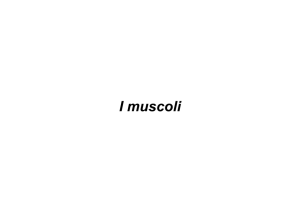Corso di Laurea Magistrale in
“Medicina e Chirurgia”
Fisiologia e Biofisica
A.A. 2015-2016
Contrazione muscolare
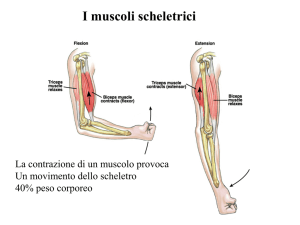
I muscoli hanno due funzioni: generare movimento
generare forza
I muscoli scheletrici generano anche calore e contribuiscono
alla termoregolazione (in ambiente freddo, il cervello ordina ai
muscoli di contrarsi e questo origina il brivido, che crea calore)
Energia chimica
muscolo
Energia meccanica
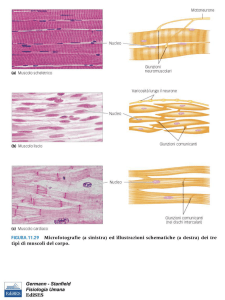
Le fibre muscolari scheletriche appaiono striate e
multinucleate. Controllano i movimenti del corpo.
Sono volontari, si contraggono in risposta
a stimoli che provengono da motoneuroni.
Le fibre muscolari cardiache sono uninucleate e striate.
Si connettono tra loro attraverso dischi intercalari.
Sono involontarie, si contraggono spontaneamente
senza stimoli esterni. L’attività è modulata dal
sistema nervoso viscerale e anche dal sistema
endocrino.
Muscolo liscio è il tipo principale di muscolo degli organi
interni e cavi, come stomaco, vescica, vasi sanguigni.
Cellule uninucleate senza evidenti striature.
Muscolo scheletrico
Connettivo
Fibre
muscolari
Vasi sanguigni
Nervi
Reticolo
sarcoplasmatico
Miofibrille
Sarcolemma
Tubuli T
Nuclei
Sarcoplasma
Mitocondri
Glicogeno
Fibra (cellula) muscolare
Tubuli T
Reticolo
sarcoplasmatico
Sarcolemma
Mitocondri
Nucleo
Filamento spesso
Filamento sottile
Miofibrilla
FILAMENTI SPESSI
2 catene pesanti (heavy chains)
p.m. 200.000
Miosina:
proteina oligomerica
p.m. 450.000
2 coppie di catene leggere
(light chains) non identiche
p.m.16.000 e 20.000
Dominio C-terminale: α-elicoidale
lunghezza 134 nm, diametro 2 nm
(coda della miosina)
Ciascuna catena pesante
Dominio N-terminale: globulare
(testa della miosina)
Coda
Ogni coppia di catene leggere è legata al dominio globulare di ciascuna
catena pesante nel tratto che connette testa e coda.
Snodo
Testa
Assemblaggio dei filamenti spessi nelle miofibrille
Nel muscolo scheletrico circa 250 molecole di miosina si uniscono a formare un
filamento spesso, il quale è sistemato in modo che le teste di miosina si raggruppano
all’estremità, mentre la regione centrale è un fascio di code di miosina
Tropomiosina: dimero a forma di bacchetta (pm = 70.000).
Le due subunità sono avvolte l’una intorno all’altra a formare un’elica.
Ogni molecola è lunga circa 40 nm.
I dimeri di tropomiosina si dispongono in sequenza con disposizione
testa-coda, formando un filamento elicoidale.
Due filamenti elicoidali si estendono per l’intera lunghezza del polimero
actina F.
Troponina
TnT
Legame
Tn-tropomiosina
TnI
Legame
Actina-TnC
TnC
Legame
Ioni Ca++
I filamenti spessi e sottili sono connessi da ponti trasversali, detti anche ponti crociati, costituiti dalle
teste di miosina che si legano lassamente ai filamenti di actina. Ogni molecola di G-actina ha un
singolo sito di legame per una testa di miosina.
