
La scoperta dei virus
Prima descrizione di patologia di origine virale: Vaiolo in Cina nel X
secolo a.C.
Alterazioni simil-vaiolose nella mummia del faraone Ramsete II
(circa 2000a.C.).
Tentativi di prevenzione e cura di patologie di origine virale in atto
prima della scoperta dei virus stessi (Jenner, vaccinazione
antivaiolosa alla fine del ‘700).
Prima identificazione di virus: fine dell’800 da parte di Loeffler (afta
epizootica).
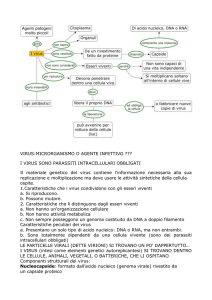
Virus: proprietà distintive
Agenti patogeni in grado di infettare cellule animali, vegetali e
batteriche.
Le particelle virali complete, virioni, sono caratterizzate da
dimensioni estremamente modeste: 10-300 nm.
Sono definiti parassiti endocellulari obbligati perché possono
replicarsi soltanto all’interno di cellule viventi: essi non sono in
grado di produrre e di immagazzinare energia né di provvedere
alla replicazione delle proprie unità costitutive.
Sono caratterizzati
strutturale.
da
una
semplicità
Possiedono particolari modalità di replicazione.
di
organizzazione
Virus: proprietà distintive
I virus rappresentano la forma vivente piu’ semplice. Non hanno una
compartimentazione di tipo cellulare.
Sono, a tutti gli effetti, dei semplici aggregati macromolecolari forniti di, ed in
grado di trasferire, informazioni genetiche.
Alcuni agenti infettanti subvirali (viroidi) sono costituiti da solo materiale
genetico (RNA) senza guscio proteico. Altri, i prioni, sarebbero costituiti da
soli frammenti glicoproteici autoreplicanti, senza acido nucleico. Alcuni di essi
colpiscono il sistema nervoso, determinando malattie mortali tutt’oggi
incurabili (kuru, scrapie, malattia di Creutzfeld-Jacob). Sono al limite tra
forma vivente e non vivente.
Electron micrograph of PRION PROTEIN
(scrapie associated fibrils) Bar=100nm
(Courtesy of Dr. A.C. Scott of Central
Veterinary Laboratory, UK)
Virus: proprietà distintive
Date le loro caratteristiche, i virus sono assolutamente incapaci di
replicarsi al di fuori delle cellule bersaglio.
Sopravvivono un tempo limitato (pochi minuti o alcuni giorni, a
seconda del tipo virale) nell’ambiente extracellulare, senza mai
replicarsi fuori dalle cellule.
Come parassiti intracellulari obbligati, i virus necessitano non solo
degli elementi nutritivi prodotti dalle cellule ospiti, ma soprattutto
dei sistemi di trascrizione e traduzione di tali cellule.
Contengono enzimi propri indispensabili per il ciclo replicativo
(polimerasi, proteasi, ecc). Tutti gli altri enzimi sono resi disponibili
dalla cellula ospite.
Virus: proprietà distintive
I virus attraversano i comuni filtri, sono invisibili al microscopio ottico
(con l’eccezione di alcuni virus particolarmente grandi, quali i poxvirus)
ma visibili al microscopio elettronico (meglio a trasmissione).
Determinano spesso, nelle cellule infettate, la presenza di corpi inclusi
intranucleari o citoplasmatici.
Micrografo con i virus numerosi di rabbia (piccolo scuro-grigio
asta-come le particelle) ed i corpi di Negri, più grandi inclusioni
cellulari tipiche dell'infezione di rabbia.
Virus: proprietà distintive
Sono coltivabili in vitro in cellule isolate da organismi, o in batteri
(a seconda del tipo virale).
Alcuni (Orthomyxovirus, paramyxovirus) possono essere coltivabili
con facilita’ in uova embrionate.
Hanno resistenza variabile al pH. Questo regola la loro patogenicita’
- Es. Rinovirus sono distrutti a pH3 (pH gastrico di circa 1) e
pertanto sono non patogeni per l’apparato gastrointestinale.
- Altri picornavirus sono resistenti al pH gastrico e pertanto
patogeni (enterovirus).
Virus: proprietà distintive
I virus rappresentano un sistema altamente
sofisticato di trasferimento genetico da cellula a
cellula.
Tramite i virus, e’ possibile modificare il corredo genetico (e
quindi fenotipico) di una cellula, favorendone, quindi,
l’acquisizione di nuovi caratteri.
