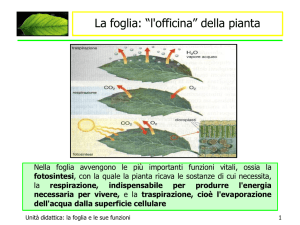
Energia di stoccaggio d’acqua nella vegetazione
Prendiamo in considerazione un sistema suolo-pianta-atmosfera chiuso e calcoliamone
rispettivamente il bilancio di energia e di massa ( fig. a-b ).
Nel primo caso ( fig.a) il bilancio sarà rappresentato dall’equazione Q∗ = Qh + Qe + ∆Qs + ∆Qp
dove Qh rappresenta il flusso di calore sensibile ( flusso legato alle variazioni di temperatura) , Qe il
flusso di calore latente ( correlato alle variazioni di stato ) ; ∆Qs è il calore fisico stoccato e ∆Qp è
l’energia biochimica stoccata ( dipende dalla CO2 e dal tipo di pianta ).
Il valore di ∆Qp può essere scritto come φ∆P : Φ, la cui unità di misura è W/m, è il calore di
assimilazione del carbonio; ∆P , la cui unità di misura è Kg/ms, è dato dalla differenza tra
l’assorbimento della CO2 da parte della pianta attraverso la fotosintesi e l’emissione di CO2 per
respirazione . Perciò il valore di ∆P e, di conseguenza, di ∆Q dipende da giorno e notte : di giorno l’
assorbimento di CO2 sarà maggiore rispetto all’emissione e quindi avremo un ∆P positivo e un ∆Q
positivo; di notte la situazione sarà inversa, l’emissione sarà maggiore dell’assorbimento quindi ∆P
e ∆Q assumeranno valore negativo.
Nel secondo caso ( fig.b ) il bilancio sarà rappresentato dall’equazione p = E + ∆r + ∆S dove p è
la precipitazione, E l’evapotraspirazione, ∆r lo scorrimento superficiale, ∆S lo stoccaggio netto di
acqua nell’aria e nel suolo.
Ricordiamo che per semplificare il sistema studiato facciamo riferimento a scambi verticali quindi
, all’interno delle nostre equazioni non abbiamo considerato i valori di ∆Qa ovvero il calore latente
sensibile trasportato orizzontalmente per avvezione nel sistema e il valore di ∆A correlato allo
scambio di umidità per avvezione. Inoltre, nel caso del bilancio di massa, non abbiamo preso in
considerazione gli spostamenti d’acqua all’interno del sottosuolo.
Flusso di CO legato all’attività stomatica
Lo scambio di CO2 e di vapore acqueo tra pianta e atmosfera è mediato dall’attività stomatica. Gli
stomi sono valvole il cui grado di apertura è determinato dall’intensità della luce, dalla
concentrazione di CO2, dall’umidità e dalla temperatura ambientale. Quindi , di giorno, gli stomi
sono aperti per catturare ed espellere CO2, di conseguenza l’evapotraspirazione è elevata. La
perdita d’acqua induce un movimento di nutrienti ed umidità attraverso la pianta e a questo è
associata una dispersione di calore latente maggiore attraverso le foglie. Si può dunque sostenere
che l’evapotraspirazione sia un ottimo sistema di dissipazione del calore in eccesso acquisito dalla
pianta.
Oltre a questa caratteristica fisiologica dobbiamo considerare anche la struttura stessa
dell’apparato fogliare della pianta poiché in molti casi il sito di scambio sono le foglie ed è quindi
importante considerare la variazione verticale della loro densità : l’architettura è in grado di
influenzare scambi di calore e di massa. Nella nostra relazione parleremo spesso di CANOPY come
aggregazione di più strati fogliari. La struttura della canopy varia a seconda del tipo di vegetazione :
se abbiamo a che fare con foreste tropicali la struttura è multistratificata, nel caso di un campo di
mais, girasoli e patate la densità fogliare è concentrata al top quindi, parliamo di un unico strato.
