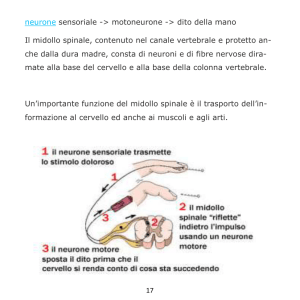
1. Il neurone
Il cervello ha più compiti, tra cui:
– tradurre le informazioni del mondo esterno, trasformando le informazioni (visive, uditive,
olfattive...) in codice neurale, ovvero in qualcosa che i neuroni possono scambiarsi tra loro. I neuroni
variano la propria attività elettrica. Di questo si occupano i recettori. Una volta trasformata,
l'informazione è elaborata e integrata dai neuroni, costruendo così la realtà. Le informazioni,
dapprima scomposte, vengono poi associate ed eventualmente anche memorizzate;
– pianificare nuove azioni;
– inviare informazioni ai muscoli, determinando un'azione o un comportamento (regolazione della
risposta motoria).
Gli invertebrati, che non possiedono SN (sistema nervoso) sono totalmente in balia degli eventi esterni. Solo
nelle meduse, invertebrati, compaiono i primi neuroni che svolgono i compiti sopracitati, potendo perciò, per
esempio, riconoscere eventuali prede e difendersi. Il SN in esse è semplice, composto da una serie di neuroni
che formano una rete. Dalle meduse arriviamo a invertebrati più complessi, in cui i neuroni si raggruppano in
gangli, da cui esce un cordone nervoso.
Le cose diventano più complesse quando neuroni differenti si uniscono in gangli differenti, come negli
insetti, in cui abbiamo la comparsa di recettori sensoriali diversi.
Nei vertebrati abbiamo una struttura di base molto simile. I neuroni si raggruppano per formare l'encefalo e
le strutture sono protette da un sistema osseo.
Nei mammiferi e nei primati abbiamo l'aumento delle dimensioni del cervello (associato all'aumento delle
dimensioni corporee). A questo è correlato l'aumento della complessità delle connessioni tra neuroni. Si
sviluppano inoltre zone che svolgono determinate funzioni molto complesse.
Abbiamo due tipi di cellule nel SN: i neuroni e le cellula gliali (glia). Le cellule gliali hanno funzione di
sostegno, alcuni di essi formano la guaina mielinica, captano e smaltiscono le molecole e regolano lo
sviluppo dei neuroni. Le famiglie sono due: microglia e macroglia. Le prime sono piccole e distruggono le
molecole di rifiuto. Abbiamo tre tipi di cellule glia:
– astrociti (che si trovano sia nel SNC che nel SNP);
– oligodendrociti (che si trovano solo nel SNC). Sono in grado ci contattare più neuroni;
– cellule si Shwann (che si trovano solo nel SNP). Sono in grado di avvolgersi attorno a un unico
neurone.
Gli astrociti devono il loro nome alla loro forma, a stella. Essi formano la barriera ematoencefalica.
L'ambiemte in cui si trovano i neuroni deve essere mantenuto costante. Questa barriera regola la quantità e il
tipo di sostanze nel passaggio sangue-neurone. La barriera è formata dagli astrociti e dai capillari, uniti tra
loro tramite le giunzioni serrate. Ha funzione selettiva; ci sono però alcune parti in cui lo è di meno, cosicchè
sia possibile consentire al cervello di monitorare la concentrazione degli ormoni nel sangue o per liberare
sostanze nel sangue.
I neuroni sono formati dagli assoni, che possono essere essere ricoperti da guaia mielinica. Le sinapsi sono
punti di contatto tra neuroni.
Per il principio della polarizzazione dinamica, le diverse parti del neurone svolgono funzioni diverse con una
direzione ben precisa, che parte dai dendriti e arriva alle terminazioni.
I dendriti ricevono le informazioni, che confluiscono nel corpo cellulare, dove vengono integrate tra loro. Se
il neurone decide di rispondere, l'informazione esce, percorre l'assone, che ha proprio questa funzione, cioè
di portare l'informazione dal corpo cellulare al neurone stesso. I dendriti sono ramificati; possiedono le spine
dendritiche, prolungamenti che formano una sinapsi. Le spine aumentano la superficie di contatto e quindi
anche la capacità di ricevere le informazioni. Sono elementi plastici.
Il corpo cellulare o soma integra le informazioni, contiene il DNA nel nucleo, in cui avviene la sintesi
proteica (precisamente nel citoplasma). Le proteiene sintetizzate raggiungono le terminazioni dell'assone,
movimento regolato dal citoscheletro. Le proteine vengono impacchettate in vescicole e trasportate dai
microtubuli. Ci sono due tipi di trasporto: lento e rapido. Quello lento viene usato per le proteine di ricambio
dell'assone stesso, quello rapido è usato per inviare le proteine alle terminazioni neurali.
Il neurone possiede un unico assone, che può essere molto lungo o molto corto. La sua funzione è quella di
condurre i segnali elettrici, dal corpo cellulare alle strutture bersaglio. La guaina mielinica aumenta la
capacità dell'assone di condurre le informazioni. Si ramifica nel terminale dell'assone.
Se ce l'ha, nel terminale l'assone perde la guaina mielinica.
I neuroni possono essere classificati in tre modi:
– a seconda del numero di prolungamenti, il neurone può essere unipolare (una propaggine), bipolare,
pseudo-unipolare (a T), multipolare (unico assone, tante propaggini);
– a seconda della funzione. I neuroni sensitivi partecipano all'acquisizione di stimoli, trasportando le
informazioni dagli organi sensoriali al sistema nervoso centrale.
Gli interneuroni, all'interno del sistema centrale, integrano i dati forniti dai neuroni sensoriali e li
trasmettono ai neuroni motori. I motoneuroni emanano impulsi di tipo motorio agli organi della
periferia corporea. Nel SNC abbiamo gli interneuroni locali e i neuroni a proiezione, così chiamati
perché proeittano al di fuori della struttura in cui si trovano.
2. La membrana neuronale
I neuroni, come tutte le altre cellule, sono completamente avvolti dalla membrana cellulare, in questo caso
chiamata neuronale. La membrana neuronale ha un doppio strato fosfolipido, con teste polari e code apolari,
disposte con la parte polare rivolta verso l'esterno e l'interno e la parte polare nel mezzo. Come tutte le altre
cellule, i neuroni sono formati da glucidi, lipidi, protidi e acidi nucleici. Gli elementi principali che li
compongono sono principalmente l'idrogeno, l'ossigeno, il carbonio e l'azoto.
Vediamo come passano le sostanze attraverso la membrana. Il neurone è infatti costantemente immerso nella
struttura cellulare e scambia con esso costantemente sostanze. Il passaggio delle sostanze dipende da due
fattori fondamentali:
– la carica elettrica: ci sono sostanze che portano una carica elettrica (positve o negative) e altre che
invece sono neutre. Le sostanze neutre passano più facilmente dalla membrana neuronale, come il
glucosio (fondamentale per il metabolismo dei neuroni), ossigeno e anidride carbonica (che oltre ad
essere neutri sono anche piccoli). Ci sono sostanze come il cloruro di sodio che invece presentano
cariche elettriche e fanno molta fatica a passare, perchè le teste polari tendono a impedire il
passaggio delle sostanze con carica elettrica;
– la dimensione: tanto più grandi sono le sostanze, tanto più difficilmente attraverseranno la
membrana. Il glucosio, nonostante sia neutro, è grande, e non riesce da solo ad attraversare la
membrana. Le sostanze che non riescono a passare facilmente o non riescono proprio a passare
passano attraverso proteine trasportatrici, che si trovano nella membrana del neurone e consentono il
trasporto delle sostanze, che legano e le trasprotano all'interno o all'esterno.
Questo tipo di strutture specializzate hanno due caratteristiche fondamentali:
– la specificità: sono estremamente specifiche, ci sono cioè proteine che legano e riconoscono solo ed
esclusivamente il glucosio, altre che invece fanno passare, per esempio, il sodio, e così via. Abbiamo
tante strutture diverse specifiche;
– la modulazione allosterica: la maggior parte delle proteine va incontro a modulazione allosterica,
cioè quando la nostra sostanza si lega alla proteina, la proteina va incontro a una modifica della
propria struttura. Grazie a questo cambiamento di struttura la protiena diventa in grado di far passare
quella determinata sostanza.