Data questa loro caratteristica naturale, sono usati, nella
manipolazione genetica, come vettori finalizzati al trasferimento di
nuovi caratteri a cellule infettabili da un determinato virus.
Virus: proprietà distintive
Sono concentrabili in vitro solo con l’uso delle ultracentrifughe
(20000 e piu’ giri al minuto). La ultracentrifugazione su gradiente di
densità permette di separare particelle virali della stessa specie
virale, ma aventi densità leggermente diversa a causa di quantità
diverse di proteine o di acido nucleico presente.
L’esame al microscopio elettronico permette sia la conta delle
particelle virali, sia la loro caratterizzazione morfologica.
Human immunodeficiency viral particles are seen at
medium magnification in this electron micrograph.
Virus: proprietà distintive
A seconda del loro specifico tropismo, possono infettare
cellule vegetali (virus del tabacco, della patata, ecc),
batteri (batteriofagi, ognuno specifico per una
determinata specie batterica), e cellule animali (virus di
insetti, pesci, uccelli, mammiferi).
Electron micrograph of TMV particles
VIRUS H5N1 O
INFLUENZA AVIARIA
BATTERIOFAGO
Virus: proprietà distintive
I VIRUS SONO INSENSIBILI
AGLI ANTIBIOTICI
Costituenti virali
Core
Acido nucleico più ogni molecola che ne
determina la stabilità
Capside
Struttura proteica che racchiude l’acido nucleico o
il core
Capsomero
Unità proteica che, ripetuta, forma il capside
icosaedrico
Nucleocapside Acido nucleico più capside
Envelope
Involucro lipoglicoproteico esterno
Peplomeri
Proiezioni superficiali che protrudono dall’envelope
Virione
Particella virale completa come si può osservare
al di fuori della cellula
Composizione chimica
Virus nudi
acido nucleico
proteine
Virus rivestiti
(con envelope)
acido nucleico
proteine
lipidi
carboidrati
Lipidi e carboidrati sono contenuti nella membrana limitante
esterna: sono identici a quelli della cellula ospite e non sono
codificati dal virus
Proteine virali
Si distinguono:
Proteine funzionali che permettono ad es. all’acido nucleico
virale di replicarsi (DNA-o RNA-polimerasi)
Proteine strutturali: incorporate nelle nuove particelle virali
come costituenti del capside o come strutture particolari (es. le
emoagglutinine) presenti nell’envelope
Proteine che alterano alcune funzioni e strutture della
cellula ospite
Proteine strettamente associate agli acidi nucleici
Proteina di matrice o M
MORFOLOGIA VIRALE
I virus, nella loro forma extracellulare (virione), hanno una
struttura cristallizzata tipica
Di contro, all’interno delle cellule in cui si replicano, non sono mai
reperibili in forma completa, eccetto che poco prima della loro
uscita dalla cellula (fase di gemmazione)
Di norma, all’interno delle cellule e’ possibile trovare materiale
genetico del virus, e proteine virali piu’ o meno assemblate
HIV
Morfologia virale
La struttura di base dei virus consiste nell’acido nucleico e in un
rivestimento proteico detto capside.
Il genoma contiene tutta l’informazione genetica necessaria per la
replicazione; è molto labile ad agenti denaturanti (nucleasi, esterasi, forza
ionica ecc.) e non può da solo penetrare nella cellula ospite .
La quantità di acido nucleico è indice della complessità strutturale del
virione.
Il capside costituisce una struttura continua e impermeabile agli agenti
esterni e protegge l’acido nucleico. Alcune proteine che lo compongono
possiedono una specifica affinità per particolari recettori della membrana
citoplasmatica della cellula ospite. Queste strutture permettono il legame del
virus alla cellula e la penetrazione dell’acido nucleico.
Alcuni virus posseggono una membrana limitante esterna detta envelope, (o
pericapside o peplos).
I virus con il solo capside vengono definiti “nudi”, quelli con envelope
vengono definiti “rivestiti”.
I virus più grandi possono avere una struttura più complessa.
Pericapside (peplos, envelope)
Strato lipidico (in alcuni casi glicoproteico) esterno all’involucro, presente solo
in alcune famiglie di virus (orthomyxovirus, herpesvirus, retrovirus, poxvirus,
ecc.).
E’ costituito da lipidi della membrana della cellula infettata, rimasti intorno al
virus vero e proprio al momento della fuoriuscita dalla cellula stessa
(“budding”).
Contiene anche alcune glicoproteine virali, che fungono da primo recettore
virale nei confronti delle cellule bersaglio.