Dischi Z: strutture proteiche a zig zag, che fungono da sito di attacco per i filamenti sottili. Un
sarcomero è composto da due dischi Z e dai filamenti tra essi compresi
Linea M: questa banda rappresenta il sito di attacco dei filamenti spessi
linea M
disco Z
disco Z
SARCOMERO
La titina è una enorme proteina elastica che occupa tutta la distanza tra disco Z e linea
M. Ha la funzione di agevolare il ritorno del sarcomero allungato alla sua lunghezza di
riposo; inoltre stabilizza la disposizione dei filamenti all’interno del sarcomero con l’aiuto
della proteina non elastica nebulina. La nebulina si trova di fianco ai filamenti sottili e si
attacca ai dischi Z. Garantisce l’allineamento dei filamenti di actina del sarcomero.
TEORIA DELLO SCORRIMENTO DEI FILAMENTI
Quando un sarcomero si contrae, i filamenti sottili e spessi non cambiano in lunghezza. Il
filamento sottile di actina scivola sul filamento spesso di miosina, spostandosi verso la linea M al
centro del sarcomero. La banda A non si modifica in lunghezza, ma sia la zona H che la banda I si
accorciano mentre i filamenti si sovrappongono.
La forza che spinge il filamento di actina è il movimento dei ponti crociati di miosina che legano
actina e miosina. La miosina è una proteina motrice che converte il legame chimico dell’ATP in
energia meccanica.
Da cosa è regolato questo processo? Nello stato rilassato la tropomiosina blocca parzialmente i siti
di legame dell’actina per la miosina.
La contrazione viene iniziata quando il Ca2+ si lega alla troponina C. Il legame col Ca2+ cambia la
conformazione della molecola di tropomiosina e scopre il resto del sito di legame per la miosina, così che
questa può completare la propria flessione.
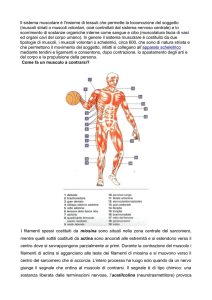
I segnali per la contrazione muscolare arrivano dal SNC ai muscoli scheletrici per mezzo dei
motoneuroni. L’acetlcolina proveniente dal motoneurone innesca un potenziale d’azione nella fibra
muscolare che a sua volta scatena una contrazione. Questa combinazione di eventi elettrici e meccanici
viene definito accoppiamento eccitazione-contrazione.
Il reticolo sarcoplasmatico avvolge ogni singola miofibrilla. Il sistema dei tubuli T è strettamente
associato al reticolo sarcoplasmatico ed è in continuità con la membrana di superficie della fibra
muscolare. I tubuli T permettono ai potenziali d’azione che originano sulla superficie cellulare a
livello della giunzione neuromuscolare di passare velocemente all’interno della fibra. Senza tubuli
T, il potenziale d’azione potrebbe raggiungere il centro della fibra solo per diffusione di cariche
positive nel citosol, processo più lento che ritarderebbe il tempo di risposta della fibra muscolare.
Quando il potenziale d’azione raggiunge un recettore diidropiridina (DHP), la connessione
meccanica fra il recettore e il reticolo sarcoplasmatico apre i canali per il Ca2+ .
L’apertura di questi canali rilascia Ca2+ nel citoplasma, dove gli ioni si possono combinare con la troponina
e permettere di iniziare la contrazione.
Il rilasciamento muscolare si ha quando il reticolo sarcoplasmatico riassorbe Ca2+ per mezzo di una Ca2+ATPasi. Quando la concentrazione citoplasmatica di Ca2+ scende, lo ione si stacca dalla troponina, la
tropomiosina scivola all’indietro per bloccare il sito legante la miosina, e la fibra si rilascia.