Queste proteine possono essere di diverso tipo. Un primo tipo sono i trasportatori, che
si incaricano fisicamente di trasportare le sostanze dall'interno all'esterno e viceversa,
ad es: il trasportatore per il glucosio.
Ci sono poi le proteine-canale, le quali sono sempre proteine che attraversano la
membrana, ma in questo caso le proteine formano al loro interno un canale, che
consente il passaggio di determinate sostanze. Questo canale può essere o aperto o
chiuso, quindi ci saranno momenti in cui le proteine non consentono il passaggio (chiuso) e altri in cui lo
faranno (aperto). Questi canali fanno passare le sostanze con carica elettrica. Le proteine presentano un filtro
selettivo, che presenta cariche di segno opposto alla sostanza in modo da attrarle. I canali sono formati da
tante proteine unite assieme. Ci sono canali sensibili a stimoli di natura chimica e quindi quando arriverà una
determinata sostanza (es: nei neurotrasmettitori) si legano ai canali sensibili, così il canale si apre e fa
passare una determinata carica elettrica. In questo caso si parla di canali ionici a porta chimica. Abbiamo poi
dei canali sensibili a sostanze di natura elettrica (canali ionici a porta elettrica). Esistono anche canali ionici
sensibili a stimoli meccanici o termici.
Le forze che regolano il passaggio delle sostanze attraverso la membrana sono due: il flusso di volume e il
flusso di diffusione. Il flusso di volume riguarda una notevole quantità di particelle (flusso d'acqua di un
fiume) in una certa direzione. A noi interessa il flusso di diffusione, perchè riguarda una quantità di particelle
molto minore. La diffusione è fondamentalmente determinata dal passaggio di molecole, che tendono a
muoversi casualmente, non hanno perciò una direzione ben precisa. Inoltre il flusso di diffusione dipende
dalla concetrazione di molecole presenti. In generale si muoverà da dove le molecole sono più concentrate a
dove sono meno concentrate. Il primo tipo di diffusione è la diffusione semplice: la quantità di particelle
determina il loro passaggio, cioè tenderanno a muoversi secondo gradiente di concentrazione, da dove sono
meno concentrate a dove lo sono di più. Questo tipo di diffusione non richiede consumo di energia.
Un secondo tipo di diffusione è quella facilitata, come nel caso del glucosio. Anche in questo caso dipende
dal gradiente di concentrazione, ma essendo il glucosio non libero di passare dalla membrana, abbiamo
l'intervento di proteine trasportatrici. I principi di base sono simili a quelli della diffusione (ci si basa sul
gradiente), ma in questo caso sono presenti le proteine trasportatrici e proprio per la presenza di queste, se c'è
una quantità di sostanza maggiore rispetto alla quantità di proteine, una certa parte di sostanza rimarrà fuori.
Abbiamo perciò la saturazione del sistema. Anche in questo caso non c'è bisogno di consumo di energia.
Il terzo tipo di diffusione è il trasporto attivo, che invece avviene contro gradiente di concentrazione. In
questo caso la sostanza verrà trasferita da dove ce n'è poca a dove ce n'è molta. Lavorando contro il gradiente
di concentrazione, si consuma energia. È il caso della pompa sodio-potassio.
Uniporto: trasporto di un'unica sostanza
Simporto: trasporto di due sostanze nella medesima direzione
Antiporto: trasporto di due sostanze in direzioni diverse
Come funzionano i neuroni?
Tutti i neuroni funzionano elaborando segnali di natura elettrica. Nel neurone a riposo (cioè non stimolato),
ci sono determinate sostanze più concentrate all'esterno e determinate sostanze più concentrate all'interno del
neurone, con una differente distribuzione. In particolare queste sostanze portano cariche elettriche. Per
definizione gli atomi o le molecole che portano cariche elettriche si chiamano ioni. Gli ioni principali sono il
sodio (Na+), il potassio (K+), il calcio (Ca++) e il cloro (Cl-). Oltre a questi ci sono struttre proteiche che
portano cariche negative, chiamati anioni proteici, indicati col termine generico A-.
A riposo, all'esterno del neurone c'è un'elevata concentrazione di sodio, calcio e cloro. All'interno: potassio e
anioni proteici. Proprio perchè gli ioni hanno una carica elettrica, non passano normalmente dalla membrana,
quindi nel neurone a riposo, se non interviene nulla, il sodio resterà fuori e il potassio dentro. Questa
differenza di ioni è creata e mantenuta grazie alle pompe metaboliche, proteine di membrana che operano
contro il gradiente di concentrazione dello ione (trasporto attivo). Un esempio di pompa metabolica è la
pompa sodio-potassio, che ad ogni ciclo porta all'esterno 3 ioni sodio e all'interno 2 ioni potassio. La pompa
lega il sodio alla proteina, in seguito al legame tenderà a buttarlo fuori, lavorando contro gradiente e quindi
consumando energia. Il meccanismo è una modificazione allosterica, cioè avviene modificando la struttura
della proteina. La stessa cosa avviene per il potassio. Ci sono anche pompe per il calcio o per il cloro.
La differente concentrazione di ioni tra interno e esterno del neurone produce una differente distribuzione di
cariche elettriche (positive e negative) tra esterno e interno. La presenza di cariche positive e negative
separate produce una differenza di potenziale elettrico (V, misutato in Volt). Questo potenziale elettrico della
membrana neuronale a risposo è detto potenziale di membrana a riposo, Vm.
Per passare dalla membrana, gli ioni hanno bisogno di canali ionici. Nel neurone a riposo abbiamo dei canali
ionici sempre aperti, chiamati per questo passivi. Vediamo cosa succede quando il canale ionico per il
potassio è aperto: il potassio quindi esce, seguendo il proprio gradiente di concentrazione. Le cose si
complicano perché il potassio ha anche una carica (positiva). Man mano che il potassio esce, escono quindi
anche delle cariche positive. A questo punto, all'esterno del neurone inizieranno ad accumularsi delle cariche
positive. Con la continua uscita di potassio, aumenterà la quantità di cariche positive che escono,
generandosi così un gradiente elettrico, che in questo caso farà sì che l'interno diventi più negativo
dell'esterno. La negatività del neurone tenderà dunque ad attrarre le cariche positive che erano uscite.
Contemporaneamente, avremo due flussi di ioni potassio in due direzioni opposte: un primo flusso segue il
gradiente di concentrazione e andrà dall'interno all'esterno; un secondo flusso avrà direzione opposta. Il
primo segue il gradiente di concentrazione, mentre il secondo il gradiente elettrico. Si arriverà ad un
equilibrio in cui la quantità di ioni che esce è esattamente uguale ed opposta alla quantità di ioni che
rientrano. Questo equilibrio è elettrochimico, in quanto si basa su due diverse forze, una di natura chimica ed
una di natura elettrica. In base all'equazione di Nerst, è possibile calcolare il valore di equilibrio per il
potassio EK+ = -90 mV.
La stessa cosa avviene per il sodio. In questo caso E Na+ = +55 mV.
Nel neurone a riposo ci sono sia canali per il potassio sempre aperti che canali per il sodio sempre aperti,
quindi i due passaggi avvengono contemporaneamente. Generalmente però i canali per il potassio sono in
numero maggiore che per il sodio. Il valore del potenziale di riposo della membrana neuronale Vm = -70 mV.
Il valore è quindi più vicino a -90 (equilibrio del potassio) che a quello del sodio (+55), in quanto il neurone
è più permeabile agli ioni potassio che a quelli sodio.