Svolge funzione di protezione, di riconoscimento antigenico (emoagglutinina
del virus influenzale) e di penetrazione nella cellula ospite tramite il
meccanismo di fusione con la membrana cellulare.
La presenza del pericapside rende ragione della sensibilita’ di alcuni virus ai
disinfettanti organici (eteri, alcooli) in grado di sciogliere i lipidi.
Componenti dell’involucro virale
(envelope)
La base di tutti gli involucri
virali è una membrana lipidica
acquisita dalla cellula ospite
durante l’assemblaggio.
Proteine virali vengono inserite
nella membrana lipidica.
La grande maggioranza di
queste sono glicoproteine
(oligomeri).
Funzioni delle proteine dell’involucro
Aggancio, fusione ed entrata
(es. HA)
Attività enzimatica
(es. NA)
Canale ionico
(es. M2)
Antigeni principali
(es. HA, NA)
Orthomyxovirus
Virus con involucro semplice
Contatto diretto tra proteine esterne e capside
Sezione equatoriale di un Togavirus con ricostruzione dell’immagine
Virus con involucro a livelli proteici
addizionali
Struttura di un herpesvirus
Capside
E’ proprio del virus, e’ geneticamente determinato e costituito da
subunità proteiche disposte in modo regolare, codificate dal
genoma virale.
Dato il limitatissimo patrimonio genetico dei virus, le proteine
costituenti i capsomeri (unità base del capside) sono molto poche,
specifiche del virus, e in grado di interagire con i componenti sulla
superficie cellulare fungendo da recettori.
Conferisce resistenza al materiale genetico virale nei confronti delle
nucleasi presenti nei fluidi.
Simmetrie del capside
Il capside è costituito da proteine codificate dal genoma virale la
cui ridotta lunghezza permette una sintesi limitata di proteine. Il
capside in genere è quindi costituito da unità ripetitive di una
o poche proteine.
Le proteine destinate a formare il capside sono capaci di
autocombinarsi
(assemblaggio)
seguendo
due
schemi
fondamentali definiti:
-a simmetria elicoidale
-a simmetria cubica (detta anche icosaedrica)
La simmetria virale, dovuta all’interazione genoma-proteine, puo’
essere:
a. Elicoidale, quando l’acido nucleico si avvolge sulla struttura
proteica di base, determinando, al microscopio elettronico, una
forma oblunga (virus del mosaico del tabacco, orthomyxovirus,
rhabdovirus., ecc). Tutte le subunita’ proteiche sono identiche tra
loro
b. Icosaedrica, quando i capsomeri (unita’ base dell’icosaedro
costituente il capside) sono organizzati a costituire una struttura
poliedrica regolare con 20 facce costituite da triangoli equilateri.
All’interno e’ situato il nucleocapside, aderente in alcuni punti ai
capsomeri. E’ presente nella maggioranza dei virus
c. Complessa, A struttura non classificabile secondo canoni ristretti:
batteriofagi, poxvirus, ecc.
Tutti i virus di un determinato tipo saranno assolutamente
uguali tra loro
Virus a simmetria elicoidale
Le sub-unità proteiche, dette protomeri, si dispongono lungo
un asse elicoidale intorno all’acido nucleico a formare il
nucleocapside che assume struttura tubulare.
Nella maggior parte dei casi i virus animali con questa
simmetria possiedono l’envelope.
Vari nucleocapsidi si differenziano per lunghezza, diametro,
passo dell’elica e numero di protomeri per spira.
Gli involucri
pleiomorfa.
pericapsidici
conferiscono
ai
virioni
forma
Influenza virus
Virus della rabbia
Rabdovirus yemando de una inclusión (cuerpo de Negri) hacia el retículo
endoplásmico en una célula del sistema nervioso.
A. Cuerpo de Negri. B.
Note la abundante ribonucleoproteína en la inclusión. C. Rabdovirus
yemando. CDC
Rabdovirus en una célula epitelial de un pez © Dennis Kunkel Microscopy, Inc
Virus a simmetria icosaedrica
Il capside in questo caso costiuisce una sorta di guscio per il
genoma.
Le sub-unità proteiche costituite da una singola molecola
polipeptidica o da un aggregato di molecole peptidiche si
riuniscono a formare le unità morfologiche, dette capsomeri, del
capside.
I capsomeri si combinano a formare solidi regolari.
L’icosaedro è un solido costituito da 20 facce triangolari e 12
vertici.
I vari virus con questa simmetria si differenziano per numero e
distribuzione dei capsomeri.
Alcuni sono virus nudi, altri posseggono l’envelope.
