Parte II
Le neuroscienze
2.1. PREMESSA
Grazie alle tecniche di neuroimaging, si dispone oggi di numerosi studi sperimentali in cui
vengono esplorate le funzioni di singoli neuroni, ma anche di cluster di neuroni, in corrispondenza a
compiti cognitivi complessi; a tali studi rivolgono l’attenzione ricercatori di diverse discipline,
come linguisti, pedagogisti, filosofi, e le possibili conseguenze dei diversi risultati iniziano ad
essere usate pervasivamente in molti campi. Sicché non è difficile immagina re le ripercussioni che
tali studi possono avere sulla comprensione dei processi linguistici e dell’apprendimento negli
esseri umani. Su questa base si può arrivare a ripensare teorie dell’apprendimento di natura
psicologica, basate sulla fenomenologia del comportamento, da quelle più classiche ad altre più
recenti (Piaget, Vygotskij, la Evolutionary Psychology1 , etc.), e ci sembra perciò ineludibile il
confronto anche nel campo della ricerca in didattica della matematica. Ma tutto ciò può essere
dispersivo e perfino pericoloso, se non si riescono ad inserire i vari contributi in un modello
coerente che li raccordi e li integri. Questo non vuol dire che ad oggi esista un tale modello globale,
ma in questa direzione sono stati comunque fatti negli ultimi anni molti passi avanti.
Presenteremo dunque in questa Parte Seconda alcuni risultati recenti di neuroscienze,
limitandoci a quanto riteniamo significativo rispetto ai nostri scopi. Non vogliamo discutere su che
cosa si intende in generale per neuroscienze: il termine è alquanto vago, troppo ampio, contiene
molte cose diverse. Sarebbe interessante certamente un excursus storico intorno a questa galassia,
ma non è nei nostri scopi e porterebbe troppo lontano.
È superfluo infine notare che non siamo esperti di alcuno degli specifici settori di ricerca in
tale ambito; così come sappiamo di non rivolgerci ad esperti di tale disciplina. Tuttavia entreremo,
nel limite del necessario, in alcune questioni tecniche che potranno creare qualche difficoltà di
comprensione per il linguaggio specialistico. Nello stesso tempo ci scusiamo per eventuali
superficialità ed imprecisioni terminologiche e disciplinari.
2.1.1. Gli apparati e gli strumenti sperimentali.
Le tecniche e apparati per le indagini sperimentali oggi disponibili, e in particolare le tecniche
di imaging cerebrale, realizzate sia su animali che su esseri umani, sono di svariati tipi. Non
sottolineeremo tuttavia nel seguito quali di queste tecniche sono di volta in volta impiegate, in
quanto questo aspetto non influenza le nostre argomentazioni. Tali metodi di indagine sono elencati
comunque in (Rizzolatti & Sinigaglia, 2006), pagg. 113-116, dove si fa riferimento alle tecniche più
classiche di elettroencefalografia (EEG), magnetoencefalografia (MEG), stimola zione magnetica
transcranica (TMS), e a quelle più recenti come la Tomografia a Emissione di Positroni (PET) e la
Functional Magnetic Resonance Imaging (fMTI). Una fonte molto recente e accessibile su tali
metodiche è anche l’articolo a carattere divulgativo (Rizzolatti et al., 2006), mentre un classico più
completo ma meno aggiornato è (Porter e Reichle, 1997).
Osserviamo, a proposito delle metodiche sperimentali, che finora la gran parte dei dati di cui
si dispone sono riferiti ad animali e in particolare ai primati, per i quali è possibile acquisire dati
direttamente sul comportamento di singoli neuroni, usando tecniche invasive; sulla base di tali dati
vengono formulate ipotesi sul cervello umano. La novità di questi ultimi anni è che tali ipotesi
possono essere sottoposte estensivamente a validazione con le moderne tecniche di brain imaging
funzionale, che vengono utilizzate con gli esseri umani.
1
Vedi, ad esempio (Cosmides & Tooby, 1997).
1
Segnaliamo qui anche una questione minore, su cui però val la pena di riflettere: secondo
Rizzolatti e Sinigaglia grandi passi avanti nella ricerca, in particolare la scoperta dei neuroni
specchio, si sono fatti adottando il paradigma sperimentale, che qui convenzionalmente chiameremo
della scimmia libera 2 . Si tratta di focalizzare l’attenzione non su singoli gesti predeterminati
dell’animale, ma su azioni complesse organizzate secondo un’intenzione. Del resto osservare un
animale (ma anche un essere umano) quanto più possibile nel suo ambiente naturale, alle prese con
la varietà e complessità di questo, è un’esigenza che già da molto tempo 3 viene sottolineata: in
polemica con procedure tipiche della psicologia sperimentale, dove l’obiettivo di osservare una
variabile molto specifica, isolata da tutte le altre, si persegue creando setting sperimentali artificiali,
in cui la complessità è ridotta a zero, con il risultato che quanto osservato difficilmente può ritenersi
poi significativo rispetto al comportamento reale. Se richiamiamo qui questo punto, è perché ci
sembra una metafora pertinente anche per la ricerca in didattica.
2.1.2. Le nostre fonti.
I lavori a cui ci si può riferire oggi sono in tumultuosa crescita. Sorgono un po’ ovunque centri
e progetti specifici di ricerca, si moltiplicano le riviste specializzate (cartacee e on-line), si
differenziano continuamente sottodiscipline, e si intrecciano variamente le vecchie. Tutto ciò anche
in Italia. In questo oceano in espansione, il nostro riferimento principale sono i lavori del
neurobiologo francese Jean-Pierre Changeux, che, oltre che in innumerevoli articoli, in due opere di
sintesi, L’uomo neuronale e L’uomo di verità, rispettivamente del 1983 e del 2002, tradotte
tempestivamente in italiano nel 1983 e nel 2003, (Changeux, 1983 e 2002), espone le linee generali
delle attuali conoscenze sul cervello ed una teoria generale sul suo funzionamento. Nel seguito
presentiamo le sue idee, che appaiono pertinenti al nostro lavoro, attingendo soprattutto al più
recente L’uomo di verità. Molto altro è stato fatto dopo il 2000, risultati di grande interesse che
spesso si riconnettono in larga misura con la teoria generale di Changeux. Faremo riferimento in
particolare alle recenti ricerche di Rizzolatti, Gallese e altri sui neuroni specchio (mirror neurons),
attingendo soprattutto da (Rizzolatti & Sinigaglia, 2006), (Gallese & Metzinger, 2003), (Gallese &
Lakoff, 2005). Tra le altre fonti di cui ci serviremo, segnaliamo (Levi-Montalcini, 1998) e
(Boncinelli, 2001).
2.2. LA MATERIA PENSANTE4
La teoria sviluppata da Jean-Pierre Changeux ci sembra configurarsi come l’anello mancante
tra, da un lato, l’enorme mole di dati sperimentali “locali” sull’attività neurale forniti dalle tecniche
di brain imaging cerebrale e quelli più classici di comportamenti cognitivi di pazienti con lesioni
cerebrali, e, dall’altro, la fenomenologia dei comportamenti cognitivi osservabili a livello “globale”.
Negli ultimi anni, grazie alle tecniche di imaging funzionale si sono sviluppate un gran numero di
ricerche in tal senso, basate su dati sperimentali 5 , che confermano e approfondiscono alc uni assunti
nodali della teoria di Changeux.
Riportiamo, in questa parte della nostra esposizione, quegli aspetti principali della teoria di
Changeux che più ci sembrano interessanti per i nostri scopi, proprio per i suoi caratteri di struttura
articolata e coerente, ma integrandoli con alcuni approfondimenti consentiti dalle ricerche più
recenti. L’idea che ci guida è che disporre dei corrispettivi neurali dei comportamenti cognitivi può
essere utile per ottenere dati sperimentali di tipo standard sull’apprendimento. Naturalmente il
2
Cfr. (Rizzolatti e Sinigaglia, 2006), pag. 24.
Vedi ad esempio (Neisser, 1976), pag. 52-53.
4
La locuzione è di Voltaire, citato in (Changeux, 2002), pag. 14.
5
Vedi ad esempio (Rizzolatti & Sinigaglia, 2006), (Gallese & Lakoff, 2005), (Gallese & Metzinger, 2003)
3
2
progetto è ambizioso e, come dice Changeux, molto lontano dall’essere compiuto; tuttavia, come
cercheremo di mostrare in seguito, già ora si può ad esempio asserire che alcuni costrutti di
Vygotskij trovano conferma nei modelli neurali sviluppati in base all’imaging funzionale: ci
riferiamo alla zona di sviluppo prossimale e allo spazio di lavoro neuronale.