[Tutti e tre i valori degli equilibri sono da sapere per l'esame]
3. Potenziali locali e potenziale d'azione
Vediamo cosa succede quando un determinato stimolo, di qualunque natura, attiva un neurone. Nel neurone a
riposo abbiamo il potenziale di membrana a circa -70 mV con tanto calcio e cloro fuori e tanto potassio
dentro. Supponiamo che uno stimolo apra il canale per il sodio. Una volta aperto, il sodio tenderà a entrare,
in base al proprio gradiente di concentrazione. Aumenteranno le cariche positive nel neurone, in quanto il
sodio porta cariche positive. Entrando cariche positive, il potenziale di membrana diventerà più positivo.
Tutte le volte che il nostro neurone, in conseguenza dell'apertura di un canale, diventa più positivo, si parla di
depolarizzazione.
Se un altro stimolo determina l'apertura di un canale per il potassio, anch'esso seguirà il proprio gradiente di
concentrazione e tenderà a entrare, ma in questo caso entreranno cariche negative. In questo caso il
potenziale di membrana, da -70 mV diventerà ancora più negativo. In generale tutte le volte che il potenziale
di membrana di un neurone a seguito dell'apertura di un canale diventa più negativo si parla di
iperpolarizzazione.
Possiamo avere depolarizzazione del neurone anche quando escono cariche negative: aumenta la positività
del neurone stesso. Abbiamo un'iperpolarizzazione sia quando entrano cariche negative che quando escono
cariche positive. Così il neurone risponde agli stimoli.
Quando abbiamo un passaggio di cariche elettriche si parla di corrente elettrica, misurata in ampere (amp).
Col termine conduttanza elettrica (g) si indica la misura della facilità con cui uno ione può attraversare la
membrana cellulare e si misura in siemens (S). L'opposto della conduttanza è la resistenza (R), cioè la
dfficoltà con cui uno ione può attraversare la membrana; si misura in ohm (Ω).
I nostri neuroni integrano tutte le informazioni che ricevono con i meccanismi di depolarizzazione e
iperpolarizzazione. Una prima caratteristica di questo tipo di meccanismi è il fatto di avere ampiezza
variabile, cioè, se arriva una stimolo debole, si aprirà un numero piccolo di canali per il sodio e avremo
quindi una piccola depolarizzazione. Se lo stimolo sarà intenso, la depolarizzazione sarà più ampia.
Una seconda caratteristica è che le depolarizzazioni e le iperpolarizzazioni sono sommabili. Oltre ad essere
sommabili, si possono anche sottrarre tra loro. Se abbiamo una depolarizzazione e una iperpolarizzazione,
alla prima si sottrae la seconda.
Una terza catteristica di questi fenomeni è il fatto che tendono a propagarsi nelle zone vicine. Una volta
innescata una corrente elettrica, questa tende a spostarsi (propagarsi) nelle zone vicine. Questo tipo di
propagazione incontra due diversi tipi di resistenze. Il primo tipo di resistenza è quella chiamata interna: il
neurone contiene il citoplasma che si oppone come un muro al passaggio delle correnti elettriche. Il secondo
tipo è determinata dalla presenza di canali ionici che fanno sì che queste cariche elettriche si disperdano,
rallentandone il passaggio: parliamo di resistenza di membrana.
Queste due resistenze, opponendosi al passaggio di depolarizzazione e iperpolarizzazione, fanno sì che
queste non vadano molto lontano: l'ampiezza della corrente elettrica diminuisce con l'aumentare della
distanza dal punto di insorgenza. Iperpolarizzazione e depolarizzazione sono chiamati per questo motivo
potenziali locali, perché non vanno a distanza. Caratteristiche dei potenziali locali: vengono condotti solo
per pochi millimetri, perché l'ampiezza tende a diminuire con la distanza fino ad esaurirsi completamente.
Riassumento: i potenziali locali hanno ampiezza variabile, sono sommabili e sottraibili. Inoltre sono passivi,
cioè non devono essere formati nuovamente.
I potenziali locali vengono usati per un'infinità di funzioni: per tradurre la realtà esterna, trasformando
stimoli di qualunque tipo in codici neurali; per ricevere le informazioni dagli altri neuroni attraverso le
sinapsi; per elaborare e integrare tra loro tutto queste informazioni.
I potenziali locali non vanno a distanza; quindi come condurre le informazioni per lunghe distanze? Oltre a
dover inventare un modo per inviare le info a lunga distanza, il cervello ha dovuto inventare anche un modo
affinchè le informazioni non fossero sensibili alle due resistenze. Il poteziale d'azione è il modo attraverso il
quale i neuroni trasferiscono gli stimoli per una lunga distanza. La caratteristica di base del potenziale
d'azione è quella di avere l'ampiezza sempre uguale indipendentemente dalla distanza che percorre. Il
potenziale d'azione si basa sul funzionamento di due diversi tipi di canali, sensibili a stimoli di natura
elettrica, quindi propriamente chiamati voltaggio-dipendenti. Abbiamo due diversi tipi di canali ionici: canali
per il sodio e canali per il potassio. La caratteristica principale di questi canali è che sono estramemente
specifici (il canale per il sodio fa passare solo ed esclusivamente il sodio, idem per il potassio). I canali per il
sodio sono molto rapidi ad aprirsi e chiudersi, quelli per il potassio invece sono più lenti. I canali per il sodio
possono essere presenti in tre diversi stati: stato aperto, stato chiuso e stato inattivato (diverso da quello
chiuso, ma anche in questo stato, come in quello chiuso, lo ione non passa). L'apertura del canale per il sodio
è determinata dalla depolarizzazione. In maniera analoga abbiamo i canali per il potassio; esistono nello stato
aperto e chiuso; sono anch'essi aperti dalla depolarizzazione.
Come funziona il potenziale d'azione? Arriva uno stimolo (interno o esterno che sia); la depolarizzazione
apre i canali per il sodio voltaggio-dipendenti (che sono più sensibili e più rapidi di quelli per il potassio). Se
la depolarizzazione ha ampiezza piccola, si apre un numero limitato di canali per il sodio voltaggiodipendenti, il sodio entra e il fenomeno si estingue. Quando però arriva una depolarizzazione particolarmente
intensa, si aprirà un numero di canali per il sodio elevati; il sodio entra e la depolarizzazione è molto intensa.
Questa depolarizzazione tenderà ad aprire gli altri canali per il sodio voltaggio-dipendenti che si trovano
nelle zone vicine. Si attiva dunque un meccanismo (feedback positivo) che fa sì che si aprano tutti i canali
per il sodio voltaggio-dipendenti presenti in quella zona; così il neurone si depolarizza enormemente.
Abbiamo la cosiddetta fase ascendente del potenziale d'azione. In termini numerici, durante la
depolarizzazione, andiamo da -70 mV a valori sempre più positivi. Per insorgere, il potenziale d'azione ha
bisogno di una depolarizzazione particolamente forte, fino a quando tutti i canali per il sodio si aprono e il
neurone si depolarizza completamente. Chiamiamo questo insieme di meccanismi fase ascendente del
potenziale d'azione. Se ci fosse soltanto sodio, il potenziale d'azione arriverebbe fino a +55 mV (valore del
sodio in equilibrio). Il picco di questa fase dovrebbe infatti arrivare a +55 mV, ma questo non succede perchè
nel frattempo hanno iniziato ad aprirsi i canali per il potassio, più lentamente, e perciò avviene l'affetto
opposto di iperpolarizzazione, fermandoci a +30 mV. Il vantaggio del potenziale d'azione è l'ampiezza è la
stessa e va da -70 a +30 mV, quindi l'ampiezza sarà più o meno di 100 mV. Ciò vuol dire che è insensibile
alle resistenze e non varia con la distanza.
Essendo i canali del sodio rapidi, una volta arrivati al valore massimo, essi passano allo stato inattivato e non
fanno più passare il sodio. Contemporaneamente si stanno aprendo i canali per il potassio, che tenderà ad
uscire; uscendo, esce una carica positiva, aumenterà la negatività all'interno del neurone determinando
un'iperpolarizzazione. Abbiamo quindi la fase discendente del potenziale d'azione: dal valore estremamente
positivo di +30 mV, con l'uscita di ioni K, il neurone diventa sempre più negativo, arrivando a un valore di
-90 mV (in quanto il valore di equilibrio del K è -90 mV). A questo punto i canali per il potassio si chiudono
e la situazione viene poi riportata nella norma; il neurone è pronto per fare un nuovo potenziale d'azione.