1 1. Il potenziale d’azione nel motoneurone somatico raggiunge il terminale assonale.
2+
2+
. 2. I canali del Ca voltaggio-dipendente si aprono. L’ingresso di Ca innesca l’esocitosi delle vescicole sinaptiche
contenenti ACh.
3. L’ACh diffonde nello spazio sinaptico e si lega ai recettori nicotinici sulla placca motrice del muscolo.
4. Il legame dell’ACh apre un canale cationico non specifico. Sia l’Na+ che il K+ si muovono attraverso il canale
secondo il loro gradiente elettrochimico. L’ingresso netto di cariche positive depolarizza la membrana
muscolare, generando un potenziale di placca.
5. Il potenziale di placca è sempre sovrasoglia e determina un potenziale di azione nella fibra muscolare.
6. Il potenziale di azione generato alla giunzione neuromuscolare diffonde lungo la membrana della fibra
muscolare, muovendosi verso l’interno della fibra tramite i tubuli T.
7. Il potenziale di azione nei tubuli T attiva i recettori diidropiridina. I recettori DHP aprono i canali del Ca2+
nella membrana del reticolo sarcoplasmatico.
8. Il Ca2+ diffonde fuori dal reticolo sarcoplasmatico e si lega alla troponina, allontanando la tropomiosina dal
sito di legame per la miosina. Questa azione permette alle teste della miosina di rilasciare il fosfato
inorganico dall’idrolisi dell’ATP e di completare la loro flessione.
9. Al termine della flessione, il ponte trasversale della miosina rilascia ADP e resta legato fortemente all’actina.
La miosina deve legarsi a una molecola di ATP per uscire da questo stato di rigor.
10. L’ATPasi della miosina idrolizza l’ATP in ADP e P, che restano legati alla testa della miosina. La miosina
torna indietro e si lega ad una nuova molecola di actina, pronta a eseguire la successiva flessione.
11. La fibra muscolare si rilassa quando il Ca2+ viene rilasciato dalla troponina e la tropomiosina torna a bloccare
il sito di legame della miosina. Il Calcio viene ritrasportato dentro al reticolo sarcoplasmatico tramite una
Ca2+-ATPasi.
Nicotina: agonista dei recettori colinergici nicotinici (recettori ionotropi)
presenti sulla membrana post-sinaptica della giunzione
neuromuscolare.
Agonista: sostanza che mima l’attività di un ligando.
Legame nicotina-recettore colinergico: determina apertura dei canali ionici
delle cellule post-sinaptiche, che si
depolarizzano.
Sebbene la nicotina sia un agonista, l’esposizione a lungo termine determina
inattivazione dei recettori colinergici con conseguente up-regulation degli
stessi.
La deprivazione da nicotina consente a tutti i recettori di ritornare allo stato
attivo: il maggior numero di recettori è probabilmente responsabile dei sintomi
da astinenza.
MIASTENIA GRAVE: malattia autoimmune in cui l’organismo non
riconosce i recettori colinergici come “self”.
Gli anticorpi anti-recettore determinano una
notevole riduzione dei recettori per l’acetilcolina. Ne consegue che, anche se il rilascio
del neurotrasmettitore è normale, la risposta
è ridotta e si manifesta debolezza muscolare.
SINDROME DI LAMBERT-EATON
In questa patologia, la debolezza muscolare è causata da un attacco
autoimmune nei confronti dei canali del calcio presenti sulla membrana
presinaptica. Ciò diminuisce il flusso di ioni calcio diretti verso l’interno
della cellula limitando la fusione delle vescicole con la membrana
e, quindi, la liberazione di acetilcolina.
Stimolazioni ripetute del motoneurone facilitano l’accumulo di calcio
nel bottone sinaptico e facilitano la liberazione del
neurotrasmettitore
PARALISI PERIODICA IPERKALEMICA
Difetto genetico dei canali voltaggio dipendenti del sodio che
non si inattivano. La fibra muscolare non si ripolarizza e non
genera un nuovo potenziale d’azione. Si ha accumulo di ioni
potassio nel LEC. La somministrazione di glucosio aiuta
a superare la paralisi.
