + ascorbato
Fe2+
C2H4 + CO2 + deidroascorbato
+ HCN + 2H2O
ORMONI
ETILENE: EFFETTI
L’etilene è un ormone da stress
epinastia dei piccioli
ORMONI
ETILENE: EFFETTI
La produzione di etilene è collegata alla maturazione
dei frutti
Andamento della
respirazione e produzione di
etilene in frutti climaterici
ORMONI
ETILENE: EFFETTI
L’etilene induce la senescenza
Il campione a sinistra è
trattato con un sale
d’argento che inibisce la
sintesi di etilene
ORMONI
ETILENE: EFFETTI
L’etilene induce la senescenza
ORMONI
ACIDO ABSCISSICO
ORMONI
ACIDO ABSCISSICO: BIOSINTESI
ORMONI
ACIDO ABSCISSICO: BIOSINTESI
ORMONI
ACIDO ABSCISSICO
La quantità di ABA citosolico è regolata non solo dalla sintesi
ma anche dalla degradazione
Acido faseico e diidrofaseico
e dall’inattivazione
esteri glucosidici
Il trasporto avviene sia via floema che via xilema
In caso di stress idrico della radice viene prodotto ABA che è
traslocato alla chioma dove provoca la chiusura degli stomi
ORMONI
ACIDO ABSCISSICO
L’acido abscissico è coinvolto
nella germinazione
Mutante vp1 di mais
insensibile a ABA
La dormienza è lo stato del seme in
cui è inibita la germinazione.
Può essere legata ai tegumenti
Può dipendere da cause ormonali:
essenzialmente il rapporto ABA/GA
ORMONI
ACIDO ABSCISSICO
L’acido abscissico è coinvolto
nella dormienza delle gemme
ORMONI
ACIDO ABSCISSICO
L’acido abscissico è coinvolto nella chiusura degli stomi
+ KCl 50 mM
+ KCl 50 mM + ABA 10 μM
ORMONI
ACIDO ABSCISSICO
L’acido abscissico è coinvolto nella chiusura degli stomi
ORMONI
ACIDO ABSCISSICO
L’acido abscissico è coinvolto nella chiusura degli stomi
ORMONI
ACIDO ABSCISSICO: MECCANISMO D’AZIONE
L’ABA è coinvolto sia in processi a lungo termine
(maturazione del seme) che in processi a breve termine
(chiusura degli stomi)
I primi implicano cambiamenti dell’espressione genica, i
secondi prevalentemente cambiamenti dei flussi ionici
In entrambi i casi sarà implicata la trasduzione di un segnale
determinato dall’interazione di ABA con una proteina
recettrice
ORMONI
ACIDO ABSCISSICO: MECCANISMO D’AZIONE
Si conoscono almeno 4 tipi di recettori, tre intracellulari e
uno extracellulare. Si conoscono anche diverse vie di
trasduzione del segnale piuttosto complesse.
ORMONI
BRASSINOSTEROIDI
Isolati per la prima volta da granuli di polline di piante di
colza (Brassica napus), sono presenti anche in diverse altre
specie. Il primo composto isolato nel 1979 è il brassinolide.
ORMONI
BRASSINOSTEROIDI
Riconosciuti come ormoni soltanto dopo che fu scoperto che
mutanti di Arabidopsis carenti di brassinosteroidi
mostravano crescita ridotta, foglie piccole, sterilità maschile,
ritardo della fioritura e in generale alterata morfogenesi.
Applicazione esogena di brassinolide ripristinava condizioni
di crescita normali.
Sono ormoni steroidei considerati analoghi agli ormoni
animali. Esistono similarità tra le rispettive vie biosintetiche.
ORMONI
BRASSINOSTEROIDI
Sono ormoni steroidei considerati analoghi agli ormoni
animali. Esistono similarità tra le rispettive vie biosintetiche.
ORMONI
BRASSINOSTEROIDI
Non necessitano di traslocazione poiché sono sintetizzati da
tutti i tipi cellulari.
Stimolano la distensione cellulare in sinergia con auxine e
gibberelline provocando sovraespressione di geni che
codificano per enzimi che controllano la plasticità della
parete.
Si pensa che possano agire anche sul turgore cellulare
regolando l’attività di acquaporine.
Possono anche promuovere la divisione cellulare.
