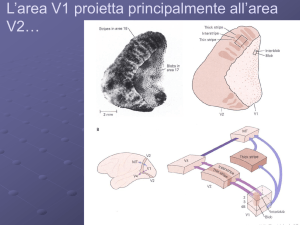
NEUROSCIENZE
Luce e visione
di Maria Concetta Morrone*
Lo studio del processo visivo ha consentito scoperte affascinanti su ciò che accade nell’occhio e nel cervello colpiti dalla luce;
ad esempio l’eliminazione delle informazioni sull’assorbimento di luce della retina. Un paradosso che ha alcuni vantaggi,
come quello di aumentare la capacità di discriminare piccolissime variazioni e quello di aumentare l’ambito di intensità
luminosa in cui la visione opera. E uno svantaggio: costringe il cervello a ricostruire la luminosità e il colore delle superfici che
compongono le immagini avendo solo frammentarie informazioni
L
a luce, così come il cervello la
percepisce, è un’illusione e non corrisponde
alla realtà fisica. Il processo visivo comincia
quando i fotoni, dopo aver attraversato i vari
stadi ottici dell’occhio e gli strati nervosi che
compongono la retina, vengono assorbiti dai
fotorecettori; eppure la quantità assorbita di
luce, sia quella mediata su tutto l’ambiente e
sia quella mediata all’interno di piccole
regioni, è una delle prime informazioni
filtrate via dai processi
visivi. I vantaggi
biologici
nell’eliminare
l’informazione su quanta luce
costantemente sia assorbita dalla
retina sono due: aumentare la
capacità di discriminare
piccolissime variazioni,
arricchendo
notevolmente la
sensibilità visiva, e
aumentare l’ambito di intensità
luminosa in cui la visione opera fino ad
estenderlo oltre un fattore di 1011,
ovvero dalla illuminazione di una notte
senza luna a quella di una spiaggia
assolata. Il cervello, non codificando
direttamente i valori della luce
dell’ambiente e quelli interni alle
superfici che compongono le
immagini del mondo esterno, deve
ricostruire la luminosità
(tecnicamente la “brillanza”) ed
il colore da poche e
frammentarie informazioni
proveniente dai
contorni delle
superfici, ma
nonostante ciò
deve assolvere questo
compito con estrema
affidabilità.
Per convincersi di quanto
illusoria sia la percezione della
luminosità bastano poche
osservazioni. Il cerchio della
Figura 1b non esiste: esso è
delimitato da un piccolissimo
contorno il cui profilo di luminanza
(proporzionale al numero di fotoni
per unità di superficie) è raffigurato
nel tracciato in basso. I valori di
luminanza delle superficie interne ed
esterne al cerchio sono identici, eppure si
Il trionfo dell’Eucarestia, La Stanza della
Segnatura, affresco, Musei Vaticani, Città del
Vaticano, (particolare).
40
NEUROSCIENZE
percepisce un disco più chiaro dello sfondo ed
omogeneo; ciò significa che una informazione
relativa al solo perimetro (il contorno) crea la
percezione di una superficie e di luce
(O’Brian, 1958), (Craik, 1966), (Cornsweet,
1970). Il fenomeno è noto ed usato nell’arte
grafica da tempo: le montagne della stampa
cinese (Figura 1a) sono analogamente
illusorie. Se si copre il perimetro, la chiarezza
della superficie del disco o delle montagne
svanisce. Non solo la sensazione di chiaro o di
scuro è illusoria, ma anche il colore. La
Figura 1c illustra un effetto simile a quello
precedente ma in questo caso è la qualità
della luce (o cromaticità) che cambia, e non la
luminanza. Il contorno è modulato in rossoverde, l’interno e l’esterno del cerchio sono di
colore giallo eppure la superficie interna
appare verde mentre quella esterna tende al
rosso. La percezione non coincide con la realtà
fisica, è una illusione e quindi un errore che il
cervello compie; occasionalmente questo potrà
comportare uno svantaggio (pensate di voler
sollevare la superficie del disco con una
pinza), tuttavia queste illusioni sono una spia
dei processi e dei meccanismi nervosi che
consentono la percezione della scena visiva a
partire dall’informazione fisica contenuta
nella luce.