Il potenziale d'azione ha una soglia per essere indotta. In generale la soglia per indurre il potenziale d'azione
oscilla intorno ai -67, -65 mV. Se è poco intensa e non raggiunge quel valore il potenziale rimane locale.
Il potenziale d'azione, riassumendo: viene innescato da stimoli di natura elettrica, precisamente una
depolarizzazione; non tutte le depolarizzazioni sono in grado di determinare un potenziale d'azione, ma solo
quelle che raggiungono la soglia; una volta raggiunta, il potenziale d'azione innesca una serie di reazioni che
portano molto rapidamente a una depolarizzazione molto intensa. Si aprono poi i canali per il potassio, la sua
uscita e la fase discendente carattrizzata da iperpolarizzazione. Il potenziale d'azione è un fenomeno tutto o
nulla, cioè insorge nella sua ampiezza massima; al di sotto dell'azione il potenziale non si forma.
Come faccio a conoscere l'intensità degli stimoli in termini di potenziale d'azione? Come fanno i neuroni a
tradurre l'intensità? Attraverso il codice di frequenza: un modo attraverso il quale i neuroni traducono
l'intensità degli stimoli. Essendo l'intensità costante, facciamo un maggior numero di potenziali d'azione, di
quantità maggiore.
I neuroni traducono gli stimoli che ricevono in potenziali locali (iperpolarizzazione e depolarizzazione).
Questi potenziali locali possono essere nei dendriti e (in questo caso) in generale arriveranno al corpo
cellulare, che integra queste informazioni. Ciò vuol dire sommare tra di loro le depolarizzazioni, le
iperpolarizzazioni e sottrarre tra loro depolarizzazione e iperpolarizzazione. Il risultato finale di tutto questo
processo di elaborazione è che: se viene raggiunta la soglia, il neurone forma il potenziale d'azione e
risponde agli stimoli che ha appena elaborato. Il potenziale d'azione si forma principalmente tra il corpo
cellulare e l'assone (immagine).
Si muoverà lungo l'assone, per il principio della polarizzazione dinamica.
Differenza tra potenziali locali e potenziali d'azione. I potenziali locali:
– sono condotti solo per pochi mm;
– hanno ampiezza variabile;
– sono sommabili;
– decrementano con la distanza;
– sono passivi.
I potenziali d'azione:
– sono condotti per lunghe distanze;
– hanno ampiezza costante;
– non sono sommabili;
– non decrementano con la distanza;
– sono attivi.
Cono di emergenza o zona di innesco: punto preferenziale in cui si forma il potenziale d'azione, tra corpo
cellulare e assone. In questa zona l'insorgenza del potenziale d'azione è facilitato dai tanti canali per il sodio
voltaggio-dipendenti.
Effetto dell'iperpolarizzazione: l'iperpolarizzazione non è in grado di aprire nè i canali per il sodio nè quelli
per il potassio, ma è importante perchè diminuisce la probabilità di insorgenza del potenziale d'azione perchè
lo allontana dalla soglia, rendendolo ancora più negativo.
4. Propagazione del potenziale d'azione
Nello stato inattivato i canali del sodio non sono sensibili alle depolarizzazioni. Mentre se sono chiusi, il
canale è comunque sensibile alle depolarizzazioni; arrivando, la depolarizzazione apre il canale. Nel picco la
maggior parte dei canali del sodio sono insensibili alle depolarizzazioni. A questo punto, poichè i canali per il
sodio sono inattivati, quando arrivano nuove depolarizzazioni, il neurone non è in grado di fare un nuovo
potenziale d'azione. Si parla di periodo refrattario assoluto. Il periodo refrattario inizia già durante la fase
ascendente (in rosso): in questo caso non succede nulla perchè i canali del sodio sono già impegnati a fare
potenziale d'azione. Dall'altra parte (in verde) non è possibile fare un nuovo potenziale d'azione perché i
canali per il sodio sono disattivati. Durante la fase discendente i canali, da disattivati, passano allo stato
chiuso divenendo sensibili ad altre depolarizzazioni. Se durante questa fase arriva una depolarizzazione,
questa dev'essere abbastanza forte da superare la negatività di -70 mV. Se questo avviene, si può riprodurre
un nuovo potenziale d'azione. Si parla di periodo refrattario relativo.
Effetto del periodo refrattario sull'andamento del potenziale d'azione: il potenziale d'azione insorge nel collo
d'emergenza. La depolarizzazione è una corrente elettrica che si muove e si propaga lungo l'assone. La
depolarizzazione si propaga nelle zone adiacenti e determina l'insorgenza di un nuovo potenziale d'azione.
Successivamente il processo è innescato e continua a propagarsi lungo i neuroni. Il potenziale d'azione è
unidirezionale grazie al periodo refrattario. Anche avanzando a lunghe distanze, il potenziale d'azione ha
sempre la stessa ampiezza.
Quando innestiamo un potenziale d'azione, questo genera una depolarizzazione che si muove lungo l'assone.
Lungo l'assone la depolarizzazione incontra due ostacoli (resistanza di membrana e resistenza interna).
Ci sono stati, nel corso dell'evoluzione, diversi tentativi di aumentare la
velocità di conduzione dei segnali elettrici durante. Negli invertebrati è
stato aumentato il diametro dell'assone, in quanto diminuisce la
resistenza che il citoplasma oppone al passaggio della corrente e la
corrente passa più rapidamente.
Per quanto riguarda i mammiferi, l'evoluzione non ha seguito questi
tentativi: c'è il vantaggio di aumentare la velocità, ma c'è lo svantaggio dell'aumento della massa corporea e
consumeremmo troppa energia. Il modo più semplice per aumentare la velocità nei vertebrati è avvolgere i
neuroni di una guaina isolante, che blocca i canali ionici, impedendo la fuoriuscita delle cariche. Questa
guaina è la guaina mielinica, che non è prodotta dall'assone stesso, ma dalle cellule gliali. La resistenza di
membrana aumenta dunque enormemente. Nel nostro assone abbiamo zone ricoperte di guaina isolante, ma
non è possibile avvolgere tutto l'assone, altrimenti la corrente elettrica che si forma nel cono d'emergenza
non riuscirebbe a passare fino alla meta. Quindi bisogna lasciare zone in cui non è presente guaina mielinica.
In queste zone si può riformare il potenziale d'azione. Queste zone nude si chiamano nodi di Ranvier, che
servono proprio al potenziale d'azione di riformarsi. La conduzione del potenziale d'azione è dunque
saltatoria. Nelle fibre mieliniche il potenziale d'azione tende apparentemente a saltare da un nodo di Ranvier
all'altro. Grazie a questo meccanismo la corrente elettrica aumenta enormemente la velocità di conduzione
del segnale elettrico. Per poter riformare il potenziale d'azione, nei nodi di Ranvier sono presenti canali per il
sodio e per il potassio.
Per usare dei termini tecnici, si dice che la velocità di conduzione, indicata con λ dipende da resistenza
interna e resistenza di membrana; in particolare, la velocità di conduzione aumenta con l'aumentare della
resistenza di membrana e con il diminuire della resistenza interna.
λ = mr / mi
Nell'encefalo abbiamo alcuni neuroni con guaina mielinica e altri senza; quelli con guaina mielinica hanno
diametro differente e quindi condurranno con velocità differenti. Le fibre più rapide sono quelle di tipo A.
Quelle di tipo C saranno quelle senza guaina mielinica, cioè le più lente.
Nella sclerosi abbiamo perdita di guaina mielinica, percui la conduzione del segnale tende a rallentarsi.
Essendo una fibra fatto per portare i segnali elettrici con conduzione saltatoria, questo causerà problemi
anche ad altre componenti.
Nel regno animale troviamo una quantità di soggetti, vertrebrati e non, che si sono inventati sostanze in
grado di bloccare canali ionici per il passaggio del potenziale d'azione; il pesce palla, per esempio, produce
una sostanza che si lega ai canali ionici e li blocca, paralizzando le proprie prede. La stessa cosa, ma per altri
motivi, cerca di fare l'uomo, inventando sostanze che bloccano la conduzione di sostanze elettrici, come la
lidocaina, composto di origine sintetica che blocca i canali per il sodio (effetto anestetico locale).