2.2.1. Il cervello umano e la sua evoluzione
In (Changeux, 2002), l’Autore si prefigge l’ambizioso obiettivo di identificare le basi
fisiologiche delle “funzioni superiori” della mente, di natura sia cognitiva che affettivo-emotiva; la
cosa interessante è che il modello che propone per la struttura e il funzionamento del cervello è
integrato e sostenuto da un modello di sviluppo sia filogenetico che ontogenetico del cervello
stesso. Secondo Changeux, il cervello è un organo attivo e dinamico caratterizzato da una marcata
“plasticità strutturale”, plasmato da un’incessante interazione con l’ambiente e dalla sua stessa
dimensione autoriflessiva (sogni, pensieri, immaginazione…).
Il cervello è dunque il risultato di due percorsi integrati: da un lato è il prodotto
dell’evoluzione biologica per selezione naturale: un organo che ha scremato, più o meno attraverso
“centomila generazioni” di homo sapiens, rappresentazioni del mondo esterno sempre più efficaci e
vantaggiose per l’individuo e per la specie, rappresentazioni che vengono trasmesse e affinate sia
per via genetica che per via epigenetica6 ; nello stesso tempo il cervello è il prodotto
dell’embriogenesi e dello sviluppo individuale. In altri termini lo sviluppo del cervello chiama in
causa la doppia realtà di ogni individuo, le sue strutture biologiche, costituite da cellule, e il suo
genoma. Se le strutture biologiche cambiano, il genoma è pressoché immutabile, dato che
l’evoluzione, cui è sottoposto l’individuo stesso, è molto lenta e i cambiamenti avvengono nell’arco
di decine e centinaia di migliaia di anni. Le strutture cellulari invece nascono e scompaiono nel giro
di ore o di giorni e “vedono” il genoma da cui derivano. L’identità di ogni particolare organismo
risiede nel genoma, anche se le istruzioni in esso contenute devono essere lette e interpretate: a
questa lettura e a questa interpretazione provvede il complesso delle strutture biologiche, costituite
prevalentemente da proteine, presenti nell’organismo stesso.
I processi di apprendimento investono, perciò, le strutture biologiche dell’individuo, non il
suo genoma, destinato a rimanere tale. Sono come degli schemi di riconoscimento, presenti negli
individui, che si attivano e si sviluppano normalmente nelle relazioni interpersonali e sotto la
pressione di fattori culturali e ambientali.
L’influenza dei fattori epigenetici sulla struttura e sulla funzionalità del cervello è ormai
largamente riconosciuta, vedi ad esempio (Levi-Montalcini, 1998), o (Boncinelli, 2001 e 2005).
Quest’ultimo autore si esprime così: “…Il mondo umano circostante non si stampa in sostanza nel
suo genoma, ma nel suo corpo e nel suo cervello”7 . Sempre con le parole di Boncinelli, “Con la
specie umana l’evoluzione biologica ha superato se stessa e ha condotto a una sorta di paradosso.
Nel nostro caso, infatti, il patrimonio genetico signore quasi assoluto della vita e del
comportamento degli animali inferiori, ha per così dire volontariamente abdicato, lasciando ampi
spazi all’azione dell’ambiente circostante, all’apprendimento e all’educazione. Ci possiamo
considerare svincolati dalla nostra biologia, ma non dobbiamo dimenticare che la libertà di cui
godiamo è una conquista e un grazioso regalo dei nostri stessi geni, regalo che non è toccato, tanto
per dirne una, né ai calamari, né ai ranocchi.”: 8
6
“Si intende per Epigenetica una qualunque attività di regolazione dei geni tramite processi chimici che non
comportino cambia menti nel codice del DNA ma possono modificare il fenotipo dell’individuo e/o della progenie”
(dall’enciclopedia on-line Wikipedia, 20/12/2006). Per Changeux l’aggettivo epigenetico combina due significati: l’idea
di sovrapposizione all’azione dei geni, in seguito soprattutto all’apprendimento e all’esperienza, e l’idea di sviluppo
coordinato ed organizzato dai geni stessi (Changeux, 2002), pag. 188.
7
(Boncinelli, 2005), pag. 20.
8
(Boncinelli, 2001), pag…
3
L’architettura neurale, dinamica lungo tutto l’arco temporale della vita dell’individuo, sarebbe
dunque il risultato di una capacità endogena del cervello, presente già a livello embrionale, di
produrre un’enorme varietà di prerappresentazioni del mondo che si concretizza in forme transitorie
delle strutture sinaptiche. 9 L’attitudine a formare prerappresentazioni sarebbe quindi di origine
genetica, quasi una predisposizione innata a comprendere ed apprendere; il processo di selezione e
di stabilizzazione avviene per tentativi ed errori ed è guidato da un principio analogo al principio di
“rilevanza” cognitiva postulato da Sperber e Wilson nell’ambito dei loro studi sulla comunicazione
(Sperber & Wilson, 1986). La rilevanza è valutata mediante corrispondenza con la realtà fisica e
sociale: “Una pre-rappresentazione data può essere stabilizzata o meno in funzione del segnale
ricevuto dal mondo esterno. La risposta venuta dall’esterno è determinante. Essa costituisce un test
dell’accordo (voce ‘Verità’ nell’Encyclopédie), della conformità, o anche dell’adeguazione della
pre-rappresentazione nei confronti dell’ambiente. Questo test permette di scoprire se essa ‘dia
senso’ o meno e potrebbe fondarsi su due meccanismi plausibili. Il primo è la ‘selezione per
ricompensa’ e sarebbe questo meccanismo a essere usato di preferenza per la valutazione delle
azioni. […] Potrebbe darsi che un altro scenario sia più adeguato al caso della percezione
sensoriale: parleremo a questo proposito di ‘selezione per risonanza’ (Dehaene, Changeux &
Nadal, 1987). Tale scenario si fonda sulla corrispondenza tra l’attività percettiva suscitata da
stimolazioni sensoriali e la pre-rappresentazione esistente al momento dell’esperienza sensoriale”
(Changeux, 2002, pag. 65-66)
Grazie a questo processo, si formano reti stabili di neuroni che costituiscono l’aspetto
“epigenetico” del cervello nonché la base neurale per rappresentazioni di ordine superiore.
L’Autore porta a sostegno della sua teoria una grande quantità di dati: tratti da studi su pazienti
affetti da lesioni cerebrali, ottenuti con l’impiego di tecniche sperimentali di imaging cerebrale e
anche desunti dalla costruzione di appositi modelli informatici di reti neurali. È impossibile
riportare qui questa messe imponente di dati, che vanno oltre i limiti del nostro lavoro e spesso
anche della nostra capacità di decodifica. Ci limiteremo pertanto a riferire più oltre solo alcune
evidenze sperimentali, che ci sembrano più significative ai nostri fini (in fondo anche in questo
nostro contesto può applicarsi il Principio della rilevanza!), rinviando al testo di Changeux per
maggiori approfondimenti.
Le capacità del cervello di formare e comunicare rappresentazioni coscienti del mondo e di
verificarne la veridicità costituiscono allora l’evoluzione, in senso darwiniano, di una capacità
innata; pur essendo di natura culturale, esse si trovano inscritte in un “involucro genetico fissato”
nei nostri cromosomi. Un primo accordo del cervello con la realtà del mondo si sarebbe infatti
realizzato nel corso dell’evoluzione genetica per selezione naturale sino a homo sapiens. Sono in
altri termini sopravvissuti i gruppi di individui capaci, a partire dal loro patrimonio genetico, di
formarsi dell’Ambiente circostante rappresentazioni sufficientemente “vere” da garantire loro
l’omeostasi e la possibilità di riprodursi. Ciò vuol dire anche che nel corso dell’evoluzione
biologica si sono considerevolmente accresciute a livello epigenetico le capacità innate del cervello
di rappresentare il mondo, dall’ambiente fisico e biologico circostante sino al campo senza limiti
delle interazioni sociali e culturali. Viene da pensare a Galileo, quando dice: “Ma io penso prima la
natura aver fatto le cose a modo suo e poi i pensieri degli uomini atti a capirla”10 . E forse può
essere riletta in questa chiave anche la lunga diatriba concernente il carattere più o meno inna to dei
concetti della matematica, dalla tradizionale visione platonica fino alle posizioni filosofiche più
relativistiche (cfr. al riguardo l’interessante dibattito in (Changeux & Connes, 1989)).