Ricostruzione di differenti particelle virali a
simmetria icosaedrica
Caratteristiche virali
Elementi caratterizzanti alcune famiglie di virus:
a. Rotavirus: doppio capside (struttura a ruota).
b. Adenovirus: estroflessione dal capside che funge da recettore
virale sulla superficie cellulare.
c. Arenavirus: aspetto sabbioso determinato dall’inglobamento di
ribosomi durante il ciclo replicativo intracellulare.
Arenavirus
Electron micrograph of
human adenovirus
Electron micrograph of rotavirus from stool sample
Struttura dei capsidi virali
Simmetria complessa
poxvirus
Simmetria complessa
Batteriofago T4: simmetria complessa
Core (o nucleocapside)
Area interna del virus, in cui si trova il materiale genetico del
virus (a RNA o a DNA, a seconda del tipo virale), frammisto a
proteine anch’esse proprie del virus, in parte strutturali (che
impacchettano e stabilizzano il genoma) e in parte funzionali
(polimerasi, proteasi, ecc.).
Puo’ essere compatto (Alfavirus), Filamentoso (Paramyxovirus),
multisegmentato (Ortomyxovirus).
Capside virale
Perché la costruzione di una sub-unità
è comune a tutti i virus?
Auto assemblamento
Nel 1955, Fraenkel-Conrat e Williams dimostrarono che quando
miscele di RNA di virus del mosaico del tabacco purificato (TMV) e
proteina coat venivano incubate insieme, si formavano particelle di
virus elicoidale. Questo indica che la particella di virus si ordina da
sola e predispone la struttura con la minima energia libera, ad es. la
più stabile. Incorporare copie multiple di una o diverse sub-unità è
presumibilmente un modo semplice per portare a termine tutto ciò.
Capside virale
I virus nudi di origine animale
esclusivamente di tipo icosaedrico.
hanno
una
simmetria
I virus con envelope possono avere il nucleocapside sia a
simmetria elicoidale sia a struttura icosaedrica.
I virus più grandi (come i Poxvirus) possono avere una struttura
morfologica più complessa.
Genoma virale
Tutti i virus contengono un solo tipo di acido nucleico.
Quelli a RNA, in particolare, rappresentano un’eccezione in natura, in
quanto la loro informazione genetica non e’ contenuta nel DNA. A
seconda del tipo virale, il genoma codifica per un numero diverso di
proteine, da un minimo di 3 a un massimo di alcune centinaia.
A seconda dei vari tipi di virus, il genoma (DNA o RNA) puo’ essere a
singolo o doppio filamento, lineare o segmentato in varie porzioni (es.
virus influenzale) ognuna codificante per una proteina.
Tra i virus a RNA, alcuni hanno il filamento cosiddetto positivo (che
funge anche da RNA messaggero), altri hanno il filamento negativo (che
deve sintetizzare, sul suo stampo, un filamento complementare di RNA
che fungera’ da messaggero).
Genoma dell’epatite B
Genoma virale
E’ costituito da un solo tipo di acido nucleico:
DNA o RNA
Virus a RNA: Ribovirus
Possiedono un RNA a singola elica (tranne la famiglia dei Reovirus
che ha un RNA a doppia elica), del peso di circa 2-10 x 106 daltons,
a volte (orthomyxovirus) diviso in frammenti. Nei reovirus e’ a
doppio filamento segmentato (piu’ resistente del singolo filamento
all’RNAsi e ai disinfettanti).
VIRUS AD RNA
Si dividono in 2 sottoclassi:
Virus con RNA a polarità positiva (+) che può fungere nella cellula
direttamente da RNA messaggero.
Virus con RNA a polarità negativa (-) che funge da stampo per la
sintesi dell’RNA messaggero- questi virus hanno l’enzima RNA
polimerasi RNA dipendente associato al virione.
I Reovirus e alcune famiglie di virus a RNA a polarità negativa
hanno un genoma con sequenze indipendenti di RNA (virus
influenzale con 8 frammenti, bunyavirus con 3, Reovirus con 12).
Genoma virale
Virus a DNA: Deossiribovirus
Normalmente hanno un DNA a doppia elica con filamento
unico e lineare.
Alcuni virus (Papovavirus ed Hepadnavirus) contengono
DNA circolare e altri (Parvovirus) hanno un DNA
monocatenario.
La grandezza del genoma dà la misura della complessità del
virus e della sua replicazione (più evidente nei virus a DNA
rispetto ai virus a RNA).
