Vie sensoriali – tutte le vie sensoriali, tranne quella olfattiva,
passano per il talamo
La capacità di un animale di relazionarsi con il mondo esterno dipende
dalla sua capacità di captare ed interpretare in modo corretto le
informazioni provenienti dall’ambiente esterno e dal suo ambiente interno.
Le risposte che l’organismo sviluppa a stimoli provenienti dall’ambiente
esterno saranno appropriate solo se i dati riguardanti gli stimoli stessi
sono fedelmente codificati in segnali che i neuroni del sistema nervoso
centrale possono ricevere ed elaborare.
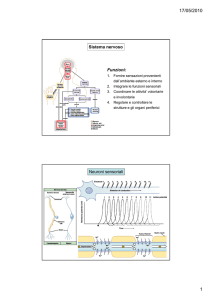
I recettori sensoriali rappresentano i canali di comunicazione tra il
mondo esterno e il sistema nervoso
Raccolgono stimoli provenienti dall’ambiente esterno. Essi sono
quindi localizzati sulla superficie dell’organismo
Raccolgono stimoli provenienti dall’ambiente interno e sono quindi
localizzati in profondità.
Gli enterocettori si distinguono in:
viscerocettori: raccolgono stimoli dai visceri per coordinare la vita
vegetativa e quindi i processi omeostatici
recettori dell’equilibrio: raccolgono informazioni sulla posizione e sui
movimenti di tutto il corpo
propiocettori: raccolgono informazioni sulle posizioni relative e i
movimenti dei vari muscoli
Gli esterocettori sono tradizionalmente classificati in recettori per la
vista, l’udito, il tatto, l’olfatto, il gusto. Occorre, però, considerare
anche tipi di informazioni sensoriali non presenti nell’uomo ma
presenti in altre specie animali.
I serpenti a sonagli sono capaci di rilevare il calore (sotto forma di
radiazioni nell’infrarosso) emesso da altri animali e possono
utilizzare questa informazione per localizzare le prede.
Alcuni pesci utilizzano segnali elettrici a frequenza molto bassa per
comunicare nell’acqua torbida. Ciò permette loro di localizzare i propri
simili.
Alcuni animali sembra che possano percepire il campo magnetico
terrestre ed utilizzarlo per l’orientamento.
Ogni stimolo, proveniente dall’ambiente interno o esterno e
percepito dall’organismo, possiede una certa quantità di
energia.
I recettori possono essere classificati in base alla
forma di energia alla quale sono selettivamente sensibili in:
chemiocettori (energia chimica)
meccanocettori (energia meccanica)
termocettori (energia termica)
fotocettori (energia luminosa)
elettrocettori (energia elettrica)
magnetocettori (energia magnetica)
I recettori sensoriali sono:
per una specifica forma di energia detta
anche modalità sensoriale
agli stimoli selezionati poiché
possono amplificare gli stimoli
che ricevono
I recettori
le entrate sensoriali in quanto
convertono l’energia contenuta negli stimoli in impulsi
nervosi
Ogni recettore è più sensibile nei confronti di uno specifico tipo di
energia. Si dice pertanto che ogni tipo di cellula recettrice normalmente
risponde solo ad uno stimolo adeguato.
Sono i meccanismi molecolari insiti nella membrana del recettore a
determinare quale sia lo stimolo adeguato di un recettore.
Le cellule fotorecettrici contengono un pigmento
visivo fatto di molecole (rodopsina) capaci di
catturare i quanti di luce; in questo processo la
struttura di tali molecole va incontro ad una
alterazione transitoria. Il pigmento stesso fa parte
della membrana cellulare, cosicché il cambiamento
conf ormazi on ale m oleco lar e p r o d u c e n e l l a
membrana una variazione di conduttanza, che
produce un segnale elettrico.