La densità della massa fogliare non solo interessa gli scambi con l’atmosfera ma interferisce
anche con la velocità del vento. Osservando infatti, il grafico 3, notiamo come la velocità del vento
misurata ad una serie di livelli sopra l’altezza h della vegetazione tenda a diminuire sino a 0, ad un
livello tra h e d.
Proprio tra h e d si ha la massima resistenza esercitata dalla vegetazione. Il valore di d dipende
dalla velocità del vento e dal tipo di vegetazione poiché la maggior parte di essa è flessibile e tende
ad assumere una forma più aerodinamica a velocità elevate.
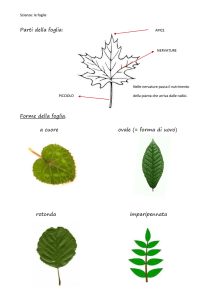
Poiché il sistema vegetale è composto da molte superfici attive
rappresentate
da
una
miriade
di
foglie,
analizziamo
le
caratteristiche di una foglia tipica.
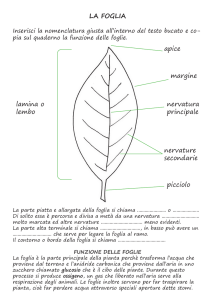
Budget radiativo di una foglia
Le foglie non sono opache alla radiazione corta.. Osserviamo la firma spettrale di una foglia,
rappresentata dal grafico 4: sull’asse delle x abbiamo le diverse lunghezze d’onda, su quello delle y
la riflettività e la trasmissività. Il range tra 0.4 e 0.7 µm viene chiamato PAR radiazione
fotosinteticamente attiva; e infatti, notiamo che in corrispondenza di 0.4 ( λ del blu ) e di 0.7 ( λ del
rosso ) abbiamo due picchi di assorbimento dovuti all’attività fotosintetica della foglia, mentre in
corrispondenza di 0.6 ( λ del verde ) abbiamo un picco di riflettività legata alle cellule palizzata (
per questo noi vediamo le foglie verdi ). Nel NIR ( infrarosso vicino ) si osserva una forte
riflessione dovuta al tessuto mesofiliare. Da 1.5 in poi notiamo un forte assorbimento dovuto alla
presenza di acqua all’interno della foglia.
L’alta riflessione a lunghezze d’onda più lunghe aiuta la foglia a restituire calore all’ambiente e
quindi a moderare la moderare la temperatura fogliare.
Nella figura a viene schematizzato il bilancio radiativo di una foglia: le onde corte che
raggiungono lo strato fogliare vengono riflesse, le onde lunghe vengono trasmesse.
Bilancio energetico di una foglia
Il bilancio energetico di una foglia è rappresentato dalla somma di Qe è di Qh. Confrontando con
il sistema chiuso iniziale notiamo l’assenza dei valori ∆Qp e ∆Qs : questi valori vengono trascurati
poiché per una singola foglia risultano essere quantitativamente insignificanti. Il calore acquisito
dalla foglia a lunghezze d’onda corte è dissipato dalla radiazione a onde lunghe attraverso la perdita
di calore sensibile e latente.
Il calore latente è perso grazie all’evapotraspirazione dell’acqua sulla parte esterna della foglia
che intercetta pioggia ed umidità oppure attraverso la traspirazione dell’acqua regolata dagli stomi.
Il trasferimento di calore sensibile, invece, avviene tra superficie della foglia e l’aria circostante.
Possiamo così scrivere Qh = Ca ( To – Ta ) / rb : To è la temperatura della superficie della foglia,
Ta è la temperatura dell’ambiente circostante, Ca la capacità di calore dell’ aria; rb rappresenta la
resistenza diffusiva di un substrato laminare aderente alla foglia. Il suo valore aumenta con l’
aumentare delle dimensioni della foglia e diminuisce con l’aumentare della velocità del vento.
Quindi per foglie piccole, in giorni ventilati o in condizioni instabili rb sarà minore e , di
conseguenza, Qh sarà maggiore.
Trasferimento di vapore acqueo dalla foglia all’ambiente circostante.