Se la quantità di luce media dell’ambiente e
all’interno di piccole superfici non viene
codificata dalla retina, e quindi non è
trasmessa al cervello, quale è l’informazione
che ci permette di ricostruirla? L’informazione
cruciale è il contrasto, ovvero un parametro
che misura la variazione nello spazio della
luminanza, e non il suo valore medio in una
regione. Questa è un’intuizione abbastanza
ragionevole: per definizione un oggetto è
associato ad un contorno e quindi ad una
variazione di luminanza o contrasto. L’analisi
del contrasto e la ricostruzione da esso della
brillanza (luminanza apparente) impone una
costanza percettiva della superficie che è
indipendente dal livello di luce reale. È un
intuizione che già Hering aveva avuto e che lo
portò ad analizzare le condizioni di costanza
della brillanza: “Senza questa costanza seppur
parziale, un pezzo di gesso in un giorno
nuvoloso dovrebbe manifestarsi come un
pezzo di carbone in un giorno di sole, e nel
corso del giorno dovrebbe assumere tutti i
possibili colori tra il nero e il bianco” (Hering,
1878), traduzione (Fiorentini, Baumgartner et
al., 1990)). L’intuito di Hering è andato oltre,
ipotizzando che il bianco e il nero percettivo
fossero il risultato dell’azione di due
meccanismi antagonisti; e successivamente, il
fisico Mach, con un’altra brillante intuizione,
associò questi meccanismi “ad una operazione
simile alla derivata seconda o Laplaciano”
(Mach, 1906), anticipando di quasi un secolo
le moderne teorie dell’analisi del contrasto e
della forma.
Una dimostrazione che il parametro
importante per la codifica della brillanza è la
differenza tra la luminanza dello sfondo e
dell’oggetto è fornita dalla Figura 2a. I
quadratini centrali hanno tutti la stessa
luminanza, mentre lo sfondo varia in
luminanza. Si nota che, più lo sfondo è scuro,
più chiaro appare il quadratino e viceversa.
Le conoscenze attuali delle proprietà del
sistema visivo permettono di capire questo
fenomeno ed anche di simularne
quantitativamente gli effetti: come aveva
intuito Mach, l’operazione cruciale è la
differenziazione della luminanza che avviene
grazie alle proprietà dei neuroni visivi, in
particolare grazie alla struttura del campo
recettivo. Uno dei principi cardini della
fisiologia sensoriale è il concetto di campo
recettivo: per la visione questo coincide con
l’area di superficie retinica che, stimolata,
produce un’alterazione della scarica del
neurone. Già dopo la prima sinapsi nella
retina, i campi recettivi dei neuroni visivi non
sono omogenei ma strutturati in regioni
concentriche antagoniste eccitatorie e
inibitorie; così i neuroni codificano
ottimalmente un’area scura su fondo bianco
(centro Off) oppure un’area bianca su fondo
scuro (centro On). L’azione antagonista della
periferia, una proprietà comune a molti
sistemi sensoriali, annulla la risposta ad uno
stimolo omogeneo per luminosità in tutto il
campo recettivo e crea una specializzazione
neuronale a variazioni locali di luminanza
come quelle associate a bordi e contorni; in
pratica l’antagonismo centro-periferia effettua
una operazione che corrisponde in termini
matematici ad una differenziazione. I neuroni
di centro On ed Off sono alla base della nostra
sensibilità al contrasto e della percezione
Figura 1a: Una stampa cinese di Pu Ru (1887-1963). La brillanza delle montagne è illusoria, se si copre il
contorno la differenza di luminanza apparente tra cielo e terra svanisce. L’effetto era noto agli artisti cinesi già
molti secoli prima, durante la dinastia Ming (1368-1644).
Figura 1b: Illusione di Craik-O’Brian e Corsweet. Il disco chiaro svanisce appena si occlude il contorno, (per
l’occlusione si può utilizzare un’apertura formata dal pollice ed dall’indice). La luminanza varia solo per una
piccolissima regione vicino al perimetro del disco eppure questa variazione è sufficiente per far apparire tutta la
regione all’interno del contorno più chiara. Il profilo di luminanza dell’immagine è illustrato dal tratto nero in
basso.
Figura 1c: L’analogo dell’illusione in B nel dominio del colore. La luminanza è costante in tutta l’immagine, il
bordo è definito solo da una piccola variazione di colore, come illustrato dal profilo in basso. Il colore all’interno
ed all’esterno del disco è giallo, eppure il disco viene percepito come verde.