Le terminazioni dell'assone sono prive di guaina mielinica. In queste arriva il potenziale d'azione. Queste
ramificazioni prendono contatto con le strutture bersaglio, che possono essere neuroni o muscoli o ghiandole.
Tra la terminazione dell'assone e la struttura bersaglio c'è molto spesso uno spazio fisico che li separa e
questo comporta un problema, perchè qui la corrente elettrica non può propagarsi. In questo spazio c'è
bisogno di qualcosa che porti il segnale dalle terminazioni dell'assone alle strutture bersaglio. Una volta che
il potenziale d'azione arriva nel terminale, non può tornare indietro a causa del periodo refrattario e non può
andare avanti. C'è quindi bisogno di qualcosa che attraversa lo spazio fisico: il neurotrasmettitore, sostanza
chimica che passa dal terminale alla struttura bersaglio.
6. Recettori ionotropi e metabotropi
Abbiamo due diversi tipi di recettori: ionotropi e metabotropi. Nello stesso recettore possono coesistere. Gli
ionotropi sono sempre formati da proteine che attraversano la membrana cellulare e formano al proprio
interno un canale ionico. Quando si attiva un recettore ionotropo, il canale si apre e fa passare determinati
ioni. Per cui un recettore ionotropo si chiama così proprio perchè fa passare gli ioni, una volta attivato il
canale ionico.
I recettori metamotropi sono anch'essi formati da proteine che attraversano la membrana, ma non contengono
nessun canale per gli ioni, quindi non sono in grado di determinare delle correnti elettriche. Essi sono in
grado, invece, di regolare il metabolismo del neurone.
I recettori ionotropi hanno un canale per il ioni che si apre quando un neurotrasmettitore si lega a questi
recettori. Nel potenziale d'azione, come abbiamo visto, lo stimolo che apriva i canali era la depolarizzazione,
cioè uno stimolo di natura elettrica. Avevamo chiamato quei canali voltaggio-dipendenti. Questi canali non
sono sensibili a stimoli di natura elettrica, ma sono aperti o chiuso da stimoli di natura chimica (canali ionici
a porta chimica o ligando-dipendenti). Il neurotrasmettitore si lega all'esterno del recettore e determina
l'apertura del canale ionico, ma il neurotrasmettitore rimane legato all'esterno del recettore senza entrare nel
neurone. Possiamo suddividere i recettori ionotropi in quattro differenti famiglie in base allo ione che fa
passare. Uno è quello che fa passare il cloro, che causa un'iperpolarizzazione perchè il cloro porta cariche
negativo. Il GABA si lega molto spesso a questi recettori. Nel secondo tipo si apre il canale per il potassio,
che è più concentrato dentro, esce e causa iperpolarizzazione. In un altro tipo di recettore passa sia il sodio
che il potassio, in quanto il canale non è molto specifico: il sodio tende a entrare, il potassio tende ad uscire.
In questo caso, se ad ogni moleca del sodio ne corrisponde una del potassio, non cambiarebbe niente; in
realtà questi recettori sono più specifici per il sodio, percui entrerà una quantità molto maggiore di sodio
rispetto a quanto potassio esca. L'effetto finale è quindi di depolarizzazione. Oltre a passare il sodio e il
potassio, nel quarto tipo di recettore, passa anche il calcio. Il calcio, nell'elemento post-sinaptico, è uno ione
importante per la plasticità sinaptica, cioè per far sì che il neurone si modifichi. Il calcio è fondamentale
anche per tutti i processi di apprendimento e memoria. Entrando molto calcio e molto sodio e uscendo il
potassio, avremo una depolarizzazione. Il segnale, da chimico, è stato nuovamente trasformato in codice
neurale, in segnale di natura elettrica. Nei neuroni la maggior parte degli stimoli è rappresentata dai
neurotrasmettitori.
Parliamo di sinapsi eccitatorie: sono sinapsi in cui in conseguenza dell'apertura dello ionotropo, abbiamo una
depolarizzazione, chiamata eccitatoria perchè può causare potenziale d'azione. Tutte quelle sinapsi la cui
apertura determina una ipepolarizzazione, ci allontana dal potenziale d'azione perchè rende l'interno del
neurone più negativo e ci allontana dalla soglia (sinapsi inibitorie). Il glutammato in generale si lega a
recettori che inducono depolarizzazione. Il GABA in generale si lega a recettori la cua apertura determina
iperpolarizzazione.
Mettiamo tutto insieme ciò che abbiamo visto nelle lezioni precedenti. Nei neuroni, contemporaneamente,
arriva una quantità di segnale enorme, portata dagli assoni degli altri neuroni, formando sinapsi. In rosso le
sinapsi eccitatorie, in blu le sinapsi inibitorie. La quantità di segnali che riceve un singolo neurone è enorme.
Le "orecchie" del neurone sono i dendriti, che contengono recettori, che si attivano quando arrivano dei...
L'informazione si propaga arrivando al corpo cellulare. L'assone è la bocca del neurone, che parla alla
strutture bersaglio. L'assone si ramifisca e forma delle sinapsi. Arrivato qui il potenziale d'azione, viene
liberato il potenziale d'azione.
Arriva il neurotrasmettitore, che si lega ai recettori inostropi, che contengono canali ionici aperti da stimoli
di natura chimica; aperti, fanno passare degli ioni e possiamo avere di potenziali locali (depolarizzazione o
iperpolarizzazione), raggiungono il cono d'emergenza, dove ci sono tanti canali ionici sensibili a stimoli di
natura elettrica. Arrivate qui i potenziali locali, i canali si aprono. I canali ionici voltaggio-dipendenti fanno
passare o solo il sodio o solo il potassio. Nei recettori ionotropi i canali sono meno specifici perchè fanno
passare sodio e potassio attraverso il medesimo canale.
Possiamo avere due diversi tipi di sommazione delle informazioni. La sommazione spaziale: potenziali
spazialmente distanti, se arrivano contemporaneamente si sommano tra loro. Nella sommazione temporale,
due depolarizzazioni si sommano tra di loro se le depolarizzazioni sono consecutive.
Le sinapsi eccitatorie sono a loro volta regolate da altre sinapsi inibitorie (inibizione presinaptica).
Il secondo tipo di recettori sono i metabotropi, anch'essi sensibili ai neurotrasmettitori, formati da proteine
che attraversano la membrana cellulare, ma non hanno nessun canale ionico, quindi non sono in grado di
trasformare lo stimolo da natura chimica a elettrica. Questi hanno una proteina G. Una volta arrivato il
neurotrasmettitore, si lega al suo recettore e in conseguenza di questo legame, la proteina G si attiva e si
stacca fisicamente dal recettore. Ci sono diversi tipi di proteine G con bersagli differenti. La prima cosa che
può fare la proteina G è quella di andare a regolare l'attività di un canale ionico vicino, regolandola. Altre
proteine G possono andare ad attivare altre proteine, chiamati secondi messaggeri, che vanno ad attivare una
serie di reazioni a cascata che portano all'attivazione di altre proteine che regolano i geni e quindi l'intero
metabolismo del neurone. Da qui il loro nome 'metabotropo'. In questo processo entra il fenomeno
dell'amplificazione del segnale: da un singolo neurotrasmettitore, viene attivato il recettore, che attiva la
proteina G; questa stimola l'adinilato ciclasi per convertire l'ATP in AMPc.
Le sinapsi di natura chimica sono estramemente e finemente regolabili, ecco perchè la maggior parte delle
sinapsi sono di natura chimica. Un'altra particolarità è che sono plastiche, cioè si modificano a seconda della
attività; questo processo sta alla base dell'apprendimento e della memoria.
7. Anatomia del sistema nervoso
Possiamo suddividere il cervello in tre diversi piani perpendicolari tra di
loro: orizzontale, frontale e sagittale. Più importanti sono però le
coordinate anatomiche.