Fin’ora abbiamo elencato i fattori che intervengono a livello della foglia per attuare
evapotraspirazione ovvero l’attività stomatica e la resistenza diffusiva. Quindi, riassumendo in
un’equazione l’attività di trasferimenti di vapore d’acqua , quest’ultimi non possono essere
trascurati,. Scriveremo dunque:
E = ( ρvto - ρva ) / ( rb + rst )
Dove ρvto è la densità di saturazione del vapore alla temperatura superficiale della foglia, ρva la
densità di vapore acqueo, rst la resistività degli stomi.
Attraverso l’attività stomatica e quindi il controllo di evapotraspirazione, la foglia riesce a
controllare la propria temperatura. Prendiamo in considerazione l’immagine 6 in cui è rappresentata
la superficie fogliare di un fagiolo e poniamo come condizioni circostanti la velocità del vento pari
a 0.7 m/sec, Ta di 25.6° , Q* 25.6° W/m ed rb da 400 a 1300 s/m. A seconda dell’esposizione al
sole la superficie della foglia presenterà diverse temperature ( nel nostro caso 7.7 corrisponde a
massima esposizione, 3.9 a minima esposizione ). Si può osservare che la temperatura della foglia
può raggiungere +10° rispetto alla temperatura dell’aria. In giornate molto calde le cellule di
guardia degli stomi si chiudono per conservare l’acqua. Le foglie sono in grado di adattarsi a forti
stress climatici attraverso l’assorbimento ridotto di Q* , una forte albedo o aumentando
l’evapotraspirazione se la foglia è piccola. L’acqua conservata dalla cuticola fogliare riduce
l’attività degli stomi aumentando l’albedo della foglia. Il valore dell’ albedo non è solo funzione
dell’acqua contenuta nella foglia ma dipende da altri due fattori ( grafici 11 e 12 ) : l’ altezza della
vegetazione e l’inclinazione del sole rispetto all’orizzonte. Come si riscontra nel grafico 11 , mano a
mano che aumenta l’altezza della vegetazione, il valore dell’albedo diminuisce. Nel grafico 12 ,
invece, notiamo come all’aumentare della posizione del sole rispetto all’atmosfera diminuisca il
valore dell’albedo. A questo punto interviene un ulteriore fattore detto di vista del cielo ( ψsky ) che
influenza l’emissione a onde lunghe della mia vegetazione; maggiore è questo valore, maggiore è
l’emissione a onde lunghe. Quindi, in un campo a vegetazione bassa ( short grass ) i valori di albedo
sono elevati ma tendono a diminuire mano a mano che il sole si alza, nel caso di querce ( oak ) i
valori sono più bassi e ancor di più diminuiscono, sempre in funzione dell’altezza del sole.
Trasferimento di CO2 dalla foglia all’ambiente circostante.
Come per il trasferimento di vapore acqueo, anche in questo caso il trasferimento di CO2 può
essere scritto sotto forma di equazione tenendo sempre in considerazione rst e rb : Fc = ( ρca – ρci )
/ ( rb + rst ) con ρca pari alla concentrazione di CO2 nell’ambiente e ρci pari alla concentrazione
di CO2 all’interno della foglia.
Concludiamo questa prima parte inerente ai bilanci energetici e di
massa con
considerazioni ed esempi inerenti all’intero sistema
vegetale.
Bilancio e flussi interni in un sistema suolo-pianta- atmosfera ( assenza
irrigazione )
L’efficienza d’intercettazione delle precipitazioni dipende dall’intensità e dal tipo di pioggia ma
anche dalle caratteristiche della canopy, dalla densità e dall’area fogliare. L’acqua raggiunge il
terreno per precipitazione diretta, per gocciolamento delle foglie o per flusso nel gambo. L’acqua si
può infiltrare nel suolo o restare sul suolo sottoforma di pozze. L’acqua può essere stoccata nel
suolo e risalire per capillarità. Durante la crescita stagionale della pianta, la maggior perdita di H2O
avviene con la traspirazione; l’H2O può essere persa dal sistema attraverso l’evaporazione del suolo
o della cuticola fogliare ( in questo caso si parla di guttazione). Il movimento di acqua nel suolo
dipende dalla conduttività idraulica del suolo. Il flusso netto di acqua nel sistema pianta à funzione
delle differenti concentrazioni di vapore tra foglie e aria e della differenza di potenziale fra foglia e
suolo. Questo processo è rappresentato in figura 7.