2.2.2. L’attività spontanea del cervello
9
L’attività elettrica spontanea si manifesta presto nello sviluppo embrionale del sistema nervoso, tanto da poter essere
registrata nel feto prima del secondo mese (Bears et al., 2000) ed è rilevabile nell’adulto sotto forma di onde
elettroencefalografiche.
10
(Galilei, 1964), vol. II, pag. …
4
I neuroni si comportano come generatori spontanei di impulsi, dando luogo ad un’attività
spontanea del cervello; quest’ultimo, come già anticipato da molti studiosi – per esempio (Neisser,
1976) e (Berthoz, 1997), – lungi dall’essere ricettore passivo di informazioni prove nienti dal mondo
esterno, si comporta come un sistema autonomo che “proietta” in permanenza informazione verso
questo. Già a livello di singole cellule nervose la scarica spontanea costituisce parte integrante del
funzionamento e della struttura del cervello durante tutti gli stadi dello sviluppo, anzi la stessa
architettura neurale sarebbe conseguenza di tale attitudine di natura genetica: in un processo
analogo a quello che un bimbo mette in atto per imparare a camminare, il cervello produrrebbe una
grande quantità di connessioni sinaptiche temporanee, di cui solo alcune sopravvivono, selezionate
da un processo di natura epigenetica e stabilizzate dalla stessa attività spontanea. Gli agenti di
natura genetica ed epigenetica risultano fortemente integrati nell’evoluzione del cervello non solo
nelle prime fasi dello sviluppo in cui si devono stabilizzare le connessioni nervose atte a garantire la
sopravvivenza “sensomotoria” del bimbo piccolo, ma anche a livello culturale più avanzato, quando
il problema della sopravvivenza diventa marcatamente culturale.
L’attività intrinseca spontanea del cervello si manifesta attraverso impulsi elettrici prodotti, in
totale assenza di stimoli, dalle cellule nervose; impulsi del tutto analoghi a quelli prodotti
dall’interazione con l’ambiente. Si può rendere conto integralmente di questa attività intrinseca a
livello di meccanismi fisico-chimici: gli impulsi sono prodotti dai cosiddetti oscillatori molecolari
(Berridge & Rapp, 1979), combinazioni di proteine che generano fluttuazioni lente del potenziale
elettrico della membrana del neurone che, al di là di una certa soglia, scatena scariche spontanee.
Il ruolo che l’attività spontanea gioca nella strutturazione dell’architettura neurale è sostenuto
da alcuni esperimenti su animali (Changeux, 2002), pag. 200. Ad esempio inducendo nei topi un
danno genetico che blocca selettivamente la secrezione dei neurotrasmettitori, si osserva (Verhage
et al., 2000) che il cervello di questi topolini mutanti, finché restano nell’utero della madre, si
sviluppa normalmente, dunque anche in assenza dell’attività sinaptica, ma alla nascita i topolini
muoiono a causa di estesi fenomeni degenerativi. L’attività spontanea dell’embrione non è dunque
indispensabile alla morfogenesi globale del cervello, ma è necessaria perché si stabilizzino e si
conservino, in modo genetico, le connessioni nervose fino all’età adulta.
Il ruolo dell’attività spontanea sarebbe altresì centrale nella formazione delle conoscenze,
funzionando come un generatore di diversità di tipo darwiniano di prerappresentazioni11 : stati di
attività dinamici, spontanei e transitori di popolazioni di neuroni. L’acquisizione di conoscenza
sarebbe allora il risultato di una selezione di prerappresentazioni e si concretizzerebbe sotto forma
di una rete di neuroni distribuiti in più aree distinte del cervello. “La distribuzione di queste aree
non sarebbe aleatoria ma rifletterebbe l’organizzazione dei sistemi sensoriali e motori.”
(Changeux, 2002, pag. 54). Questa intuizione, appena accennata da Changeux, verrà ampiamente
ripresa e argomentata in seguito da (Rizzolatti & Sinigaglia, 2006) e (Gallese & Lakoff, 2005),
centrando soprattutto l’attenzione su specifici aspetti di carattere sensomotorio (tipicamente l’azione
di “grasping”).
A livello funzionale si può interpretare l’attività spontanea di insiemi specializzati di neuroni
con un’attitudine intrinseca dell’organismo ad esplorare e modellizzare l’ambiente fisico, in un
processo “spontaneo” di rimodellizzazione progressiva e di esplorazio ne organizzata guidato
dall’esperienza delle risposte provenienti dall’ambiente. L’attività elettrica spontanea è allora il
corrispettivo fisiologico delle caratteristiche di autoattivazione ed autoorganizzazione del cervello
come sistema (Laplane & Dubois, 2001).
In (Gallese & Lakoff, 2005) si propone un’analisi più fine delle basi neurologiche del
processo di concettualizzazione che prolunga la funzione delle prerappresentazioni: “Understanding
is imagination”.
2.2.3. La plasticità neuronale
11
Dette anche schemi preliminari o neuronali (Arbib et al., 1998). Cfr. (Changeux, 2002), pag. 63, per ulteriori
riferimenti.
5
L’aver riconosciuto la natura evolutiva del cervello a livello epigenetico prelude al
riconoscimento dell’effetto dell’apprendimento sull’architettura neurale. Con il termine plasticità
neuronale Changeux indica la capacità dei neuroni e delle loro sinapsi di cambiare proprietà in
funzione del loro stato di attività e ne riporta rilevazioni effettuate a livello elettrico e molecolare
(Changeux, 2002, pag. 31).
Lo sviluppo stesso del cervello a livello embrionale è frutto della plasticità neuronale; infatti si
è potuto osservare che numerose cellule nervose prodotte dalle divisioni cellulari embrionali
muoiono prima di diventare neuroni adulti e questa morte cellulare è ritardata o accelerata
dall’attività nervosa spontanea; ad analogo processo sono soggette le sinapsi, che crescono e si
dividono durante lo sviluppo, ma possono essere anche eliminate e quindi, successivamente,
rigenerarsi per gemmazione. Il processo non è limitato al periodo dello sviluppo embrionale; infatti,
dopo la nascita, al “motore” di natura genetica (l’attività nervosa spontanea) si affianca un “motore”
di natura epigenetica: l’interazione con l’ambiente che dà luogo ad una regolazione epigenetica
dello sviluppo delle reti neurali ed alla costruzione delle conoscenze individuali. Nell’uomo quasi il
50% delle sinapsi si forma dopo la nascita e il loro numero continua a crescere o diminuire per tutta
la vita12 .
Il cervello sarebbe quindi geneticamente predisposto ad “apprendere”: l’attitudine a creare
prerappresentazioni del mondo e di sé sarebbe genetica e costituirebbe la chiave, non solo del
funzionamento, ma anche dell’organizzazione neurale del cervello; la stessa attitudine evolverebbe
a livello epigenetico garantendo una marcata plasticità neurale che si conserva per tutta la vita,
strettamente vincolata all’apprendimento. È la plasticità che dà conto della flessibilità funzionale
delle reti di neuroni, della loro autoorganizzazione nonché della capacità di accumulo delle
conoscenze; essa è inoltre, in un certo senso, il frutto dell’attività elettrica spontanea, in quanto i
meccanismi di trasmissione delle informazioni sono regolati dall’attività spontanea o evocata di cui
sono mediatori.
Nella letteratura sono riportati innumerevoli e disparati dati sperimentali a sostegno della tesi
della pla sticità. Vedremo più avanti un esempio tratto da (Rizzolatti & Sinigaglia, 2006, pag. 133),
che si riferisce agli effetti che uno stesso evento (in questo caso musicale), produce in soggetti
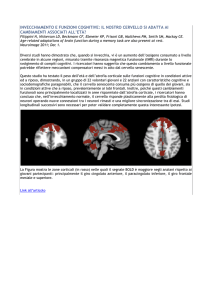
diversi a livello di zone del cervello coinvolte; un’analoga osservazione è descritta nella Fig. 1,
tratta da …., dove si evidenzia come l’ascolto di musica attivi risposte più o meno intense in termini
di estensione di aree cerebrali coinvolte, a seconda che l’ascoltatore sia più o meno competente nel
campo specifico.