41
NEUROSCIENZE
diversa della serie di quadratini della Figura
2a. Analogamente, altri neuroni, anche questi
a livello retinico, sono sensibili anche a
variazioni del colore in particolari direzioni,
avendo un antagonismo centro-periferia
selettivo alla lunghezza d’onda della luce.
Questi neuroni sono alla base dell’analisi del
contrasto cromatico dello stimolo e quindi
svolgono, per esempio, un ruolo essenziale per
la codifica dello stimolo in Figura 1c. Per
brevità l’argomento del colore non verrà qui
approfondito, ma è importante notare che
molti degli effetti percettivi e dei meccanismi
nervosi che verranno illustrati in seguito per
la percezione della brillanza hanno dei
corrispettivi analoghi nella percezione del
colore.
Negli esempi riportati fino a qui, la brillanza
della superficie è dettata dal salto brusco (o
“bordo”) fra due regioni estese di diversa
luminanza; in questi casi il contrasto è
definito come la semi-differenza tra le
luminanze delle due superfici normalizzata
alla luminanza media. Ma non sono solo i
bordi di contrasto a generare la percezione
della luce o ad essere importanti per la
codifica delle informazioni visive. La Figura
2b illustra la famosa illusione di Chevreul, un
famoso chimico francese che notò per primo
questo effetto percettivo mentre sintetizzava
coloranti per tessuti (Chevreul, 1890):
sebbene ogni gradino sia di luminanza
costante, la scalinata viene appena percepita e
ogni gradino appare concavo, in accordo con
un meccanismo di differenziazione. Tuttavia
se si sommano al gradino sbarre sottili,
vengono a formarsi superficie più scure che
globalmente vengono percepite illusoriamente
come triangoli; se si occludono le linee, i
triangoli spariscono. Anche questo è un
fenomeno di contorno o perimetro, ma non è
consistente con la semplice idea di
differenziazione effettuata dal meccanismo
antagonista centro-periferia dei neuroni visivi
e richiede un ulteriore approfondimento del
problema.
Ogni scena naturale contiene un numero
elevatissimo di profili di luminanza associati a
Figura 2a: Effetto di contrasto simultaneo: un oggetto di riflettanza moderata può apparire più chiaro o più
scuro se esso è circondato da una regione molto più scura o chiara dell’oggetto stesso. I quadratini centrali hanno
tutti la stessa luminanza, ma quello in alto appare più scuro di quello in basso.
Figure 2b, c e d: Esempi dell’illusione di Chevreul. Come illustrato dal profilo in D, i gradini della scalinata
hanno la stessa altezza e sono piatti, ma sono percepiti concavi e di altezza variabile (A). Aggiungere delle linee
chiare all’interno di ogni gradino induce una differenza di brillanza tra le regioni che esse separano (C): i
triangoli scuri all’interno di ogni gradino sono illusori.
42
bordi, a linee o sbarre, a rampe e tutti questi a
diversi livelli di sfuocamento e di rumore.
Inoltre l’illuminazione con sorgenti multiple e
oblique porta spesso a creare profili ibridi che
si possono descrivere come una somma o la
combinazione di sbarre e di bordi. L’effetto
può essere cosi forte che punti di contatto fra
due superficie sono spesso associati a linee
piuttosto che bordi (si noti, per esempio, il
profilo del contatto tra il soffitto ed una
parete dello stesso colore; occludendo la zona
di contatto la differenza di brillanza fra le due
superficie sparisce).
È comune esperienza che l’informazione
contenuta nei bordi possa essere rappresentata
con semplici linee senza togliere alcuna
salienza cognitiva della scena. Linee e bordi
sono intercambiabili come continuamente ci
mostrano gli artisti (si pensi ad esempio ai
disegni di Picasso), ma i profili di linee e
bordi sono entità distinte e sono, in effetti,
matematicamente ortogonali (per dettagli si
veda la legenda della Figura 3). L’invarianza
percettiva tra linee e bordi è da attribuire
interamente alla funzionalità del nostro
sistema visivo e alla organizzazione dei
circuiti nervosi operanti nella corteccia visiva.