VIRUS A DNA
DNA: La molecola di DNA e’ unica, del peso da 1,5 x 106 daltons
(parvovirus, gran parte dei batteriofagi) a 200 x 106 daltons
(poxvirus), solitamente a doppio filamento lineare, con l’eccezione dei
parvovirus (singolo), papovavirus (doppio filamento circolare),
hepadnavirus (doppio filamento incompleto, circolare).
Il contenuto in guanina-citosina nei retrovirus (che si integrano nel
genoma delle cellule ospiti) e’ simile a quello riscontrato nelle cellule
stesse (circa 50% del totale), e questo probabilmente favorisce
l’interazione tra i due DNA.
Enzimi virali
Altri componenti virali
Proteine non strutturali
Aiutano la replicazione virale ma non sono incorporate nel
virione
Enzimi necessari per la sintesi e la replicazione del DNA
(es. RNR, TK, DNA polimerasi di Herpesviruses)
Proteine che influiscono sulla fisiologia della cellule
(es. E6/E7 di papillomavirus umani)
Proteine che sopprimono le difese della cellula ospite
(es. Inibitori della presentazione di antigeni)
Alcune forme virali
Adenovirus
Alcune forme virali
Papillomavirus
Alcune forme virali
Herpesvirus
Alcune forme virali
Paramyxovirus
Orthomyxovirus
Alcune forme virali
Vesicular stomatitis virus
Alcune forme virali
Human immunodeficiency virus
Alcune forme virali
Ebola virus
Classificazione virale
Si basa su criteri morfologici, strutturali, chimici e replicativi:
forma,dimensioni, tipo e struttura del genoma, strategia replicativa
In base a queste caratteristiche i virus vengono suddivisi in:
Famiglie (con suffisso –viridae)
Sottofamiglie (generalmente con suffisso –virinae)
Generi (con suffisso –virus)
Specie
Ulteriori suddivisioni in sottospecie, tipi, ceppi ecc. vengono di volta
in volta stabilite da apposite commissioni per la nomenclatura in
base a criteri che risultino via via accettati.
Classificazione virale
Nella pratica medica si continuano a indicare i virus secondo la
nomenclatura comune:
con il nome “virus” seguito da quello della malattia che determina
nell’ospite principale (per es. virus del morbillo, dell’influenza, della
poliomielite ecc.)
oppure
con il nome che gli è stato attribuito al momento dell’isolamento
(per es. virus Ebola, di Epstein-Barr, Norwalk ecc.).
Classificazione virale
L’attuale classificazione si basa:
sul tipo di genoma (DNA o RNA) e la sua configurazione (mono o
bicatenario, lineare o circolare, monofilamento o segmentato, a
polarità positiva o negativa)
sul tipo di simmetria
sulla presenza di involucro pericapsidico
sul meccanismo di replicazione
Molti virus non sono stati ancora classificati oppure lo sono solo
provvisoriamente
Per alcuni agenti non è ancora stata accertata la natura virale
(per es. gli agenti delle encefaliti spongiformi dell’uomo e di vari
animali)
Classificazione dei virus
Criteri di classificazione
International Committee on
Taxonomy of Viruses
• Ospiti: animali, piante, batteri.
• Natura dell’acido nucleico nel virione : RNA o DNA
• Simmetria del capside: icosaedrico, elicoidale o
complesso
• Presenza o assenza di un involucro
• Architettura del genoma: ds, ss, frammentato, dimensione
• Omologia di sequenza
• Strategie di replicazione
Nomenclatura: alcune regole basilari
•
•
•
•
•
Ordine ( - virales)
Famiglia ( - viridae)
Sottofamiglia ( -virinae)
Genere ( - virus)
Specie (common names)
Nomenclatura: alcune regole basilari
Picornaviridae
famiglia
genere o sottofamiglia
enterovirus
cardiovirus
rhinovirus
aphthovirus
hepatovirus
specie
poliovirus
67 sierotipi
mengovirus
rinovirus
> 100 sierotipi
FMDV-C
virus dell’epatite A
Il sistema di classificazione Baltimore
Class II
ss DNA
Class I
ds DNA
Picornavirus
Flavivirus
Togavirus
Adenovirus
Herpesvirus
Papovavirus
Poxvirus
Orthomyxovirus
Paramyxovirus
Rhabdovirus
Parvovirus
(B19)
Retrovirus
Class VI
Reovirus
ss RNA (+)
Class IV
Class V
Class III
ss RNA (+)
ss RNA (-)
ds RNA (+/-)











![Lezione 15 Virus [modalità compatibilità]](http://s1.studylibit.com/store/data/000771737_1-84b1cca561c5813066d1b76125338a98-300x300.png)