Immagine (Fig. 1)
Riportiamo qui invece da (Changeux, 2002, pag. 210), una significativa evidenza sperimentale
riguardante lo studio di individui affetti da cecità precoce, esaminati dopo l’apprendimento del
sistema di lettura Braille. Tale sistema richiede una sensibilità marcata dei polpastrelli per
distinguere la distribuzione dei punti in rilievo; le tecniche di imaging funzionale rivelano, dopo un
anno di addestramento intensivo, un’estensione significativa della corteccia parietale
somatosensoriale dell’emisfero cerebrale sinistro, un’area coinvolta in particolare nella percezione
tattile dello spazio. Ma l’imaging funzionale rivela anche un fenomeno inatteso: una forte
attivazione delle aree visive primarie e secondarie della corteccia occipitale, zone specializzate nella
visione nei soggetti vedenti: in seguito all’acquisizione del Braille, le aree visive dei soggetti ciechi
diventano capaci di ricevere ed elaborare informazioni tattili. Dunque l’apprendimento del Braille
produce un cambiamento significativo nella connettività neuronale e nella sua funzionalità: l’ipotesi
più plausibile è che già alla nascita esistano connessioni funzionali tra la corteccia somatosensoriale
12
Cfr. (Changeux, 2002), pag. 194, dove sono riportati altri dati, riguardanti ad esempio il confronto con varie specie
animali.
6
e quella visiva e che l’apprendimento del sistema Braille abbia l’effetto di selezionare ed
amplificare queste vie preesistenti a vantaggio della lettura tattile.
Alcuni studi condotti su animali diversi e reperti archeologici di cervelli di ominidi fanno
ritenere che la plasticità cerebrale sia attiva anche su scala biologica: nel corso dell’evoluzione
biologica si sono considerevolmente accresciute le capacità innate del cervello di rappresentare il
mondo 13 . Gli studi nell’ambito dell’epigenetica sono peraltro in pieno sviluppo e non mancano
risultati che inducono a rivedere consolidati paradigmi. Ha destato scalpore ad esempio la
recentissima osservazione della capacità di una pianta (Arabidopsis Thaliana) di trasmettere ai
propri discendenti informazioni ricavate dall’esperienza maturata in relazione al suo ambiente
(Molinier et al., 2006).
In questo quadro le capacità numeriche innate (messe in luce ad esempio in (Dehaene, 1997))
sarebbero il risultato di una selezione epigenetica di reti neurali sopravvissute nel cervello di homo
sapiens (ma anche di tanti animali) perché funzionali alla sopravvivenza. 14
2.2.4. La strutturazione del cervello
Per stabilire un legame causale tra anatomia e funzioni del cervello occorre un modello globale
di organizzazione delle reti neurali; intanto le prove sperimentali della variabilità epigenetica della
connettività cerebrale, della plasticità – modulata dall’uso – di sinapsi, vie di connessioni e mappe
corticali, delle capacità neuronali d’attivazione, spontanea o indotta, di preparazione, di
anticipazione, di proiezione d’ipotesi, di decisione, di stimolazione interna delle azioni, degli
avvertimenti o dei processi interni, inducono ad abbandonare modelli semplicistici del cervello come
un organo definitivamente costituito e incapace di rigenerazione, essenzialmente recettivo e reattivo,
rigidamente compartimentato e gerarchizzato a partire dai recettori periferici sino ai centri
associativi e, da lì, verso la fuoriuscita motrice.
L’analogia con altri sistemi meglio studiati, e i dati sperimentali di cui si dispone, conducono
Changeux a identificare due livelli principali di organizzazione globale della rete neurale
sovrapposti e interrelati: strutture che si sviluppano “verticalmente” attraverso gli strati della
corteccia e sono organizzate gerarchicamente (Jackson, 1932) e strutture “orizzontali” di territori
specializzati a livello funzionale, collegati tra loro per mezzo di mappe corticali (Changeux &
Dehaene, 1989).
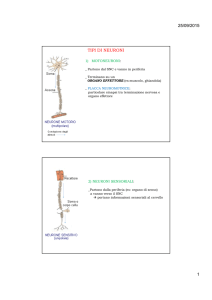
Figura 1. Le aree del cervello
Numerosi studi sull’anatomia della corteccia cerebrale dei vertebrati (ma in (Rizzolatti &
Sinigaglia, 2006) sono aggiunte conferme da esperimenti sull’uomo) suggeriscono l’esistenza di
una distribuzione di aree corticali proprie di ciascuna specie e investite di funzioni specializzate; si
dispone inoltre di evidenze sperimentali del fatto che al crescere del grado di evoluzione dei
vertebrati aumenta sia il numero di livelli gerarchici della corteccia che il numero delle mappe in
parallelo (Changeux, 2002, pag. 35). In altri termini, con l’evoluzione sia biologica che individuale,
l’organizzazione del sistema nervoso diviene sempre più complessa, sui registri gerarchico e in
parallelo: le sue capacità di esplorazione e di rappresentazione evolvono anch’esse e si estendono
dall’ambiente fisico e biologico all’ambiente sociale e culturale. Parallelamente, la plasticità della
rete e le sue capacità di apprendimento aumentano, le disposizioni innate si arricchiscono in termini
di complessità e di flessibilità.
Le diverse aree cerebrali non hanno tuttavia un’autonomia funzionale rigida, come pensava
Gall, che per primo ne ha ipotizzato l’esistenza (Gall, 1822-1825). Ciò è oggi noto anche a livello
13
14
Cfr. (Levi-Montalcini, 1998) e (Changeux, 2002), pag. 180.
Un’ipotesi analoga è avanzata in (Devlin, 2000).
7
anatomico: sono state evidenziate fin dagli anni Trenta connessioni reciproche tra cellule nervose
distanti e, più recentemente, connessioni di questo tipo sia verticalmente tra livelli sovrapposti che
orizzontalmente tra aree corticali distinte anche di emisferi diversi (Dehaene, Kerszberg &
Changeux, 1998). Di conseguenza non si può capire il significato “funzionale” di queste aree
cerebrali senza analizzare i rapporti che ciascuna di esse mantiene con gli altri elementi del sistema
e, più in generale, con il contesto fisico e socioculturale in cui si sviluppa.
Per avere un’idea del livello di complessità dell’integrazione tra le due strutture principali di
organizzazione, si prenda ad esempio il sistema visivo della scimmia, per il quale si dispone di un
numero maggiore di dati (Bears et al., 2000): la funzionalità si appoggia su 14 livelli sovrapposti,
dalla retina alla corteccia prefrontale. Di essi almeno 6 sono situati fra le 32 aree visive della
corteccia cerebrale e contribuiscono contemporaneamente all’elaborazione della forma, del colore e
del movimento degli oggetti a tre dimensioni. Estremamente significativo per l’analogia con il
funzionamento del cervello è il fatto che il sistema può funzionare dal basso verso l’alto, per
esempio al momento della percezione di un oggetto visivo, come pure dall’alto verso il basso,
quando si richiama alla memoria l’immagine dell’oggetto.
Dati sperimentali ancora più recenti (Rizzolatti & Sinigaglia, 2006) sostengono e raffinano il
modello di architettura neuronale proposto da Changeux; di questi alcuni risultano particolarmente
significativi per i nostri scopi e saranno riportati come approfondimenti.
2.2.5. Compiti elementari e compiti “con sforzo”
Una distinzione importante che si ritrova in molti autori è quella tra compiti elementari e
compiti “con sforzo”. I primi sono sostanzialmente quelli a cui gli esseri umani (e non solo) sono
sottoposti quotidianamente per soddisfare esigenze primarie: procurarsi acqua quando si ha sete,
fuggire davanti a un pericolo, riconoscere un viso o un oggetto, eccetera. Essi sono eseguiti in
condizioni normali attivando circuiti “innati” o comunque precablati nel nostro cervello e appunto
non richiedono sforzo. Appartengono a questa categoria anche compiti aritmetici elementari come
valutare la numerosità di piccoli insiemi (Dehaene, 1997), (Devlin, 2000).
Costituiscono invece compiti con sforzo tutti quelli che richiedono un ricorso ad altre aree e
ad altre funzioni del cervello (la memoria, l’attività di immaginazione, la produzione di numerose
prerappresentazioni e la loro selezione, ecc.). Sotto altro nome Vygotskij distingue tra funzioni
elementari e funzioni superiori del cervello. È anche interessante il punto di vista di (Gallese &
Lakoff, 2005), secondo i quali anche per compiti di questa natura sono attivi gli stessi circuiti
cerebrali che sono preposti alle attività sensoriali e motorie (“parassitate” per scopi diversi).
Appartengono a questa categoria gran parte dei compiti richiesti nell’apprendimento della
matematica, e risulta perciò importante ai nostri fini esaminare più da vicino cosa accade
nell’affrontare questo tipo di compiti.