Nei
mammiferi
a
quattro zampe abbiamo
davanti la porzione
anteriore,
dietro
la
porzione posteriore, sul
dorso c'è la porzione
dorsale e sul ventre c'è
la posizione ventrale. Tutto ciò che sta dentro viene
chiamato mediale, ciò che sta ai lati viene chiamato
laterale. La stessa cosa avviene nell'uomo.
Il sistema nervoso è formato da due diversi tipi di sostanze: sostanza bianca e sostanza grigia. Abbiamo visto
che ci sono assoni ricoperti da mielina, che al microscopio ci appare bianca. La sostanza bianca è quindi
formata da assoni ricoperti da mielina (è suddivisa in nervi, tratti, fasci). Tutto quello che non è bianco fa
parte della sostanza grigia. In questa ci saranno dendriti, corpi cellulari, assoni non ricoperti da guaina
mielinica. La sostanza grigia è suddivisa in corteccia e nuclei. Parliamo di corteccia quando al microscopio
vediamo i neuroni disposti su file ben precise. La corteccia più antica evolutivamente aveva un numero di
strati limitato, due o tre: parliamo di archicorteccia o paleocorteccia. La corteccia più recente è formata da un
numero maggiore (precisamente 6) di strati (neocorteccia). Parliamo di nuclei o gangli quando i neuroni non
sono disposti in maniera ordinata. Nuclei e corteccia sono collegati tra di loro attraverso gli assoni, molto
spesso ricoperti da mielina.
Possiamo dividere il sistema nervoso in due parti: sistema nervoso centrale e sistema nervoso periferico. Il
periferico è formato da tutti quegli assoni che si ramificano e prendono contatto con muscoli e organi interni.
Il sistema nervoso centrale viene suddiviso in midollo spinale ed encefalo. Nel sistema nervoso periferico
abbiamo il sistema nervoso autonomo, che regola il nostro ambiente interno, come il battito cardiaco o la
respirazione, autonomo in quanto indipendente dalla nostra volontà. Oltre a questo abbiamo una parte
dettasomatica, formata da una parte sensoriale e una motoria. Nel sistema nervoso centrale nell'encefalo
abbiamo innanzitutto il midollo allungato o bulbo, che è una continuazione del midollo spinale, il ponte, il
mesencefalo, il diencefalo, il telencefalo e il
cervelletto. Il sistema nervoso è collegato in
maniera bidirezionale sia col nostro ambiente
interno che con quello esterno: il sistema
nervoso riceve e invia informazioni. Tutte le
strutture che portano le informazioni dalla
periferia al sistema nervoso centrale, sono
chiamate afferenti. Viceversa, sono chiamate
efferenti.
Il midollo spinale lo troviamo presente lungo
tutta la colonna vertebrale e continua col midollo
allungato o bulbo, seguito dal ponte e dal
mesencefalo. Ponte, midollo allungato e
mesencefalo formano il tronco encefalico. Sopra
il cervelletto, poi il diencefalo e infine tutto il
resto viene chiamato telencefalo. Durante lo sviluppo a livello embriale, tutto prende origine da un vero e
proprio tubo che presenta una cavità al proprio interno e rivestito da una serie di neuroni che si dividono:
tubo neurale. In questo tubo è possibile identificare diverse regioni, da cui prenderanno origine le diverse
strutture che abbiamo visto. Queste regioni, con lo sviluppo, daranno origine alle diverse parti del sistema
nervoso. La cavità ripiena di liquido all'interno del tubo viene chiamato liquido cerebrospinale. Nella prima
fase, chiamata a 3 vescicole, abbiamo solo tre segmenti. Da qui i neuroni si dividono e proliferano, verso la
fase a 5 vescicole, dando vita alle diverse parti. Anche la cavità si accresce formando delle cavità presenti nel
nostro sistema nervoso centrale di adulto. Il cervello non è una struttura unica e compatta, ma vi sono delle
cavità presenti in tutto l'encefalo. Nel midollo spinale c'è il canale centrale, che si allarga a formare i
ventricoli (cavità ripiene di liquido cerebrospinale). A livello del tronco encefalico abbiamo il quarto
ventricolo, dopodichè abbiamo l'acquedotto cerebrale (chiamato di Silvio), che continua col terzo ventricolo.
Questo si suddivide a formare i ventricoli laterali destro e sinistro. Una prima funzione dei ventricoli è di
tipo metabolico. La funzione fondamentale è però quella meccanica: la porzione esterna non va a schiacciare
il tronco encefalico grazie ai ventricoli, che sono strutture elastiche.
Il midollo spinale si trova nella colonna vertebrale e viene suddiviso in diverse parti:
abbiamo la porzione cervicale, toracica, lombare e sacrale. Il midollo spinale è
suddiviso in veri e propri segmenti a livello delle vertebre. I segmenti sono anche
chiamati metameri e questa suddivisione viene chiamata metamerica. Il midollo non
arriva sino al fondo della colonna vertebrale, ma si arresta a livello lombare-sacrale.
Da lì usciranno una serie di assoni che andranno a innervare le altre parti del nostro
corpo. Questo fascio di assoni forma la cauda equina.
In un segmento ci saranno assoni che portano informazioni di natura sensoriala;
parliamo di dermatomero. Nello stesso segmento partiranno informazioni per i
muscoli: parliamo di miomero. L'insieme della componente sensoriale e della
componente motoria è chiamato mielomero. Questo avviene perchè nel nostro
midollo abbiamo sia una componente motoria che sensoriale.
Il midollo spinale è formato da una sostanza bianca ed una sostanza grigia. Nel
midollo spinale la sostanza grigia si trova nella parte più interna, mentre la sostanza
bianca si ritrova nella parte esterna. Il puntino al centro è il canale cerebrale dove si
trova il liquido. La sostanza bianca viene suddivisa in colonne, chiamate dorsali se
si trovano a livello del dorso o ventrali se si trovano a livello del ventre. Di lato
abbiamo la colonna laterale. La sostanza grigia viene suddivisa in corna e anche qui
parleremo di corna dorsali e ventrali.
Il midollo spinale è funzionalmente diviso in porzione ventrale e dorsale. A livello
dorsale, nel midollo spinale, arrivano le informazioni di natura sensoriale. A livello ventrale escono le
informazioni di natura motoria. Partendo dalla periferia, stimolando il tatto, prendono origine dal polpastrello
degli assoni che originano informazioni a livello dorsale. Tutte le informazioni sensoriali vengono portate al
midollo spinale, ma il corpo cellulare degli assoni si trova fuori. Il corpo cellulare non è ricoperto da mielina
e formerà sostanza grigia: parliamo di gangli. I neuroni hanno perciò il corpo cellulare nei gangli. Le
informazioni entrano dunque nel midollo spinale attraverso la cosiddetta radice. Da qui, una parte si ramifica
e una parte si porta a livello ventrale. A livello ventrale abbiamo il corpo cellulare di tutti quei neuroni che
inviano il proprio assone fuori dal midollo spinale e raggiungerà i muscoli. Questi neuroni sono chiamati
motoneuroni. L'assone, uscendo, si ricopre di mielina (avremo dunque la radice ventrale) e si avvicinerà
fisicamente al neurone sensoriale, formando il nervo spinale. Si forma una sinapsi neuromuscolare, che
funziona esattamente come tutte le altre sinapsi di natura chimica e il neurotrasmettitore è l'acetilcolina.
Oltre ad avere questo circuito, abbiamo che il midollo spinale è in contatto diretto col tronco encefalico ed
telencefalo. Dal midollo spinale ci sono assoni che salgono e portano informazioni sensoriali al tronco
encefalico e telencefalo (vie sensoriali ascendenti/afferenti) e altri che scendono e portano informazioni
motorie dal tronco encefalico e telencefalo (vie motorie discendenti/efferenti).