Variazione della concentrazione di CO2 all’interno del sistema vegetale.
A seguito delle considerazioni fin’ora esposte facciamo un bilancio diurno e annuale della
concentrazione di CO all’interno della pianta e nell’ambiente circostante.
Nell’arco delle ventiquattro ore la pianta acquisterà maggior quantità di CO2 ,tramite attività
fotosintetica, a metà giornata. Quindi, nell’ambiente circostante, questo sarà il momento in cui la
quantità di CO2disponibile in atmosfera raggiungerà livelli minimi. Al contrario, nella fase notturna
, il flusso di CO2 sarà negativo all’interno della pianta ( la CO2 viene rilasciata tramite la
respirazione ) mentre la concentrazione in atmosfera sarà molto elevata. L’intensità di questa
attività, però, varia a seconda della fase vitale della nostra pianta : da Maggio a Luglio si nota un
incremento di Fc poiché ho la fase di crescita della pianta, ma sul finire della stagione la Fc
diminuisce poiché la pianta va in senescenza. In inverno la situazione raggiunge i minimi livelli. A
livello dell’atmosfera da Maggio a Luglio la differenza tra valori diurni e notturni è notevole, mano
a mano che ci si avvicina all’inverno la disparità tende ad annullarsi. ( vedi grafici 8 e 9 ).
Variazione di K e Q* all’interno della vegetazione
Come possiamo leggere dai grafici 10-a e 10-b, mano a mano che ci si sposta verso il suolo, i
flussi di K e di Q* diminuiscono. Questo si verifica perché la superficie vegetativa attiva assorbe la
radiazione diretta attuando la fotosintesi e , ostacolando le zone più basse, non permettete che
queste vengano raggiunte da altrettanta K e che attuino con la stessa efficacia l’ attività
fotosintetica. Maggiori sono i valori di K e Q , più veloce sarà la diminuzione come dimostra la
seguente legge: K z = Ko e
dove K z è la radiazione dalle onde corte ad una certa altezza, a è
il coefficiente di estinzione dovuto alle foglie, A z è l’area fogliare di accumulo al top della canopy
e K o è la radiazione ad onde corte ricevuta sopra la canopy
LA MICROMETEOROLOGIA DELLE SUPERFICI
VEGETATE
Bilancio radiativo, energetico e resistività della canopy. (campo di
orzo-Inghilterra 23 giugno 1963)
Dalla fig 4.15 (a) si può notare come il massimo di radiazione a onde corte si abbia
durante le ore centrali della giornata e mostri 2 picchi. Questo andamento potrebbe
essere il risultato di un passaggio nuvoloso intorno alle ore 14.
Il bilancio energetico totale è positivo e riflette l’andamento della radiazione a onde
corte entrante.
Il massimo di onde corte uscente lo abbiamo in corrispondenza del massimo di onde
corte entranti.
Il bilancio a onde lunghe risulta essere negativo.
Nella fig 4.15(b) si nota che in questo sinora che in questo sistema vegetato il miglior
sistema per dissipare energia avviene attraverso emissione di calore latente di
evapotraspirazione.
Nella fig 4.15(c ) si osserva l’andamento durante la giornata della resistività della
canopy.
La resistività è bassa alle prime ore del mattino poiché la canopy è ricoperta da rugiada.
Si mantiene costante durante la giornata poiché gli stomi sono aperti.
In serata invece subisce un forte aumento dovuto ad un decremento di luce o ad un
possibile stress d’acqua, i quali contribuiscono a chiudere gli stomi.
Stress idrico di un campo irrigato
Il campo è stato irrigato il 28 maggio, in seguito si sono avute condizioni secche e molto
soleggiate.
Nella fig 4.16 (a) si nota con il trascorrere dei giorni un forte incremento del potenziale
d’acqua nel suolo e un successivo aumento della resistività della canopy tali da portare
la pianta ad un intenso stress idrico.
Nella fig 4.16 (b) si nota la variazione di evapotraspirazione e resistività della canopy
dopo 23 e 31 giorni dall’irrigazione del campo.