Alcuni esperimenti condotti da Oliver Houdé, ricercatore del Groupe d’Imagerie
Neurofonctionelle (GIP) del CNRS, su soggetti impegnati in compiti cognitivi complessi risultano
particolarmente interessanti non solo per mettere in discussione modelli di apprendimento di tipo
stratificato, in cui conoscenze di ordine superiore “scardinano” quelle più elementari, ma anche
perché forniscono uno strumento interpretativo forte di molti “comportamenti fallimentari” dei
nostri studenti. L’esperimento (Houdé et al., 2000), prevede l’esposizione di un gruppo di volontari
al test di Wason, un buon esempio di “compito con sforzo”. Com’è noto, tale test viene usato
spesso, ad esempio in (Leron, 2004), a sostegno dell’ipotesi che modalità di ragionamento di tipo
inferenziale non sono alla portata di tutti, in virtù della forte percentuale di fallimenti che una
sterminata letteratura sul test ha evidenziato. Riportiamo qui brevemente alcuni elementi salienti
dell’esperimento di Houdé, rinviando a (Iannece et al., 2006) per maggiori dettagli.
Nella versione classica del test di Wason (Wason, 1983) si usano delle speciali carte che
hanno su un lato un numero (naturale) e sull’altro lato una lettera. Disposte su un tavolo quattro
8
carte come in figura, viene posta la domanda: Quali delle quattro carte dobbiamo sollevare come
minimo, per controllare se è soddisfatta la regola “Dietro ogni vocale c’è un numero dispari”?
A
1
B
2
Si tratta dunque di un quesito che coinvolge l’uso dell’implicazione logica e la cui soluzione
non è certo immediata: come si è detto, indipendentemente dal livello scolare e di età, le risposte
sbagliate costituiscono la grande maggioranza.
L’esperimento di Houdé si realizza con tecniche di brain imaging e si svolge in due fasi.
Sottoposti all’inizio alcuni volontari ad una variante del test di Wason, si riscontra che le zone del
cervello che vengono attivate durante l’esecuzione del compito, in caso di errore, sono quelle,
localizzate nella parte posteriore, normalmente coinvolte nelle funzioni percettive. Nella seconda
fase il campione viene diviso in due gruppi, assoggettati a due diversi tipi di sollecitazione,
rispettivamente sul piano razionale e su quello emotivo: al primo gruppo si forniscono informazioni
più dettagliate sugli aspetti logici del test, mentre i soggetti del secondo gruppo vengono messi in
guardia rispetto alla possibilità di cadere in errore. Nuovamente sottoposti al test, si verifica che la
percentuale di errori dei soggetti del primo gruppo resta sostanzialmente immutata, mentre nel
secondo gruppo il tasso di successo diventa pari al 90%. L’osservazione in iconografia cerebrale
indica che le zone del cervello attivate dagli individui del primo gruppo risultano essere le stesse
utilizzate per risolvere il problema durante la prima fase. Gli individui sollecitati ne lla sfera emotiva
attivano invece delle zone differenti, situate nella zona anteriore del cervello: precisamente la
corteccia frontale ventro- mediana (CPVM, un'area del cervello che collega emozioni e
ragionamento).
L’esperimento è stato progettato ed interpretato da Houdé a sostegno della teoria esposta in
(Damasio, 1994) sull’interazione dei fattori emotivi con i processi del pensiero “razionale”; esso
mostra infatti che l’attivazione di emozioni (in questo caso negative) può risultare decisiva anche
nei ragionamenti logici, perché rende i soggetti capaci di inibire la strategia percettiva utilizzata
precedentemente e li induce a scegliere una strategia di tipo logico più idonea al compito. Questa
stretta connessione tra emozioni e razionalità in ogni comportamento cognitivo degli esseri umani è
un aspetto in sé molto importante, ma in questa fase noi la trascureremo: per i nostri scopi risulta
maggiormente significativa la dimostrazione dell’esistenza, nel cervello, di due zone funzionali
diverse deputate all’elaborazione di due tipi diversi di “ragionamento”. I circuiti cerebrali
specializzati nel trattare le informazioni provenienti dal sistema sensomotorio sono naturalmente i
primi ad attivarsi, in quanto consentono decisioni rapide, essenziali per garantire la sopravvivenza
(Damasio, 1994). Ma questi circuiti, come già detto, non sono i soli, ed il modo di funzionare
percettivo- motorio non è l’unico modo naturale di funzionare verso cui il cervello è geneticamente
predisposto. Altri circuiti sono dedicati al ragionamento logico e si dà il caso che siano gli stessi che
regolano e governano le emozioni.
2.3. ACQUISIZIONI RECENTI
La velocità con cui si susseguono le ricerche nel campo della neurofisiologia, ci induce a
integrare la teoria di Changeux con dati più recenti. Accedere direttamente ai lavori di ricerca del
settore non è tuttavia agevole, sia per le questioni analizzate, spesso troppo specifiche, sia per il
linguaggio specialistico. Risulta perciò prezioso il libro di Giacomo Rizzolatti e Corrado Sinigaglia
So quel che fai. Il cervello che agisce e i neuroni specchio, uscito molto di recente, (Rizzolatti &
Sinigaglia, 2006) che seleziona e riformula in linguaggio accessibile alcuni sviluppi sperimentali e
9
teorici di neuroscienze. La tesi di fondo degli Autori è la centralità della corteccia motoria per il
funzionamento cognitivo in senso lato: “….al di là dell’organizzazione dei nostri comportamenti
motori, anche certi processi di solito considerati di ordine superiore e attribuiti a sistemi di tipo
cognitivo, quali per esempio la percezione e il riconoscimento degli atti altrui, l’imitazione e le
stesse forme di comunicazione gestuali o vocali, possono rimandare al sistema motorio e trovare in
esso il proprio substrato neurale primario”15 .
In ogni caso alcuni aspetti nodali della teoria di Changeux vengono meglio definiti,
approfonditi ed arricchiti sia a livello teorico che sperimentale. Ci riferiamo in particolare alla
complessità dell’architettura neurale; ai meccanismi neurali che sottendono il forte livello di
integrazione tra processi percettivi, cognitivi e motori; alla natura “pragmatica” del funzionamento
cerebrale, alla centralità dell’azione in ogni funzionamento cognitivo.
2.3.1. Complessità e integrazione
Rimandiamo a (Rizzolatti & Sinigaglia, 2006) per i dettagli sull’architettura neurale,
limitandoci a riportare qui alcune conclusioni che gli autori traggono dai dati sperimentali in loro
possesso: “I dati sperimentali di questi ultimi vent’anni hanno profondamente cambiato la
concezione del sistema motorio che a lungo ha dominato la scena nella fisiologia e nelle
neuroscienze. La corteccia frontale agranulare e la corteccia parietale posteriore risultano
costituite da un mosaico di aree anatomicamente e funzionalmente distinte, che sono fortemente
connesse fra di loro e formano circuiti destinati a lavorare in parallelo e a integrare le
informazioni sensoriali e quelle motorie relative a determinati effettori. E lo stesso vale per i
circuiti che coinvolgono le aree della corteccia prefrontale e di quella del cingolo, e che sono
responsabili della formazione delle intenzioni, della pianificazione a lungo termine e della scelta
del momento in cui effettivamente agire”16 .
Buona parte del libro è dedicata a fornire evidenze sperimentali del fatto che l’elaborazione
dell’informazione sensoriale e di quella motoria sono riconducibili ad un formato comune,
codificato da specifici circuiti parieto- frontali. In quest’ottica non esiste una percezione “a priori”,
ad esempio visiva, che determina i nostri movimenti nello spazio, è la nostra “intenzione e capacità”
di azione che determina la percezione degli oggetti e dello stesso spazio intorno a noi17 . “La
presenza nei circuiti neurali di risposte visive connesse ad attivazioni motorie sembra suggerire che
(nella codifica) dello spazio (fisico) […] la scarica dei neuroni (coinvolti) non segnali
semplicemente la posizione dello stimolo entro uno spazio puramente visivo, sulla base di un
qualche sistema di coordinate geometriche, […] bensì rifletta l’evocazione di un atto motorio
potenziale diretto verso quello stimolo e in grado, indipendentemente o meno dalla sua attuazione,
di localizzarlo nei termini di una possibilità di azione”18 .
Una base fisiologica dell’integrazione costitutiva tra strutture che elaborano l’informazione
motoria e sensoriale è costituita dall’esistenza dei neuroni canonici e dei neuroni specchio di cui si
dirà più avanti.
2.3.2. La natura pragmatica del funzionamento cerebrale
Grazie alla modalità scimmia libera (cfr. § 2.1.1), si è scoperta una proprietà estremamente
importante sul piano teorico dei neuroni dell’area F5 della corteccia premotoria 19 : “la maggior parte
15
(Rizzolatti & Sinigaglia, 2006), pag. 22.