Il tronco encefalico è formato da tre strutture: midollo
allungato o bulbo, ponte e mesencefalo. Il tronco encefalico,
visto al microscopio, è molto più complesso del midollo
spinale, perché abbiamo tutte queste vie di sostanza bianca
discendenti e ascendenti e, immerse, abbiamo delle vere e
proprie isole di sostanza grigia. Come cavità abbiamo il quarto
ventricolo. Nel tronco encefalico abbiamo dei neuroni che
escono per andare a regolare funzioni vitali come il battito
cardiaco. Oltre a questo, ci sono anche neuroni che portano
informazioni di natura sensoriale e altri che vanno a regolare i
nostri movimenti. Il tronco encefalico è fondamentale per la
sopravvivenza dell'individuo, tanto che è il punto più cruciale del sistema nervoso. Anche dal tronco
encefalico entrano ed escono nervi, in particolare 12. Una caratteristica del trocno encefalico è che qui
possiamo avere nervi o solo sensoriali o solo motori o misti, mentre nel midollo spinale osno tutti misti.
Sotto il tronco encefalico, abbiamo il cervelletto, importante per il movimento e l'apprendimento motorio. È
separato dal tronco encefalico fisicamente dal IV ventricolo. Il cervelletto è collegato al tronco grazie ai tre
peduncoli cervellari. Quello superiore è collegato al mesencefalo, il medio è collegato al ponte e quello
inferiore al bulbo.
Il diencefalo presenta come cavità il terzo ventricolo. Nel diencefalo abbiamo due strutture fondamentali:
talamo e ipotalamo. L'ipotalamo è il punto dove il nostro cervello prende contatto con il sistema endocrino.
L'ipotalamo è in contatto fisico diretto con una ghiandola importante: ipofisi. Tanto che si parla di asse
ipotalamoiposale. Nell'ipofisi c'è una parte in cui arrivano i neuroni e una parte formata da. L'ipotalamo
svogle anche altri compiti importanti come regolare il sistema nervoso autonomo, i cicli sonno-veglia, il
senso di sazietà e fame. Il talamo, invece, svolge una funzione totalmente differente. Il talamo non è un unica
struttura, ma un insieme di tanti nuclei. Il talamo è la stazione a cui arrivano la maggior parte delle info
sensoriali; dalla periferia le informazioni tattili, visive, uditive arrivano direttamente al talamo. Dal talamo le
informazioni sono ritrasmesse alle corteccie, le quali si servono per regolare la visione, etc. Talamo e
ipotalamo sono vicine anatomicamente ma svolgono funzioni diverse. Entrambi sono formati da sostanza
grigia. Le informazioni al talamo arrivano dalla periferie; si tratta di vie principalmente formati da guaina
mielinica (sistanza bianca), chiamata capsula interna. Il talamo è nella profondità del nostro encefalo. Sotto il
talamo, a livello delle pareti del terzo ventricolo, abbiamo l'ipotalamo.
Il telencefalo occupa la maggior parte dell'encefalo. È formato da corteccia cerebrale, dal corpo calloso, dai
gangli della base, dai nuclei del setto, dall'amigdala. All'opposto del midollo spinale, la sostanza grigia si
trova principalmente all'esterno. Questa sostanza grigia forma la corteccia e in particolare vedremo essere
principalmente neocorteccia. Immersi come delle isole nella sostanza bianca, abbiamo dei nuclei. Abbiamo i
nuclei della base (o gangli della base), nuclei del setto e l'amigdala. Il telencefalo viene suddiviso in due
emisfero, di destra e di sinistra. Questi due emisferi sono separati fisicamente dalla scissura interemisferica. I
due emisferi comunicano tra di loro attraverso un fascio di sostanza bianca chiamato corpo calloso. Un'altra
caratteristica è che all'esterno vi è neocorteccia: questa nell'uomo presenta solchi e circonvoluzioni. Nei
mammiferi però non è sempre così, perchè la corteccia è presente ma liscia. Inizia a presentarli nel gatto fino
a raggiungere il massimo nell'uomo. Questo avviene perchè nei primati e nell'uomo aumenta enormemente il
volume di neocorteccia presente e per farla stare dentro il crano, dev'essere ripiegata su se stessa. Nella
neocorteccia identifichiamo quattro lobi: un lobo frontale, che si trova davanti, un lobo temporale che si
trova di lato, a livello delle tempie, un lobo parietale, sopra, e infine, dietro, un lobo occipitale. Questi lobi
sono anatomicamente separati tra di loro da scissure, ad esempio il lobo frontale è separato dal parietale dalla
scissura di Rolando. Un altro solco, scissura di Silvio, separa il lobo temporale da quello frontale e parietale.
La maggior parte del nostro telencefalo è formato da sostanza grigia che è neocorteccia nei primati e
nell'uomo. Ci sono però, come abbiamo detto, altri tipi di corteccia, formate da meno di 6 strati. L'ippocampo
è suddiviso in tre diversi strati ed è archicorteccia. Oltre all'ippocampo un'altra archicorteccia è quella
olfattiva. Evolutivamente, nei vertebrati fino all'uomo, ciò che succede nei mammiferi è un auemnto enorme
del telencefalo rispetto alle altre parti del cervello. Quello che si sviluppa di più nel telencefalo dei
mammiferi è la neocorteccia.
La neocorteccia è suddivisa in sei strati. Abbiamo due diversi
tipi di neuroni: i cosiddetti a proiezione, cioè quei neuroni il cui
assone tende ad uscire dalla neocorteccia. Questi hanno il corpo
cellulare che richiama la forma triangolare a piramide, per
questo questi neuroni sono anche chiamati piramidali. Il
secondo tipo è dato da neuroni che non escono dalla corteccia e
vengono chiamati interneuroni. La forma di questi neuroni
richiama quella di una stella e per questo vengono chiamati
cellule stellate.
Le informazioni, dalla periferia, arrivano principalmente al IV
strato della corteccia. Qui si trovano tanti interneuroni che
ricevono queste informazioni (IV strato) e inviano il loro assone
agli strati I, II e III. In questi ci sono anche dei dendriti di
neuroni il cui corpo neurale si trova nel V e VI strato, quindi a
proiezioni, inviandole ad altre strutture sottocorticali. Nel I, II e
III strato ci sono anche dei neuroni che inviano il proprio
assone fuori (a proiezione) che però raggiunge altre corteccie,
connettendo le corteccie tra loro.
Ciascuno di questi strati presenta una spessore diverso. Ci sono corteccie che presentano un IV strato molto
sviluppato, mentre un V poco sviluppato. Per altre corteccie è l'opposto. In altre IV e V sono poco sviluppati
rispetto agli altri. Brodmann ha misurato e mappato le aree a seconda dello spessore di questi strati. Questo
tipo di classificazione non è funzionale, ma in base allo spessore degli strati. Le diverse corteccie sono
diverse di spessore ma anche di funzione. Ci sono corteccie che svolgono funzioni princiapalmente sensoriali
(visive, uditive), altri motorie. Determinate strutture del nsotro cervello svolgono u ndetermiato compito, ma
non possiamo localizzare funzioni superiori ad un determinata parte del nsotro cervello, come faceva la
frenologia.
La corteccia visiva elabora stimoli visivi, quindi uno degli strati più sviluppati in questa sarà il IV strato,
perché riceve le informazioni. Quindi nelle cortecce sensoriali lo strato particolarmente sviluppato è il IV,
perchè è lo strato a cui arrivano le informazioni dal talamo. Invece, V e VI strato non sono molto sviluppati,
perchè non è importante inviare le informazioni alle strutture sottocorticali. Nelle corteccie motorie non è
importante ricevere ed elaborare stimoli, quindi il IV strato non sarà molto sviluppato, invece saranno più
sviluppati questi strati (VI e V) che inviano le informazioni alle strutture sottocorticali. Soprattutto nell'uomo
ci sono cortecce che non rispondono solo a stimoli visivi e non sono neanche motorie, sono le cortecce
associative: cortecce nè strettamente sensoriali nè strettamente motorie, ma associano e integrano tra loro
tutte le informazioni sensoriali e motorie. Il IV e V strato non è molto sviluppato, ma sono molto sviluppati
quegli strati che servono a integrare le informazinoi, cioè I, II e III.