L’evapotraspirazione dopo 31 giorni è diminuita poiché la pianta tende a conservare
l’acqua mentre la resistività aumenta poiché gli stomi tendono a restare più chiusi.
Profilo della velocità del vento, della temperatura, della pressione di
vapore e della concentrazione di anidride carbonica dentro e sopra un
campo di orzo
Nella fig 4.17 (a) si osserva che il profilo della velocità del vento durante le ore notturne
si mantiene piuttosto costante.
Durante le ore centrali della giornata (11-12) il profilo subisce una notevole variazione.
Infatti la canopy riduce la velocità del vento fino a valori di 0 m/s al suolo.
Il massimo del decremento della velocità del vento lo si riscontra dove si ha il max della
densità fogliare.
Nella fig 4.17 (b) si nota invece il profilo verticale della temperatura.
Nel suolo si osserva che l’onda di calore arriva con una sfasatura rispetto all’atmosfera
sovrastante.
Infatti nel suolo durante la notte si registrano temperature maggiori rispetto
all’atmosfera mentre di giorno avviene l’opposto.
All’interno dello strato di aria influenzato dalla vegetazione si nota un massimo di
temperatura durante le ore centrali della giornata in corrispondenza del max di densità
fogliare, mentre sotto la temperatura decresce.
Nella fig 4.17 (c) si osserva un profilo della pressione di vapore costante durante le ore
notturne, mentre durante le ore centrali della giornata abbiamo un max di pressione di
vapore in corrispondenza del max di densità fogliare dove avviene la maggior parte
dell’evapotraspirazione
Nella fig 4.17 (d) si osserva che durante le ore notturne si ha in prevalenza respirazione
e quindi concentrazioni elevate di CO2.
Mentre durante le ore centrali della giornata si ha un minimo di concentrazione di CO2
in corrispondenza dello strato fogliare dove avviene la fotosintesi.
Efficienza di intercettazione delle precipitazioni attraverso la canopy
Sono state prese in esame due tipi di foreste: una foresta tropicale e una temperata.
E’ stata misurata l’efficienza di intercettazione delle piogge dopo un temporale.
Nella fig 4.19 si nota che la massima capacità di intercettazione della pioggia, per tutti i
tipi di foreste, è fra gli 0.2 mm e i 2mm.
L’efficienza di intercettazione delle piogge moderate (10- 30 mm) è migliore per le
foreste tropicali.
Le foreste decidue intercettano fra il 10% e il 25% delle precipitazioni totali annuali, per
le conifere il 15- 40 %.
Modello schematico di scambi radiativi sopra e sotto una foresta
Il bilancio radiativo delle foreste dipende soprattutto dallo strato della canopy, mentre la
zona del fusto è meno importante.
La penetrazione della radiazione dipende dall’altezza, dalla densità della canopy e
dall’angolo di incidenza solare.
Dalla fig 4.20 si nota che solo il 20 % del flusso 1 raggiunge il terreno.
Una parte dell’energia incidente a onde corte viene riflessa dalla canopy (flusso 5), una
parte viene trasmessa ( flusso 2), una parte subisce riflessioni multiple ( flusso 3 e 4),
una ultima parte attraversa la canopy e viene riemessa nello spazio ( flusso 6).
Per le onde lunghe abbiamo una radiazione in entrata (flusso 8), una parte viene
trasmessa (flusso 9) al suolo.
Anche la canopy emette (flusso 10) verso il suolo 9+10=11).
Abbiamo poi una radiazione uscente dal suolo, emissione del terreno (flusso 12) che
viene trasmessa attraverso la canopy ( flusso 13).
Anche in questo caso la canopy emette verso lo spazio (flusso 14 ).
Relazione tra la radiazione a onde corte entrante sopra la canopy e sul
terreno durante il giorno
Dalla fig. 4.21 si può notare come si abbia un massimo di radiazione a onde corte
entrante a metà giornata.
La radiazione a onde corte sul terreno mostra una serie di assorbimenti.
Infatti la luce che arriva al terreno è depauperata delle regioni blu e rosse dello spettro.