(Rizzolatti & Sinigaglia, 2006), pag. 20-21.
17
Questo punto di vista riprende in effetti quanto già asserito da altri autori come (Neisser, 1976), (Bozzi, 1990),
(Berthoz, 1997), e intuito ancora prima ad esempio da Poincaré e Mach. Cfr. anche (Gallese & Metzinger, 2003).
18
(Rizzolatti & Sinigaglia, 2006), pag. 65.
19
Area molto studiata, ad esempio in (Gallese & Lakoff, 2005), perché contiene rappresentazioni motorie della mano e
della bocca e molte delle evidenze sperimentali interessano scimmie che afferrano cibo e lo portano alla bocca.
16
10
dei neuroni di F5 non codifica singoli movimenti bensì atti motori, cioè movimenti coordinati da un
fine specifico”20 . Nell’area F5 ci sarebbe qualcosa che può essere paragonato ad un “vocabolario di
atti motori”, le cui parole sarebbero rappresentate da popolazioni di neuroni che si attivano nel
momento in cui la scimmia compie delle azioni dirette ad uno scopo come tenere, afferrare,
manipolare 21 . I neuroni non si attivano quando la scimmia compie i medesimi gesti senza l’obiettivo
specifico, ad esempio il circuito che si attiva quando la scimmia afferra il cibo con la bocca non si
attiva quando l’apertura e la chiusura della bocca è determinata da stimoli di tipo emotivo. È
sorprendente questa specificità dei neuroni, ma altrettanto sorprendente è l’idea che l’attivazione di
un neurone non sia correlata al movimento di un braccio tout court, ma lo sia solo se il movimento è
effettuato per raggiungere uno scopo. (vedi anche (Gallese & Lakoff, 2005), pag. 7).
Una parte dei neuroni di F5 che si attivano durante l’esecuzione di atti motori rispondono
anche a stimoli visivi (Rizzolatti et al., 1988): in altri termini, i messaggi inviati da tali neuroni,
detti anche visuomotori, agli altri centri sono esattamente gli stessi sia quando la scimmia
interagisce con un oggetto o con il cibo, sia quando si limita a osservarlo. Nel caso dell’effettiva
esecuzione dell’atto, la scarica del neurone rappresenta l’attivazione di un comando motorio, nel
caso della semplice osservazione la sua attivazione riflette l’evocazione dello stesso pattern motorio
che rimane però allo stato di atto potenziale (Rizzolatti e Sinigaglia, 2006, pag. 46). È come dire
che la vista di un oggetto attiva nel cervello una prerappresentazione dell’azione necessaria a
raggiungerlo. Ma c’è di più: alcuni recenti risultati sperimentali 22 hanno confermato un assunto un
po’ ardito di (Gibson, 1979) (cfr. anche (Bozzi, 1990) e (Luccio, 1993)), e cioè che la percezione
visiva di un oggetto comporta l’immediata e automatica selezione delle proprietà dell’oggetto stesso
che ci consentono di interagire con esso (affordances); si mostra ad esempio in uno degli
esperimenti che l’attivazione neurale generata dalla prensione o dalla vista di oggetti di forma
diversa è la stessa se la stessa è la modalità di prensione: “il cervello che agisce è innanzitutto un
cervello che comprende”, sintetizzano gli Autori (Rizzolatti e Sinigaglia, 2006, pag. 3).
2.3.3. I neuroni specchio
La scoperta del funzionamento di questi neuroni nelle scimmie avvenne fortuitamente. Fu per
caso, infatti, che in laboratorio si osservò come nel momento in cui un ricercatore aveva teso la
mano per afferrare delle arachidi, l’EEG di uno dei macachi osservati registrasse un aumento di
potenziale in corrispondenza dell’area F5, lo stesso aumento di potenziale registrato quando erano
gli stessi macachi ad afferrare le arachidi. Fu così che i ricercatori scoprirono un gruppo di neuroni
fino allora sconosciuti. Sono cellule che diventano attive non solo quando le scimmie compiono
movimenti della mano guidati da un’esplorazione del campo visivo, ma anche ogniqualvolta
percepiscono movimenti analoghi nelle persone presenti nell’ambiente circostante, e persino
quando odono il suono associato ad un’azione, come rompere le noccioline. Infatti sono stati
identificati non solo mirror bimodali di tipo visuo- motorio, ma anche mirror trimodali, audiovisuo-motori23 .
Un’azione compiuta da un altro fa “risuonare”, nell’interno di chi osserva l’azione o ode il
rumore, i neuroni che si attiverebbero se fosse lui stesso ad agire. Queste cellule furono chiamate
dagli scopritori “neuroni specchio” perché rappresentano in modo speculare l’azione motoria di un
animale nel cervello di un altro.
20
(Rizzolatti & Gentilucci, 1988), (Rizzolatti et al., 1988).
Accanto ai neuroni che codificano lo scopo generale dell’atto (tenere, afferrare, rompere, ecc.) se ne sono rivelati
altri che codificano la maniera in cui un atto motorio specifico può essere eseguito (presa di precisione, presa con le
dita, ecc.); altri, infine, devoluti al controllo della segmentazione temporale dell’atto nei movimenti elementari che lo
compongono (apertura della mano, chiusura della mano) (Rizzolatti & Sinigaglia, 2006), pag. 45.
22
(Murata et al., 1997), (Rizzolatti et al., 2000), (Gallese, 2000). Cfr. (Rizzolatti e Sinigaglia, 2006), pag. 35-38.
23
(Kohler et al., 2002).
21
11
Più recentemente, la messa a punto di tecniche di osservazione cerebrale meno cruente ha
consentito di rilevare e analizzare dal punto di vista funzionale il sistema dei neuroni specchio
nell’uomo 24 . Com’è naturale, tale sistema risulta più esteso e funzionalmente più raffinato di quello
delle scimmie; in particolare “esso codifica atti motori transitivi e intransitivi; è in grado di
selezionare sia il tipo d’atto sia la sequenza dei movimenti che lo compongono; infine, non
necessita di un’effettiva interazione con gli oggetti, attivandosi anche quando l’azione è
semplicemente mimata” (Rizzolatti & Sinigaglia, 2006, pag. 121).
Concludendo, il vocabolario d’atti, grazie all’azione dei neuroni specchio costituisce il nostro
budget cognitivo per l’interpretazione e la comprensione delle azioni degli altri. Si tratta qui di una
“comprensione pragmatica, preconcettuale e prelinguistica, e tuttavia non meno importante, poiché
su di essa poggiano molte delle nostre tanto celebrate capacità cognitive”25 . Nella scimmia, come
nell’uomo, “tale comprensione non investe solo singoli atti, bensì intere catene d’atti, e le diverse
attivazioni del sistema dei neuroni specchio mostrano come esso sia in grado di codificare il
significato che ogni atto osservato viene ad assumere a seconda delle azioni in cui potrà trovarsi
immerso”26 .
Ci sembra importante rimarcare che si sta parlando di una comprensione che non è legata
all’evocazione di esperienze pregresse a livello personale, è piuttosto un processo che si attiva
automaticamente, senza l’intervento cosciente dell’individuo, potremmo dire “senza sforzo” per
usare il linguaggio introdotto sopra per interpretare l’esperimento di Houdé.
Non appena vediamo qualcuno compiere un atto o una catena d’atti, i suoi movimenti, che lo
voglia o meno, acquistano per noi un significato immediato; naturalmente, vale anche l’inverso:
ogni nostra azione assume un significato immediato per chi la osserva. Il possesso del sistema dei
neuroni specchio e la selettività delle loro risposte determinano così uno spazio di azione
condiviso, all’interno del quale ogni atto e ogni catena d’atti, nostri o altrui, appaiono
immediatamente iscritti e compresi, senza che ciò richieda alcuna esplicita o deliberata
‘operazione conoscitiva’” (Rizzolatti & Sinigaglia, 2006, pag. 127).
Un interessante esperimento (Buccino et al., 2004), riportato dagli autori a sostegno della loro
tesi, consiste nel mostrare a dei volontari dei video privi di sonoro in cui un uomo, una scimmia e
un cane eseguono atti comunicativi (parlare, schioccare le labbra, abbaiare). (vedi Figure). Le aree
del cervello attivate durante la visione sono, come è prevedibile, diverse: la vista dell’uomo che
muove le labbra per parlare induce una forte attivazione nella parte posteriore del giro frontale
inferiore; questa diventa più debole durante l’osservazione della scimmia, ed è assente alla vista del
cane che abbaia. Ciò non vuol dire naturalmente, commentano gli autori, che non siamo in grado di
comprendere i movimenti di un cane che abbaia, ma che la loro comprensione appare legata
principalmente all’attivazione delle aree del cervello deputate alla visione. Queste si attivano anche
alla vista degli altri atti comunicativi, ma a differenza dell’abbaiare che non ci appartiene, negli altri
casi l’informazione proveniente dalle aree visive attiva gli atti motori potenziali codificati dal
sistema dei neuroni specchio, consentendo una comprensione immediata “in prima persona” delle
azioni osservate.