Una cellula meccanorecettrice è dotata di una membrana che risponde ad una lieve
distorsione o ad uno stiramento (possiede canali ionici che si aprono in risposta ad
uno stiramento della membrana).
chemocettore
Modelli teorici dei
meccanismi di
trasduzione in tre
diversi tipi di recettori
meccanocettore
fotorecettore
A
stimoli
B
C
Trasduttore
con sensibilità
selettiva
Cellula
recettrice
Amplificatore
di grande
potenza
Segnale nervoso
Conversione
dell'informazione nei
recettori sensoriali
Tra le molte forme di energia
che possono arrivare su un
recettore, solo una viene captata
a livello di stimolo debole o
moderato: è questa energia che
viene trasdotta e amplificata in
un segnale nervoso
()3
()2
()1
)1(
)2(
)3(
Variazioni della conduttanza di membrana
(1)L’energia dello stimolo adeguato modifica la membrana provocando l’apertura
o la chiusura di canali ionici con conseguente variazione del potenziale di
membrana. Questa variazione del potenziale di membrana è detta potenziale del
recettore
In molti recettori sensoriali vengono aperti canali cationici aspecifici che lasciano passare K+ e
Na+. Poiché il Na+ ha un gradiente elettrochimico favorevole al suo ingresso in cellula maggiore
rispetto a quello che ha il K+ per fuoriuscire dalla cellula, l’apertura di questi canali provoca un
ingresso di cariche positive che sopravanza l’uscita e una conseguente depolarizzazione della
membrana
Ambiente extracellulare
+
Na+
K
++++++++++++++++++++
--------------------------
Ambiente intracellulare
Questi canali non sono voltaggio dipendenti per cui la depolarizzazione non provoca
l’apertura di nuovi canali e non evolve in potenziale d’azione. Si aprono più o meno canali in
funzione dell’intensità dello stimolo.
Il potenziale del recettore può verificarsi secondo i casi nel senso di una depolarizzazione
(così è in genere) o nel senso di una iperpolarizzazione (es. recettori visivi). Esso è sempre
proporzionale allo stimolo, quindi è un fenomeno graduato, non ha i connotati di un
fenomeno tutto o nulla.
)1(
)2(
)3(
(2) Il potenziale del recettore si propaga solo a brevi distanze elettrotonicamente.
Se il recettore è la terminazione sensitiva di una cellula di senso primaria, oppure se è una cellula di senso
secondaria che forma sinapsi elettrica con la cellula di senso primaria, le correnti elettrotoniche si spingono
fino al primo punto della fibra nervosa capace di generare potenziali d’azione. Il potenziale nel primo punto
eccitabile della membrana viene detto potenziale generatore. Se il potenziale generatore ha un valore tale
da raggiungere la soglia per l’innesco del potenziale d’azione (ossia se il potenziale generatore è critico)
insorge il potenziale d’azione che si autopropaga, trasportando l’informazione a livello del sistema nervoso
centrale
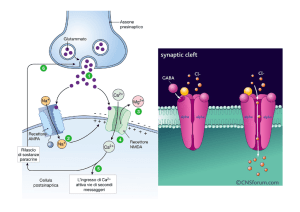
Se il recettore è una cellula di senso secondaria che forma una sinapsi chimica con la cellula di senso
primaria, le correnti elettrotoniche generate dal potenziale del recettore si arrestano alla membrana
presinaptica. Qui provocano ingresso di Ca2+ in cellula e conseguente esocitosi del neurotrasmettitore,
che, liberato nello spazio sinaptico, si lega alle molecole recettrici postsinaptiche, provocando la
formazione di un potenziale postsinaptico. Questo, mediante correnti elettrotoniche, evoca un potenziale
generatore nel primo punto capace di generare potenziali d’azione. Se il potenziale generatore è critico
insorge il potenziale d’azione autopropagato.
Sequenza di eventi nella trasduzione sensoriale
STIMOLO
Modificazioni strutturali di
canali di membrana
1: la corrente del recettore nasce nella zona
sensitiva e si propaga elettrotonicamente
depolarizzando la zona d’inizio degli impulsi.
Le cellule recettrici dispongono di una fibra
sensitiva afferente.
Variazioni di conduttanza
Corrente del recettore
Potenziale del recettore
Frequenza
degli impulsi
modulata
nell’assone
della cellula
recettrice
Rilascio modulato
di mediatore da
parte della cellula
recettrice
Frequenza degli impulsi
modulata dal neurone di
2° ordine
2: interposizione di una sinapsi tra la zona
sensitiva del recettore e una fibra afferente
secondaria
Il potenziale del recettore prodotto
nelle terminazioni del neurone
afferente (sensitivo) è una
traduzione analogica dello stimolo
Esso diffonde attraverso il soma e
fa nascere nell’assone dei potenziali
d’azione tutto-o-nulla.
Questi si propagano lungo l’assone
e causano il rilascio di un mediatore
che fa nascere, nel neurone
successivo, un potenziale sinaptico
graduato che potrà eventualmente
generare un nuovo pot. d’azione o
un treno d’impulsi.