Questo accade perché le foglie attuano la fotosintesi.
La luce che arriva al terreno è quindi fotosinteticamente meno attiva e questo costituisce
un limite alle piante di sottobosco.
Il bilancio radiativo a onde corte di un aranceto e di un singolo strato
di foglie di un arancio
Nella fig.4.22 (a) è stato preso in considerazione un aranceto.
Si nota che il 15% della radiazione a onde corte entrante viene riflesso dalla parte alta
della canopy dell’aranceto, il 21% viene trasmesso al suolo e di questo il 4% viene
riflesso dal suolo.
Di questo 4%, il 3% viene assorbito dalla canopy.
In totale nella canopy rimane una quota considerevole della radiazione entrante, il 67%.
Nella fig 4.22 (b) il discorso è analogo applicato però ad un singolo strato di foglie.
Variazione diurna del flusso radiativo di una foresta di conifere
Dalla fig. 4.23 si può notare come il massimo di radiazione a onde corte entranti si
abbia durante le ore centrali della giornata a cui corrisponde un massimo di riflessione a
onde corte.
Il bilancio energetico totale è positivo.
Per quanto riguarda le onde lunghe, per quelle entranti si evidenzia un profilo costante
durante la giornata, mentre per il flusso uscente abbiamo un massimo in corrispondenza
delle ore notturne.
Relazione del bilancio energetico diurno in 2 siti:
conifere ( Thetford) e abeti ( Haney)
Dalla fig. 4.24 si può notare come il flusso di calore sensibile nel sito di Thetford sia più
importante del flusso di calore latente.
Nel sito di Haney invece il flusso di calore latente è maggiore del flusso di calore
sensibile.
Per evidenziare queste differenze dobbiamo considerare due importanti fattori: la
resistività della canopy e la resistenza aerodinamica.
Nella fig. 4.25 si nota che la resistività della canopy nel sito di Thetford risulta essere
maggiore di quella di Haney.
Poiché la resistività della canopy influenza l’attività stomatica, in Thetford
l’evapotraspirazione è minore.
Per quanto riguarda la resistenza aerodinamica, questa è maggiore per Haney (legata
alla stagionalità dei venti).
Quindi la cessione di calore sensibile per Haney è minore.
Profilo verticale di alcune proprietà climatiche in una foresta di abeti
Quando entriamo in una foresta di giorno, il clima varia notevolmente e risulta essere
più fresco ed umido poiché ci troviamo sotto il livello della superficie attiva che assorbe
la radiazione e dove è collocata l’evapotraspirazione.
Si può notare dalla fig.4.26 come durante il giorno si abbia un max di temperatura e un
max di umidità dove è massima la densità fogliare.
Infatti qui nono solo si ha assorbimento della gran parte della radiazione ma anche una
forte traspirazione.
La canopy è quindi più calda del suolo di giorno; la notte il profilo della temperatura è
inverso: si hanno temperature più fredde nella parte superiore della canopy e più calde
nella parte sottostante poiché la canopy ostacola l’emissione delle onde lunghe.
Si nota anche una drastica riduzione della velocità del vento all’interno della canopy.
Per la CO2 abbiamo una elevata concentrazione a livello del suolo ( respirazione) ,
mentre la sua concentrazione è minima a livello della canopy ( fotosintesi).
Perdita e utilizzo d’acqua all’interno di sistemi vegetali (foresta e
campo coltivato)
•
Condizioni secche
In condizioni secche la perdita di acqua avviene soprattutto per traspirazione.
Osservando l’equazione 1 poiché la temperatura della canopy in un campo
coltivato è molto maggiore della temperatura ambientale, il numeratore di E crop
è maggiore del numeratore di E forest e quindi la traspirazione dei raccolti è
maggiore della traspirazione delle foreste.
•
Condizioni umide
In condizioni umide la perdita di acqua avviene soprattutto per evaporazione.
Osservando l’equazione 2 poiché la resistenza aerodinamica al vapore per le
foreste è minore rispetto a quella dei campi coltivati, la perdita di acqua per
evaporazione delle foreste è maggiore rispetto a quella dei campi coltivati.