I campi recettivi dei neuroni visivi a livello
corticale sono molto più complessi che nella
retina ed oltre a codificare grandezza ed
orientamento dello stimolo, codificano anche
la forma d’onda utilizzando simmetrie di
campo opportune come quelle rappresentate
graficamente in Figura 3. Il campo recettivo
schematizzato in rosso nella Figura 3a è un
campo a simmetria centrale dispari, mentre
quello in blu è a simmetria pari: entrambi
rispondono a bordi o linee, ma è solo la loro
combinazione che univocamente determina la
posizione del bordo o della linea e il contrasto
percettivo del profilo (per dettagli si rimanda
a Morrone, Burr, 1988 e a Burr, Morrone,
1992. Un’altra famosa illusione, per prima
notata da Mach (Mach, 1865) e riscoperta e
studiata quantitativamente un secolo più tardi
da Adriana Fiorentini (Fiorentini, 1972), ci
aiuta a capire come il nostro cervello riesca a
percepire la struttura di scene visive reali ed
illusorie grazie alla combinazione delle
risposte di questi due tipi di neuroni. Bande
chiare o scure vengono ad essere
illusoriamente percepite dove si congiungono
NEUROSCIENZE
rampe di diversa pendenza, come illustrato
nella Figura 3: il profilo fisico trapezoidale
viene ad essere percepito come tre regioni di
brillanza costante, separate da bande molto
strette. Questa illusione è complementare
all’illusione della Figura 2c dove le linee
inducono la percezione illusoria di bordi:
l’onda trapezoidale dovrebbe produrre un
forte salto di luminanza o bordo nel punto in
cui la rampa raggiunge il punto medio, invece
la luminanza della rampa appare abbastanza
costante e, agli apici della rampa, dove
dovrebbero esserci bordi appaiono delle linee.
Non bisogna pensare che questi siano artifici
di laboratorio, percepiamo continuamente
bande di Mach, anche se esse passano spesso
inosservate: ad ogni profilo d’ombra sono
associate bande di Mach molto vivide e si
potrebbe ipotizzare che la loro presenza
indichi al nostro cervello che quel salto di
luminanza deve essere interpretato come
un’ombra e non come un oggetto. Anche i
neuroni con campi recettivi lineari (come
quelli in Figura 3a) segnalerebbero, con
picchi e valli, la presenza di bordi che non
vengono però percepiti (Figura 3b e 3c);
questo ha portato a non considerare solo le
risposte dei singoli neuroni ma anche le
combinazioni non lineari delle loro attività.
Un semplice modello (Morrone, Burr, 1988),
plausibile biologicamente e
computazionalmente poco complesso, che
effettua la somma pitagorica delle risposte di
neuroni con campi recettivi a simmetria
dispari e a simmetria pari riesce a simulare
quantitativamente la posizione dei segni
importanti di una immagine, come i bordi
delle Figure 1 e 2 e anche le bande di Mach. Il
modello simula anche la visibilità e la
percezione di questi segni (Morrone, Ross et
al., 1986; Ross, Morrone et al., 1989). Per
l’onda trapezoidale (Figura 3d), ad esempio,
la somma quadratica delle risposte dei campi
recettivi pari e dispari (detta Energia Locale)
produce solo due picchi (massimi locali) in
corrispondenza dei due segni più salienti
dell’immagine, ovvero delle bande di Mach; in
questi punti l’attività massima è prodotta
principalmente dai neuroni con simmetria di
campo pari, segnalando così la presenza di
linee; queste sono localizzate spazialmente
nella posizione in cui si percepiscono le bande
di Mach.
Questo modello riesce a predire con successo
la sensibilità al contrasto e la brillanza
apparente di numerose illusioni visive (quelle
mostrate in precedenza e altre); inoltre riesce
a catturare la struttura spaziale con cui il
sistema visivo organizza le varie immagini che
si formano sulla retina e per questo esso è
utilizzato in molti sistemi di visione artificiale
come meccanismo di segmentazione. Uno
degli esempi più chiari di come la struttura
dell’immagine sia molto spesso lontana dalla
struttura fisica è fornita dall’illusione “di
quantizzazione”, un metodo ormai
comunemente utilizzato in televisione e nella
stampa per impedire il riconoscimento di
immagini. Esso ha affascinato molti artisti, in
particolare Chuck Close, che lo utilizza con
squisita eleganza. Da vicino l’immagine in
Figura 4a sembra una elaborata composizione
astratta, vista da lontano emerge un viso.