Risposte elettriche a stimoli deboli e forti si traducono in
variazioni di frequenza di scarica dei potenziali d’azione
)1(
)2(
)3(
(3) Il potenziale d’azione, che si autopropaga, trasporta l’informazione sensoriale
al sistema nervoso centrale
I singoli potenziali d’azione che portano l’informazione proveniente dai diversi organi di
senso sono sostanzialmente indistinguibili tra loro. In altre parole la modalità dello
stimolo non è codificata da alcuna caratteristica intrinseca dei potenziali stessi, ma
piuttosto dalla specificità anatomica della connessione che quel dato neurone stabilisce
con il cervello.
L’informazione relativa all’intensità dello stimolo viene codificata come frequenza
dei potenziali d’azione.
La frequenza dei potenziali d’azione è proporzionale al logaritmo dell’intensità dello
stimolo (legge di Weber e Fechner) :
E = a logI - b
E = effetto biologico (frequenza dei potenziali d’azione)
a e b sono costanti
logI è il logaritmo dell’intensità dello stimolo
Rapporti fra segnale in entrata
e in uscita negli organi di
senso
A: in molti recettori l’ampiezza del
potenziale del recettore è in
relazione lineare con il logaritmo
dell’intensità dello stimolo, per un
esteso campo di valori
B: La frequenza degli impulsi è in
relazione lineare con l’ampiezza del
potenziale del recettore
C: come risultato di A e B la
frequenza degli impulsi varia
linearmente col logaritmo
dell’intensità dello stimolo
Le risposte dei recettori sensoriali presentano diversi gradi di adattamento, in
relazione al tipo di informazione che deve essere trasmessa:
1) recettori tonici sono a lento
adattamento, continuano a rispondere
dando una scarica d'impulsi oer tutta la
durata di uno stimolo costante
2) recettori fasici si adattano
rapidamente a uno stimolo costante e si
inattivano; alcuni di questi rispondono
solo a variazioni dell'intensità di
stimolazione
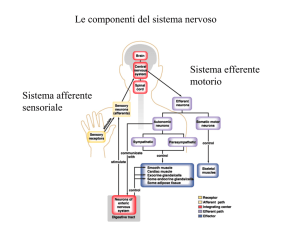
Sistema sensoriale generale o Somatoviscerale
Meccanocettori
Termocettori
Nocicettori
I recettori sensoriali cutanei comprendono tutti i tipi di recettori
I Muscoli scheletrici contengono Recettori da stiramento e nocicettori
Le articolazioni e i visceri sono innervati da meccanocettori e nocicettori
Sistemi sensoriali Speciali
Organi sensoriali specifici a livello cefalico connessi con appropriati sistemi
nervosi cerebrali
Visivo
Olfattivo
Uditivo
Gustativo
Vestibolare
I meccanorecettori piu semplici sono terminazioni
nervose morfologicamente indifferenziate presenti
nel connettivo della pelle
In alcuni recettori si sono evolute strutture accessorie il cui ruolo é quello di
realizzare un efficiente trasferimento di energia alla cellule recettrice
Meccanocettori a rapido adattamento
Meccanocettori a lento adattamento
Le differenze di sensibilità agli stimoli meccanici nelle varie parti della
superficie del corpo
Campi Recettivi dei neuroni sensoriali
La convergenza di diversi neuroni sensoriali primari permette a
stimoli simultanei sottosoglia di sommarsi a livello del neurone
secondario e quindi di innescare un potenziale d’azione
Gemme gustative con fibre sensitive secondarie
che innervano le cellule recettrici primarie, in
viola. In bianco le cellule di supporto
Le gemme gustative contengono sino a 150
cellule recettrici in associazione con cellule di
sostegno e basali che ricambiano in
continuazione le cellule recettrici; le membrane
apicali delle cellule recettrici possiedono
microvilli che sporgono in un poro gustativo
dove entrano in contatto con la saliva
Meccanismo di trasduzione del segnale
1) le molecole recettrici localizzate sui microvilli
riconoscono le sostanze chimiche presenti nella
saliva
2) variazioni conduttanza di membrana
3) genesi del potenziale del recettore
4) liberazione di un neurotrasmettitore
eccitatorio
5) comparsa di un potenziale generatore nella
fibra afferente primaria
6) scarica di impulsi nervosi nelle fibre afferenti
primarie dirette al SNC