ORMONI
ACIDO SALICILICO
Noto per stimolare la respirazione cianuro-resistente
(Aracee). La produzione di calore provoca la volatilizzazione
di sostanze volatili che attirano gli insetti impollinatori.
Ha anche funzione di stimolazione della resistenza a
fitopatogeni. Induce la produzione di una o più proteine
tipiche della patogenesi (PRP) che determinano la SAR
(systemic acquired resistance).
ORMONI
TURGORINE
Sostanze che agiscono sul turgore delle cellule dei pulvini,
strutture che controllano i movimenti nastici delle piante.
POLIAMMINE
Cationi polivalenti contenenti due o più gruppi amminici per
la presenza di lisina e arginina. Agiscono a concentrazioni
millimolari, hanno effetto di stabilizzazione di protoplasti
isolati, inducono lo sviluppo di alcuni frutti, riducono lo
stress idrico, ritardano la senescenza.
ORMONI
I MOVIMENTI DELLE PIANTE
TROPISMI
Movimenti che prevedono una modificazione del modello di
crescita di un dato organo che porta alla variazione del suo
orientamento in relazione alla direzione dello stimolo.
NASTIE
Movimenti dovuti a variazioni di turgore in modo
indipendente dalla direzione dello stimolo.
NUTAZIONI
Movimenti oscillatori o rotatori che causano cambiamenti
periodici dell’orientamento a causa di una diversa velocità di
crescita su lati opposti degli organi.
ORMONI
I MOVIMENTI DELLE PIANTE
TROPISMI
Oltre al fototropismo e al gravitropismo esistono:
Eliotropismo (inseguimento del sole)
Idrotropismo (risposta all’acqua)
Tigmotropismo (risposta al contatto)
L’eliotropismo è sempre indotto dalla luce blu ma è
provocato da variazioni di turgore di particolari cellule situate
alla base della lamina fogliare dette PULVINI
Esistono due tipi: diaeliotropismo e paraeliotropismo
ORMONI
I MOVIMENTI DELLE PIANTE
Il caso D mostra che poco prima dell’alba le foglie si
riorientano verso la direzione da cui spunterà il sole il che è
indice di un controllo esercitato da un “orologio biologico”
interno (ritmo circadiano).
ORMONI
I MOVIMENTI DELLE PIANTE
ORMONI
I MOVIMENTI DELLE PIANTE
Mentre il diaeliotropismo è chiaramente finalizzato alla
ottimizzazione dello sfruttamento dei raggi solari, il
paraeliotropismo – orientazione parallela ai raggi solari – ha
la evidente finalità di evitare il surriscaldamento e l’eccessiva
perdita di acqua.
Anche in questi casi la luce blu è quella efficace e il
meccanismo coinvolge autofosforilazione di fototropine,
pompe H+-ATPasiche del plasmalemma e movimento di ioni
K+ alla base di variazioni di turgore delle cellule del pulvino.
Curiosamente sono coinvolti gli stessi elementi della
macchina che porta all’apertura degli stomi ma con effetti
opposti: turgore negli stomi e deturgescenza nei pulvini.
ORMONI
I MOVIMENTI DELLE PIANTE
Idrotropismo (risposta all’acqua) della radice
Evidenza di una risposta della radice al gradiente di umidità
nel terreno si è avuta negli anni ottanta grazie a mutanti di
Arabidopsis.
Le cellule del lato esposto a una minore umidità si
accrescono più di quelle del lato esposto a maggiore umidità.
Si ritiene che la percezione dello stimolo avvenga nell’apice
mentre la risposta è nella zona subapicale come nel
gravitropismo ma a differenza di questo non è coinvolta
l’auxina.
Potrebbe essere coinvolta l’idrolisi dell’amido negli statoliti
della columella ma non esistono prove definitive in
ORMONI
I MOVIMENTI DELLE PIANTE
Tigmotropismo (risposta al contatto)
Come negli altri tropismi si tratta di una alterazione della
crescita (morfogenesi) dovuta al contatto (tigmomorfogenesi)
ORMONI
I MOVIMENTI DELLE PIANTE
Tigmotropismo (risposta al contatto)
La tigmomorfogenesi è mediata da diversi segnali chimici:
ormoni, NO, ROS o derivati del metabolismo lipidico
(jasmonati).
Si ritiene che lo stimolo del contatto sia percepito a livello di
plasmalemma e che ciò porti principalmente a un aumento
del Ca2+ citoplasmatico che funge da secondo messaggero
insieme a NO e H2O2.