Due figure da Rizzolatti, pag.129 e pag. 132
2.3.4. I neuroni specchio e la plasticità neuronale
24
In verità una sorta di risonanza dell’osservatore con le azioni di un altro individuo era già stata rilevata (Fadiga et al.,
1995) con la registrazione dell’aumento di potenziali elettrici nei muscoli dell’osservatore, omologhi a quelli usati
dall’individuo sotto osservazione (Rizzolatti & Sinigaglia, 2006, pag. 115).
25
(Rizzolatti & Sinigaglia, 2006), pag. 3
26
Ivi, pag. 122.
12
Il sistema dei neuroni specchio è modificato dall’apprendimento. “La vista di atti eseguiti da
altri comporta una diversa attività cerebrale a seconda delle competenze motorie specifiche dei
soggetti in questione” (Rizzolatti & Sinigaglia, 2006), pag. 133. I due autori riportano in proposito
un esperimento (Calvo-Merino et al., 2005), in cui un video in cui erano rappresentati alcuni passi
di capoeira veniva sottoposto all’osservazione di un campione di volontari comprendente danzatori
classici, maestri di capoeira e persone che non avevano mai preso lezioni di ballo. La proiezione del
video determinava nei maestri di questa arte un’attivazione del sistema dei neuroni specchio
maggiore di quella registrata negli altri individui, ballerini classici o principianti che fossero.
Questo esperimento, ed altri simili riportati dagli autori, confermano il ruolo decisivo della
conoscenza motoria per la comprensione del significato delle azioni altrui, comprensione tanto più
immediata e profonda quanto più esteso è il patrimonio motorio dell’osservatore, non solo quello
proprio della specie, ma anche quello di ogni singolo individuo. Possiamo concludere dicendo che
l’ambito delle cose che si comprendono “senza sforzo” si estende man mano che si progredisce
nell’apprendimento.
2.3.5. L’influenza degli strumenti (da scrivere)
2.3.6. La concettualizzazione (da scrivere)
Bibliografia 27
Arbib, M.; Erdi, P. e Szentágothai, J. (1998). Neural Organization: Structure, Function and Dynamics, MIT Press,
Cambridge, Mass.
Bears, M.; Connors, B. e Paradiso, M. (2000). Neuroscience-Exploring the Brain, William and Wilkins, Baltimore.
Berridge, M. e Rapp, P.E. (1979). ‘A Comparative Survey of the Function, Mechanisms and Control of Cellular
Oscillations’, J. Exp. Biol., 81, 217-280.
Berthoz, A. (1997). Le Sens du mouvement, Éditions Odile Jacob, Paris.
Boncinelli, E. (2001). A caccia di geni, Di Renzo, Roma.
Boncinelli, E. (2005). ‘Necessità e contingenza della natura umana’, Micromega. Almanacco di
filosofia, 4.
Boncinelli, E. e Bottazzini, U. (2000). La serva padrona. Fascino e potere della matematica,
Raffaello Cortina, Milano.
Bozzi, P. (1990). Fisica Ingenua, Garzanti, Milano.
Bruner, J. (1966). Toward a Theory of Instruction, Harvard University Press, Cambridge, MS –
Trad. ital.: Verso una teoria dell’istruzione, Armando, Roma, 1967.
Buccino, G.; Lui, F.; Canessa, N.; Patteri, I.; Lagravinese, G.; Benuzzi, F.; Porro, C.A. e Rizzolatti, G. (2004). ‘Neural
circuits involved in the recognition of actions performed by non con-specifics: an fMRI study’, Journal of
Cognitive Neuroscience, 16, 114-126.
Butterworth, B. (1999). The Mathematical Brain. MacMillan, London – Trad. ital.: Intelligenza
matematica, Rizzoli, Milano, 1999.
Calvo-Merino, B.; Glaser, D.E.; Grèzes, J. E.; Passingham, R.E. e Haggard, P. (2005). ‘Action observation and acquired
motor skills: an fMRI study with export dancers’, Cerebral Cortex, 15, 8, 1243-1249.
Changeux, J.-P. (1983). L’Homme Neuronal. Fayard, Paris – Trad. ital.: L’uomo neuronale,
Feltrinelli, Milano, 1983.
Changeux, J.-P. (2002). L’Homme de Verité. Éditions Odile Jacob, Paris – Trad. ital.: L’uomo di
verità, Feltrinelli, Milano, 2003.
Change ux, J.-P. e Connes, A. (1989), Matière à pensée, Éditions Odile Jacob, Paris – Trad. ital.:
Pensiero e materia, Bollati Boringhieri, Torino, 1991.
Changeux, J.-P. e Dehaene, S. (1989). ‘Neuronal Models of Cognitive Functions’, Cognition, 33, 63-109.
27
Sono riportati in carattere normale i testi a cui facciamo riferimento diretto e in carattere più piccolo quelli che sono
ripresi da altre fonti, come ad esempio da (Changeux, 2002) o da (Rizzolatti & Sinigaglia, 2006).
13
Cosmides, L. e Tooby, J. (1997). Evolutionary Psychology: a Primer, dal sito internet
http://www.psych.ucsb.edu/research/cep/primer.html.
Damasio, A.R. (1994). Descartes’ Error. Emotions, Reason and the Human Brain. Putnam, New
York – Trad. ital.: L’errore di Cartesio, Adelphi, Milano, 1995.
Damasio, A.R. (...). The Feeling of What Happens. Body and Emotion in the Making of
Consciousness, Harcourt Brace, NY – Trad. ital.: Emozione e coscienza, Adelphi, Milano, 2000.
Dehaene, S. (1997). La Bosse des Maths. Éditions Odile Jacob, Paris. – Trad. ital.: Il pallino della
matematica, Mondatori, Milano, 2000.
Dehaene, S.; Changeux, J.-P. e Nadal, J.-P. (1987). ‘Neural Networks that learn temporal Sequences by Selection’,
Proc. Natl. Acad. Sci. USA, 84, 2727-2731.
Dehaene, S.; Kerszberg, M. e Changeux, J.-P. (1998). ‘A Neuronal Model of a Global Workspace in effortful Cognitive
Tasks’, Proc. Natl. Acad. Sci. USA, 95, 14529-14534.
Devlin, K. (2000). The Math Gene: how mathematical Thinking evolved and why Numbers are like
Gossip. Basic Books, New York. – Trad. ital.: Il gene della matematica, Longanesi, Milano,
2002.
Duval, R. (…). …
Edelman, G. (1987). Neural Darwinism. Basic Books, NY – Trad. ital.: Darwinismo neurale: la
teoria della selezione dei gruppi neuronali, Einaudi, Torino, 1995.
Fadiga, L.; Fogassi, L.; Pavesi, G. e Rizzolatti, G. (1995). ‘Motor facilitation during action observation: a magnetic
stimulation study’, Journal of Neurophysiology, 35, 2608-2611.
Galilei, G. (1964). Opere, UTET, Torino.
Gall, F.J. (1822-1825). Sur les fonctions du cerveau et sur celles de chacune de ses parties, Baillière, Paris (6 voll.).
Gallese, V. (2000). ‘The inner sense of action. Agency and motor representation’, Journal of Consciousness Studies, 7,
10, 33-40.
Gallese, V. e Lakoff G. (2005). ‘The Brain’s Concepts: the Role of the Sensory-Motor System in
Conceptual Knowledge’, Cognitive Neuropsychology, 21, …
Gallese, V. e Metzinger, T. (2003). ‘Motor Ontology: the representational reality of goals, actions
and selves’, Philosophical Psychology, 16, 3, 365-388.
Gibson, J.J. (1979). An Ecological Approach to Visual Perception, Houghton Mifflin, Boston –
Trad. ital.: Un approccio ecologico alla percezione visiva, Il Mulino, Bologna, 1999.
Guidoni, P. (1985). ‘On natural Thinking’, Eur. J. Sci. Educ., 7, 133-140.
Guidoni, P.; Iannece, D. e Tortora, R. (2004). La formazione matematica dei futuri maestri. Appunti
ed
esempi
di
attività.