I grafici a destra mostrano i potenziali d’azione nel terminale assonale e nella fibra muscolare,
seguiti dalla curva di tensione della scossa muscolare. Il periodo di latenza rappresenta il tempo
necessario perché il Ca2+ venga rilasciato dal reticolo sarcoplasmatico e diffonda verso i filamenti
della miofibrilla.
Un singolo potenziale d’azione evoca una
singola scossa muscolare.
Le scosse sono diverse da fibra a fibra
per la velocità con cui sviluppano tensione
(fase ascendente della curva della scossa),
la tensione massima raggiunta (altezza della curva)
e durata della scossa (ampiezza della curva).
T<C
Contrazioni isotoniche (T = C)
Muscolo a riposo
Contrazione isometrica:il muscolo non si accorcia.
I sarcomeri si accorciano, generando forza, ma gli
elementi elastici si allungono, così che la lunghezza
Muscolare resta costante
Contrazione fibre muscolari
scheletriche
Forza (Tensione)
Movimento
Resistenza ad un
carico
La tensione generata da un muscolo è direttamente proporzionale
all’interazione tra filamenti spessi e filamenti sottili.
La tensione sviluppata dalla contrazione dipende direttamente dalla lunghezza dei sarcomeri
prima che la contrazione inizi. Ogni sarcomero si contrarrà con il massimo della forza se è alla
lunghezza ottimale, né troppo lungo né troppo corto.
La teoria dello scorrimento dei filamenti prevede che la tensione che la fibra muscolare può
generare sia direttamente proporzionale al numero di ponti trasversali che si formano tra
filamenti spessi e filamenti sottili. Se la fibra comincia la contrazione quando il sarcomero è
troppo lungo, i filamenti sottili e spessi sono scarsamente sovrapposti e formano pochi ponti
trasversali quindi non possono generare molta forza. Viceversa, se il sarcomero è più corto della
sua lunghezza ottimale, i filamenti saranno troppo sovrapposti per cui i filamenti spessi possono
spostare i filamenti sottili solo per una breve distanza. Inoltre, i filamenti spessi finiscono a
contatto coi dischi Z e non possono formare ponti trasversali per cui la tensione cala rapidamente.
Adenosintrifosfato (ATP)
Fosfato
Adenina
Ribosio
Legame ricco
d’energia
La contrazione del muscolo scheletrico dipende dal rifornimento continuo di ATP. La quantità di
ATP presente nel muscolo è sufficiente per otto contrazioni circa. Come fonte di energia di
riserva, il muscolo contiene fosfocreatina, una molecola i cui legami fosfato ad alta energia
vengono generati a partire da creatina e ATP quando i muscoli sono a riposo
Debito di O2
Durante il lavoro muscolare, i vasi sanguigni che irrorano il muscolo
si dilatano aumentando l’apporto di ossigeno. In una prima fase,
l’energia è fornita da processi aerobici ed il
consumo di O2 è proporzionale al lavoro svolto.
Se il lavoro da compiere è molto grande, la sintesi aerobica delle
riserve energetiche non è più sufficiente e l’energia è fornita
da processi anaerobici.
Al termine dello sforzo, è necessario utilizzare una quantità di O2
supplementare per rimuovere l’eccesso di acido lattico e ripristinare
i depositi di ATP e fosfocreatina. L’extraconsumo di O2 è tanto
maggiore quanto più lungo è stato il ricorso a processi anaerobici.
Produzione di calore nei muscoli
W, lavoro meccanico
Energia fornita = Energia liberata
Sintesi ATP e PC
Q, calore
Rendimento = W/Energia fornita
0≤ Rendimento ≤50%
Contrazione isometrica
Contrazione isometrica
Calore attivazione
Calore iniziale
Calore accorciamento
Q
Calore ristoro
Calore di riposo
Processi metabolici di
base
Calore di riposo
Start
Stop
Tempo
La forza della contrazione aumenta con la sommazione delle scosse muscolari.