Indubbiamente la inibizione della crescita delle cellule
“toccate” può coinvolgere l’azione di etilene e ABA nonché la
distruzione di auxina.
ORMONI
I MOVIMENTI DELLE PIANTE
NASTIE
Movimenti che non dipendono dalla direzione dello stimolo
ma che possono essere influenzati dalla sua intensità.
Esempi:
Termonastia (temperatura)
Nictinastia (variazione giorno/notte)
Tigmonastia (contatto)
Sismonastia (scuotimento)
Idronastia
ORMONI
I MOVIMENTI DELLE PIANTE
Termonastia (temperatura)
Tipica dei tepali di certi fiori sembra essere dovuta a
accrescimento differenziale delle cellule epidermiche per
effetti sulla pompa protonica (crescita acida).
ORMONI
I MOVIMENTI DELLE PIANTE
Nictinastia (variazione giorno/notte)
Controllata fondamentalmente dal ritmo circadiano. La luce
non ha effetto se non quello di sincronizzare il ritmo.
La luce blu (fotorecettore sconosciuto) può dare avvio al
ritmo mentre la luce rossa (fotorecettore fitocromo) può far
riconoscere l’inizio del periodo di buio.
Una pianta posta artificialmente a buio continuo mantiene il
movimento nictinastico per un certo tempo.
ORMONI
I MOVIMENTI DELLE PIANTE
Nictinastia (variazione giorno/notte)
Alla base del movimento le variazioni di turgore delle cellule
dei pulvini.
ORMONI
I MOVIMENTI DELLE PIANTE
Nictinastia (variazione giorno/notte)
ORMONI
I MOVIMENTI DELLE PIANTE
Nictinastia (variazione giorno/notte)
ORMONI
I MOVIMENTI DELLE PIANTE
Nictinastia (variazione giorno/notte)
Sembra che in questi movimenti ritmici siano coinvolti fattori
chimici non identificati con azione opposta (LOF e LCF).
L’attività di questi fattori dipenderebbe dal loro stato di
glicosilazione. Esistono prove sperimentali in proposito.
ORMONI
I MOVIMENTI DELLE PIANTE
Tigmonastia (contatto)
Un classico esempio di tigmonastia è quello delle trappole
fogliari (piante carnivore).
In questa attività
sembrano
particolarmente
coinvolte
potenziali d’azione
e acquaporine
ORMONI
I MOVIMENTI DELLE PIANTE
Tigmonastia (contatto) e Sismonastia
La tigmonastia è propria anche di piante che presentano
nictinastia (es. Mimosa pudica detta sensitiva).
In genere sono dette sensitive le piante che chiudono le
foglie in risposta al contatto o ad altri stimoli, quali traumi
meccanici, bruciature, ferite e scuotimento (sismonastia).
Il maccanismo della variazione di turgore è simile a quello
che si ha nella nictinastia ma avviene con più rapidità.
ORMONI
I MOVIMENTI DELLE PIANTE
Sismonastia (scuotimento)
ORMONI
I MOVIMENTI DELLE PIANTE
Sismonastia (scuotimento)
Nei movimenti sismonastici della Mimosa è coinvolto anche
un segnale chimico che su ipotizzato per la prima volta agli
inizi del ‘900 dall’italiano Ubaldo Ricca e fu quindi chiamato
“fattore di Ricca”. Negli anni ‘80 fu isolato un composto
indicato come turgorina che risultò essere acido gallico di
cui fu anche trovato il recettore sulla membrana plasmatica.
Il meccanismo d’azione della turgorina è simile a quello
dell’acetilcolina nelle cellule nervose animali.
ORMONI
I MOVIMENTI DELLE PIANTE
Idronastia
Consiste nella chiusura delle foglie in condizioni di stress
idrico. Tipica delle poacee. Dovuta a variazioni di turgore di
particolari cellule dette “cellule bulliformi”
ORMONI
I MOVIMENTI DELLE PIANTE
NUTAZIONI
Movimenti di flessione autonomi che, se sono rotatori, si
definiscono circumnutazioni.
Sono provocati da una
combinazione di ritmi ultradiani
e forza di gravità. Anche in
questo caso le variazioni di
turgore sono legate a pompe
ATPasiche e canali ionici.
Potrebbe essere coinvolta
anche l’auxina.