Progetto
CNR,
dal
sito
internet
http://didmat.dima.unige.it/progetti/CNR/napoli/present.html
Guidoni, P.; Iannece, D. & Tortora, R. (2005). ‘Forming Teachers as Resonance Mediators’, Proc.
of the 29th Conf. of the Intern. Group for the PME, Melbourne, 3, 73-80.
Guidoni, P.; Mellone, M. & Pezzia, M. (2005). ‘Understanding basic arithmetics by “Resonance”
approach: from addition to multiplication in first grade’, Proc. of SEMT 05, Prague.
Hawkins, D. (1974). The Informed Vision, Agathon Press, NY. – Trad. ital.: Imparare a vedere.
Saggi sull’apprendimento e sulla natura umana, Loescher, Torino, 1979.
Houdé, O. (2000). ‘L’intelligence avance de manière tout à fait bisconnue’, dal sito internet
http://www.snuipp.fr/aimprimer.php3?id_article=270.
Houdé, O.; Zago, L.; Mellet, E.; Montier, S.; Pineau, A.; Mazoyer, B. & Tzourio-Mazoyer, N.
(2000). ‘Shifting from the Perceptual Brain to the Logical Brain: the neural Impact of cognitive
Inhibition Training’, J. of Cognitive Neuroscience, 12, 721-728.
Iannece, D.; Mellone, M. & Tortora, R. (2006). ‘New insights into learning processes from some
neuroscience issues’, Proc. of the 30th Conf. of the Intern. Group for the PME, Praga, 3, 321328.
Iannece, D. & Tortora, R. (2002). ‘Un tentativo di ricostruzione del pensiero matematico nella
formazione dei maestri’, in N.A. Malara, C. Marchini, G. Navarra e R. Tortora (a cura di),
Processi didattici innovativi per la matematica nella scuola dell’obbligo, Pitagora, Bologna,
153-164.
14
Iannece, D. & Tortora, R. (2004). ‘The evolution of graphic representations in a Vygotskijan
perspective’,
Proceedings
of
CERME
3,
Bellaria,
dal
sito
internet
http://www.dm.unipi.it/~didattica/CERME3/proceedings/Groups/TG1/TG1_tortora_cerme3.pdf.
Jackson, J.H. (1932). Selected Writings (a cura di J. Taylor), Hodder and Stroughton, London.
Kohler, E.; Keysers, C.; Umiltà, M.A.; Fogassi, L.; Gallese, V. e Rizzolatti, G. (2002). ‘Hearing sounds understanding
actions: action representation in mirror neurons’, Science, 297, 846-848.
Lakoff, G. e Núñez, R. (2000). Where Mathematics comes from: how the embodied Mind brings
Mathematics into Being. Basic Books, New York. – Anche in traduzione italiana.
Laplane, D. e Dubois, B. (2001). ‘Autoactivation Deficit: A Basal Ganglia Related Sindrome’, Movement Disorders,
16, 810-814.
Lehrer, R. e Schauble, L. (…). ‘Modelling in Mathematics and Science’, in R. Glaser (ed.), Advances in instructional
Science, Lawrence Erlbaum Associates, Mahwah, NJ, vol. 5.
Leron, U. (2004). Mathematical Thinking & Human Nature: Consonance & Conflict. Proc. of the
28th Conf. of t he Intern. Group for the PME, Bergen, 3, 217-224.
Levi-Montalcini, R. (1998). L’asso nella manica a brandelli, Baldini e Castoldi, Milano.
Luccio, R. (1993). Presentazione, in (Neisser, 1976), edizione italiana, 1993, 9-16.
Marx, K. (1974). Il Capitale, VIII edizione, Editori Riuniti, Roma.
Mason, J. (2201). ‘Modelling modelling: where is the centre of gravity of- for-when teaching
modelling?’, in J.F. Matos, W. Blum, S.K. Houston, S.P. Carreira (eds.), Modelling and
Mathematics Education. ICTMA9: Applications in Science and Technology, Horwood,
Chichester, UK, 39-61.
Mason, J. (2002). ‘Learning Mathematics Mathematically: Knowledge for a New Century’, Proc. of
EARCOME 2 – SEACOME 9.
Molinier, J; Ries, G; Zipfel, C e Hohn, B. (2006). ‘Transgeneration memory of stress in plants’, Nature, 442 (7106),
1046-1049.
Neisser, U. (1976). Cognition and Reality. Principle and Implications of Cognitive Psychology,
Freeman & Co., San Francisco – Trad. ital.: Conoscenza e realtà, Il Mulino, Bologna, 1981,
1983.
Niss, M. (…). ‘Quantitative Literacy and Mathematical Competencies’, dal sito internet
http://www.maa.org/ql/pgs215_220.pdf.
Organization for Economic Cooperation and Development (OECD) (2000). Measuring Student
Knowledge and Skills: The PISA 2000 Assessment of Reading, Mathematical and Scientific
Literacy, dal sito internet http://www.pisa.oecd.org/.
Piattelli Palmarini, M. (1995). L’illusione di sapere, Mondadori, Milano.
Porter, M.I. e Raichle, M.E. (1997). Images of Mind, Scientific American Books, NY.
Rizzolatti, G.; Berti, A. e Gallese, V. (2000). ‘Spatial neglect: neurophysiological bases, cortical circuits and theories’,
in F. Boller e J. Grafman (a cura di), Handbook of Neurophysiology, 2nd edition, Elsevier, Amsterdam, 503-537.
Rizzolatti, G.; Camarda, R.; Fogassi, L.; Gentilucci, M.; Luppino, G. e Matelli, M. (1988). ‘Functional organization of
area 6 in the macaque monkey. II. Area F5 and the control of distal movements’, Experimental Brain Research, 71,
491-507.
Rizzolatti, G.; Fogassi, L. e Gallese, V. (2006). ‘Specchi nella mente’, Le Scienze, 460, 54-61.
Rizzolatti, G. e Gentilucci, (1988). ‘Motor and visual-motor functions of the premotor cortex’, in P. Rakic e W. Singer
(a cura di), Neurobiology of Neocortex, John Wiley & Sons, Chichester, 269-284.
Rizzolatti, G. e Sinigaglia, C. (2006). So quel che fai. Il cervello che agisce e i neuroni specchio,
Raffaello Cortina, Milano.
Rösken, B.; Rolka, K. (2006). A picture is worth 1000 words- the role of visualization in
mathematics learning, PME 30, 4-457.
Schoenfeld, A.H. (1991). ‘On Mathematics as Sense- making: an informal Attack on the unfortunate
Divorce of formal and informal Mathematics’, in J.F. Voss, D.N. Perkins, J.W. Segal (eds.),
Informal Reasoning and Education, Lawrence Erlbaum Associates, Hillsdale, NJ, 311-343.
Sfard A. (2000). ‘On reform movement and the limits of mathematical discourse’, Math. Thinking and
Learning 2(3), 157-189.
Sperber, D. e Wilson, D. (1986). Relevance: Communication and Cognition, Blackwell, Oxford,
Harvard University Press, Cambridge, Mass. – Trad. ital. La pertinenza, Anabasi, Milano, 1993.
15
Tafuto, R. (…). ‘Un approccio vygotskjiano alle equazioni in un Istituto Tecnico’, L’insegnamento
della matematica e delle scienze integrate, ….
Verhage, M. et al. (2000). ‘Synaptic Assembly of the Brain in the Absence of Neurotransmitter Secretion’, Science,
287, 864-869.
Verschaffel, L. (2002). ‘Taking the modelling perspective seriously at the elementary school level:
Promises and pitfalls’, Proceed. of PME XXVI, Norwich, 1, 64-80.
Verschaffel, L.; Greer, B. e De Corte, E. (2000). Making sense of word problems. Swets &
Zeitlinger, Lisse, Olanda.
Vinner, S. (1997a). ‘The pseudo-conceptual and the pseudo-analytical thought processes in
mathematics learning’, Educational Studies in Mathematics, 34, 97-129.
Vinner, S. (1997b). ‘From Intuition to Inhibition – Mathematics, Education and other Endangered
Species’, Proceed. of PME XXI, Lahti, 1, 63-78.
Vygotskij, L.S. (1934). Myšlenie i rec’. Psichologiceskie issledovanija, Mosca, 1934. – Trad. Ital.:
Pensiero e Linguaggio. Ricerche psicologiche. (a cura di L. Mecacci), Laterza, Bari, 1990.
Wason, P.C. (1983). ‘Realism and Rationality in the Selection Task’. In: Evans, J.S.T.B. (ed.)
Thinking and Reasoning, Routledge & Kegan Paul.
Zan, R. (…). ….
16