Una fibra muscolare risponde ad uno stimolo (▲
▲) con una scossa. Se gli stimoli sono
separati nel tempo, il muscolo si rilassa completamente tra le scosse.
La durata della scossa è inferiore al tempo intercorrente tra
due stimolazioni successive.
Quando gli stimoli sono ravvicinati nel tempo, la fibra muscolare non ha il tempo di
rilassarsi e le contrazioni si sommano, generando una contrazione con una tensione
maggiore.
La tensione generata da un muscolo aumenta se
si sommano più scosse singole ravvicinate
Se gli stimoli arrivano molto rapidamente, il muscolo raggiunge la sua massima
tensione. Se il muscolo ha ancora la possibilità di rilassarsi tra gli stimoli, si
realizza il tetano incompleto.
Se il muscolo raggiunge una tensione costante, è nella condizione di tetano completo.
Tetano completo
E’ quindi possibile aumentare la tensione sviluppata da una singola fibra muscolare
variando la frequenza dei potenziali d’azione nella fibra, che sono scatenati dal
rilascio di acetilcolina da parte dei motoneuroni.
CARATTERISTICHE DEI VARI TIPI DI FIBRE MUSCOLARI
Ossidative lente
muscolo rosso (I)
Velocità
contrazione
Ossidative rapide
muscolo rosso (IIa)
Glicolitiche rapide
muscolo bianco (IIb)
molto lenta
intermedia
Attività
ATPasica
lenta
rapida
molto rapida
Diametro
piccolo
medio
grande
Durata
contrazione
molto lunga
lunga
breve
Metabolismo
ossidativo
Colore
rosso scuro
glicolitico/ossidativo
rosso
molto rapida
glicolitico
pallido
Fibre muscolari
ossidative lente
Fibre muscolari
glicolitiche rapide
La grande quantità di mioglobina , numerosi mitocondri(M) e un’estesa
rete di capillari sanguigni (cap) distinguono il muscolo ossiodativo a lenta
contrazione (R) dal muscolo glicolitico a contrazione rapida (W).
Unità motoria: gruppo di fibre
innervate da un solo motoneurone
Unità motoria: è l’unità fondamentale della contrazione nel muscolo
scheletrico
Fibre dello stesso tipo.
La contrazione di un’unità è
un evento tutto o nulla.
Il numero di fibre muscolari in un’unità motoria è
variabile. Nei muscoli che governano i movimenti fini,
come quelli oculari o della mano, un’unità motoria può
contenere anche solo 3-5 fibre. Se l’unità motoria viene
attivata, solo poche fibre si contraggono e la risposta
muscolare è piccola. Se vengono attivate altre unità, la
risposta aumenta di poco perché si aggiungono solo poche
fibre. Ciò consente graduazioni fini nei movimenti.
Nei muscoli usati per movimenti grossolani, come stare in
piedi o camminare, un singolo motoneurone può
innervare centinaia di fibre muscolari.
Il muscolo può generare contrazioni graduate di varia forza e durata in quanto è
composto da molte unità motorie di diversi tipi. Questa varietà gli permette di variare
tipo di contrazione cambiando i tipi di unità motorie in attività e il numero di unità che
rispondono in un certo tempo.
Muscolo : insieme
di unità motorie.
La tensione sviluppata da un muscolo è in relazione al numero e al tipo di unità
motorie reclutate.
S (slow)
Tipo I
FR(fast resistent)
Tipo IIa
FR(fast fatigable)
Tipo IIb
Recettori sensoriali per il coordinamento dell’attività
motoria
Fuso neuromuscolare
Fibre
extrafusali
Fibre
intrafusali
Recettori sensoriali per il coordinamento dell’attività
motoria
AFFATICAMENTO MUSCOLARE