Cosa impedisce al nostro cervello di utilizzare
l’informazione della faccia per
l’organizzazione dell’immagine vista da
vicino? Nella versione classica dell’illusione
(Harmon, Julesz, 1973), (Figura 4b)
l’immagine appare come una serie di blocchi
di brillanza costante, da lontano emerge il
volto di una bimba. Misure quantitative
dell’illusione, indicano che la base
Figura 3a: Esempio delle bande di Mach. L’onda ha un profilo trapezoidale, con una rampa limitata dai punti ?
e ?, come indicato in E. Vivide linee scure e chiare vengono percepite agli apici della rampa (? e ?). Sovrapposti
all’immagine sono rappresentati esempi di Campi Recettivi di neuroni corticali di aree visive primarie. L’area
tratteggiata e quella a puntini rappresentano le zone in cui la presentazione di uno stimolo chiaro (On) e di uno
stimolo scuro (Off) producono una risposta eccitatoria del neurone. Il campo recettivo in rosso ha una simmetria
di campo dispari (bi-partito) mentre quello in blu ha una simmetria di campo pari (tri-partito).
Figura 3b: Simulazione della risposta all’onda trapezoidale di una popolazione di neuroni con campi recettivi a
simmetria dispari e distribuiti omogeneamente nello spazio. Neuroni con campi recettivi posizionati interamente
sulle porzioni a luminanza costante sono silenti e mostrano una attività nulla. Neuroni in prossimità degli apici
della rampa mostrano una chiara risposta eccitatoria o inibitoria, indicando erroneamente la possibile presenza
di bordi di diversa polarità.
Figura 3c: Simulazione delle risposte di neuroni a simmetria di campi pari. La loro attività è massima e minima
in corrispondenza degli apici della rampa, indicando correttamente la presenza di linee a questi punti. Tuttavia
questa popolazione indicherebbe erroneamente la presenza di altre linee in corrispondenza degli altri massimi e
minimi relativi della risposta. Si noti come i massimi relativi della funzione corrispondano a punti di zero della
funzione in B e viceversa; le due funzioni infatti sono ortogonali nel dominio L2.
Figura 3d: Energia locale calcolata sommando i quadrati delle risposte dei campi recettivi a simmetria pari e
dispari (profili in B e in C). La funzione presenta solo due massimi e questi sono in corrispondenza degli apici
della rampa, dove sono percepite le bande di Mach.
43
NEUROSCIENZE
della visione che abbiamo, dotata di milioni e
milioni di sfumature e strutture. ●
*Istituto di Neuroscienze CNR di Pisa
Bibliografia
Burr D.C., Morrone M.C., “A Non-Linear Model of
Feature Detection”, in Non-Linear Vision, Nabet RBPaB,
pp. 309-328. CRC Press, Inc.,1992, pp. 309-328.
Figura 4a: Particolare di un ritratto dell’artista Chuck Close (Bill II, 1991). Da vicino l’immagine appare come una
elaborata composizione astratta, vista da lontano emerge un viso.
Chevreul M.E., “The Principles of Harmony and
Contrast of Colours”, (trad. di Martel C.) Bell, London,
1890.
Figura 4b: Illusione di “quantizzazione”: da vicino l’immagine appare una composizione più o meno simmetrica di
quadrati, da lontano emerge il volto di una bimba.
Cornsweet T.N., Visual Perception, Accademic Press,
New York, 1970.
Figura 4c: Questa immagine è stata ottenuta dall’immagine quantizzata in B trasformando i bordi di ogni
quadratino in linee. L’informazione importante per il riconoscimento del volto è identica in B e in C, infatti le due
immagini osservate da lontano appaiono uguali. Da vicino il volto della bimba è riconoscibile in C, ma non in B.
Craik K., The Nature of Psychology, Cambidge Univ.
Press, Cambridge, 1966.
dell’illusione stia nella congruenza tra
l’informazione dei bordi dei blocchi e
l’informazione sfuocata dell’immagine del
volto (Morrone, Burr et al. 1983). Se si altera
questa congruenza, trasformando ogni bordo
in una linea, questi nuovi segni perdono la
capacità di catturare l’informazione rilevante
al riconoscimento, e cosi il volto (Figura 4c)
può emergere in trasparenza dietro una griglia
(Morrone, Burr, 1997). Il modello dell’energia
locale misura esattamente questa congruenza
ed è in grado di predire quantitativamente
entrambe le percezioni della Figura 4. Esso
fornisce le condizioni necessarie di come e
quando una informazione al perimetro possa
propagarsi con costanza a tutta la superficie
che il contorno stesso racchiude, come succede
per l’illusione di Cornsweet e per ogni
quadrato della faccia quantizzata.
Il modello si basa su proprietà fisiologiche
note dei neuroni della corteccia visiva. I premi
Nobel Hubel e Wiesel (Hubel, Wiesel, 1962)
per primi hanno descritto campi recettivi
visivi con diversa simmetria (cellule pari e
dispari della Figura 3, da loro definite
Fiorentini A., Baumgartner G., Magnussen S., Schiller
P.H., Thomas J.P., The Perception of Brightness and
Darkness, Academic Press, San Diego, 1990.
Il trionfo dell’Eucarestia, La Stanza della Segnatura,
affresco, Musei Vaticani, Città del Vaticano,
(particolare). Appartengono al semicerchio celeste, a
partire da sinistra, San Pietro, Adamo, San Giovanni
Evangelista, David, Santo Stefano protomartire e
Geremia.
“semplici”) e campi recettivi omogenei ed
invarianti a stimoli On e a stimoli Off (cellule
definite “complesse”): entrambe queste
proprietà sono essenziali per implementare
biologicamente un’operazione simile a quella
richiesta dal modello di energia locale. Le
cellule complesse potrebbero effettuare la
somma pitagorica e le cellule semplici
potrebbero operare sia gli stadi iniziali di
filtraggio lineare sia l’operazione di
identificazione. I metodi moderni di Imaging
Funzionale hanno permesso recentemente di
convalidare queste ipotesi nell’uomo.
Utilizzando immagini come quelle della
Figura 1b e delle Figure 4b e 4c si è mostrato
che l’attività globale di aree visive primarie
non è differente per stimoli apparentemente
molto diversi, dotati però della stessa
struttura di Energia Locale; ovvero queste
aree corticali effettuano una analisi che molto
da vicino assomiglia al calcolo dell’ Energia
Locale. Solo ad un livello più avanzato
dell’analisi visiva, in una area non identificata
in precedenza, ma localizzata all’interno
dell’area associativa Broadman 19, si è
osservata una risposta che discrimina come
diversi due stimoli che hanno la stessa energia
locale, come ad esempio gli stimoli della
Figura 4b a e 4c. Questa area potrebbe
assolvere il difficile ma essenziale compito di
ricostruire la brillanza delle superficie da
poche e frammentarie informazioni relative al
contorno e quindi permetterci la ricchezza
45
Fiorentini A., “Mach Band Phenomena”, in Handbook
of Sensory Physiology, Vol VII/4, Huvich DjaLM,
Springer-Verlag, Berlin, 1972.
Harmon L.D., Julesz B. “Masking in Visual Recognition:
Effect of Two-Dimensional Filtered Noise”, in Science,
vol. 180, 1973, pp. 1194-1197.
Hering E., Zur Lehre vom Lichtsinn, Gerald und Söhne,
Wien, 1878.
Hubel D.H., Wiesel T.N., “Receptive Fields, Binocular
Interaction and Functional Architecture in the Cat’s
Visual Cortex”, in J. Physiol., (Lond.), vol. 160, 1962,
pp. 106-154.
Mach E, “Uber den Einfluss Raumlich und zeitlich
variierender Lichtreize auf die Gesichtswahrnehmung”,
in S.-B. Akad. Wiss., Wien, math.-, vol. 115, 1906, pp.
633-648.
Mach E. “Uber die Wirkung der raumlichen Vertheilung
des Lichreizes auf di Neztzhaut”, in I.S.-B. Akad. Wiss.,
Wien, math, vol. 54, 1865, pp. 303-322.
Morrone M.C., Burr D.C. “Capture and Transparency in
Coarse Quantized Images”, in Vision Research, vol. 37,
1997, pp. 2609-2629.
Morrone M.C., Burr D.C., “Feature Detection in Human
Vision: A Phase Dependent Energy Model”, in Proc. R.
Soc. (Lond), vol. B235, 1988, pp. 221-245.
Morrone M.C., Burr D.C., Ross J., “Added Noise
Restores Recognition of Coarse Quantised Images”, in
Nature, vol. 305, 1983, pp. 226-228.
Morrone M.C., Ross J., Burr D.C., Owens R., “Mach
Bands Depend on Spatial Phase” in Nature, vol. 324,
1986, pp. 250-253.
O’Brian V., “Contour Perception, Illusion and Reality”,
in J. Opt. Soc. Am., vol. 48, 1958, pp. 112-119.
Ross J., Morrone M.C., Burr D.C., “The Conditions for
the Appearance of Mach Bands”, in Vision Res., vol. 29,
1989, pp. 699-715.