Abbiamo tanti diversi tipi di cortecce motorie e
sensoriali. Le cortecce sensoriali primarie o di primo
ordine sono quelle che elaborano gli aspetti più
semplici ed elementari. Altre svolgono funzioni più
complesse e sofisticate, chiamate cortecce sensoriali
di ordine superiore. Questo avviene per le cortecce
uditive, visive, etc. La stessa cosa avviene nelle
cortecce motorie: abbiamo una corteccia motoria
primaria, che regola l'esecuzione del movimento, e
delle cortecce motorie di ordine superiore che servono
per pianificare quale azione fare, come agire, etc.
Evolutivamente, nell'uomo succede che aumentano
enormemente le cortecce di ordine superiore e le
cortecce associative. La corteccia associativa limbica
prende parte all'associazione degli stati emotivi. La
corteccia associativa parieto-temporo-occipitale le cui
funzioni sono oggi poco note. Tutte queste cortecce
sono collegate tra di loro grazie al corpo calloso.
8. Circolo ematico cerebrale
Possiamo distinguere un circolo antieriore e un circolo
posteriore. Le arterie carotidi e vertebrali danno vita a un
circuito complesso chiamato circolo di Willis. Questo circuito
ha la forma di un poligono e avendo questa forma consente che
semmai succedesse un danno a un'arteria cartotine o vertrebrale
il sangue portato dagli altri vasi riesce lo stesso ad arrivare
all'encefalo. Partendo dal cuore, attraverso le arterie pulsante
porta il sangue al circolo di Willis, da qui i vasi diventano
sempre più piccoli (capillari) che si distribuiscono su tutta la
massa del sistema nervoso centrale. Il sangue, a livello dei
capillari, porta sostanze nutrienti che vengono scambiati con i
neuroni e ritorna come sangue venoso al cuore per essere
inviato ai polmoni.
Tutto l'encefalo e il midollo è ricoperto da membrane chiamate
meningi. Abbiamo tre meningi: la dura madre, più esterna,
attaccata all'osso del cranio; l'aracnoide, formata da strutture
trammentose che formano una sorta di ragnatela. Lo spazio
subaracnoide ci sono i capillari e il liquido cerebrospinale.
Infine la meninge più attaccata all'encefalo è la pia madre e separa l'encefalo dai capillari. Dai capillari e la
pia madre abbiamo uno spazio chiamato perivascolare, che evita che le pulsazioni arteriose siano trasmesse
al tessuto nervoso.
Il liquido cerebrospinale viene prodotto nei ventricoli da cellule epiteliali che formano i plessi corioidei. Una
volta prodotto dai ventricoli, passerà dallo spazio subaracnoideo e verrà poi assorbito dal circolo venoso.
Dal cuore il sangue attraversa le arterie, raggiunge il circolo e l'interno del nostro encefalo,
contemporaneamente il sangue raggiunge i plessi corioidei, dove abbiamo . Dopodichè i lsangue verrà
assorbito dal circolo venoso e così anche il liquido cerebrospinale. Ecco come avviene in maniera schematica
la circolazione nell'encefalo.
Esempio di compito d'esame (vero o falso):
- Nei mammiferi tutti gli assoni sono ricoperti da guaina mielinica.
- La guiana mielinica determina riduzione di resistenza interna cito.
- I nervi spinali fanno parte del SNC.
- I ventiricoli cerebrali sono cavità contenenti liquido cerebrospinale.
- Le meningi sono strutture membranose che avvolgono l'encefalo ma non il midollo spinale.
- Il telencefalo comprende corteccia cerebrale, corpo calloso e alcuni nuclei sottocorticali come i gangli della
base.
- Durante la fase discendente del potenziale, i canali epr il sodio voltaggio-dipendenti sono inattivati.
- Col crescere dell'intensità degli stimoli, aumenta l'ampiezza dei potenziali locali.
- Le sinapsi elettriche sono più rapide delle chimiche.
9. Omeostasi
Il sistema nervoso autonomo partecipa alla regolazione del nostro ambiente interno, cioè quella che viene
definita omeostasi. Il sistema nervoso è in cotnatto diretto sia con l'ambiente esterno che con quello interno.
All'interno del corpo ci sono tanti parametri di valori che devono essere mantenuti il più costante possibili,
finchè il nostro organismo funzioni bene. Questi valori sono, per esempio, la temperatura corporea, la
concentrazione di glucosio nel sangue, la quantità di ossigeno nel sangue, la quantità di anidride carbonica
disciolta nel sangue, la pressione arteriosa. Questi parametri sono chiamati omeostatici. Di questo compito se
ne occupa il sistema nervoso, che, attraverso i suoi contatti con il sistema muscolare e endocrino, cerca di
mantenere costanti questi parametri. Questo avviene attraverso la regolazione a feedback negativo: abbiamo
un determinato parametro che viene costantemente monitorato da determinati sensori. Questi sensori inviano
le informazioni a una struttura chiamata effettore. L'effettore, rilevando la modificazione del parametro,
metterà in modo una serie di reazioni che hanno come scopo quello di riportare il parametro ai suoi valori
originali.
Tutte le informazioni vengono inviate al sistema nervoso centrale e da qui raggiungono due strutture chiave
per la regolazione dell'omeostasi: l'ipotalamo e il nucleo del tratto solitario, in cui si contronta il valore
originario con le eventuali modifiche di questo. L'ipotalamo paragona il valore ricevuto con quello settato e
se rileva qualche modificazione, si attiva per riportare il parametro ai suoi valori originali. L'ipotalamo può
attivare diversi sistemi per ripristinare il valore: il primo è il sistema nervoso autonomo e il secondo è
l'ipofisi, che regola le ghiandole endocrine. L'ultimo meccanismo attraverso il quale l'ipotalamo può
ripristinare il valore sono i comportamenti motivati. Esempio: pressione arteriosa. Supponiamo che aumenta;
l'aumento viene rilevato dai sensori, recettori sensibili alla variazione della pressione e chiamati barrocettori.
Questi, che si trovano nei vasi, inviano l'informazione al nucleo del tratto solitario e all'ipotalamo. Questi
ultimi aittivano il sistema nervoso autonomo e verrà diminuita la gittata cardiaca e la pressione arteriosa
diminuisce. Un altro esempio è la temperatura, settata intorno ai 36,5 °C. Il valore diminuisce, i termocettori
informano l'ipotalamo, che si attiva, rileva che la temperatura si è modificata e va ad attivare il sistema
nervoso autonomo per far sì che i vasi che portano il sangue ai distretti perifici tendono a restringersi così
che arrivi meno sangue in periferia; un'altra cosa che farà sarà di attivare il sistema endocrino per attivare la
tiroide; comportamenti motivati, cioè quello di indossare un maglione. Grazie all'attivazioen di questi tre
sistemi il nostro organismo cerca di riportare il parametro omeostatico al valore originario.
Il sistema nervoso autonomo deve questo nome al fatto di essere indipendente dalla nostra volontà, ma
nonostante ciò è fortemente influenzato dalle esperienze dell'individuo. È formato da una rete di neuroni che
si distribuisce in tutto l'organismo, raggiungendo tutti gli organi interni e la cute. I bersagli principali saranno
tutte le ghiandole e i muscoli involontari, la muscolatura liscia. Nel sistema nervoso autonomo possiamo
distinguere due diverse componenti: SNA ortosimpatico (o simpatico) e SNA parasimpatico. In linea
generale, queste due componenti hanno effetti opposti. Il SNA parasimpatico serve per mantenere costanti i
parametri omeostatici e funziona quando siamo in una situazione di riposo. Il SNA ortosimpatico serve per
spostare e modificare rapidamente i parametri omeostatici; in determinate situazioni, come di pericolo o di
stress, questo interviene modificando i valori.
Esempio di attivazione del SNA ortosimpatico, definita reazione da stress. Quando siamo in una situazione di
pericolo devono funzionare particolarmente bene due strutture: cervello (per elaborare gli stimoli
rapidamente) e i muscoli (per reagire al pericolo). Per farli funzionare bene sono necessari due elementi:
zucchero (glucosio) e ossigeno. Il glucosio è immagazzinato nel fegato; le riserve di glucosio più facilmente
disponibili sono lì. Viene immagazzinato legando tante molecole di glucosio formando il glicogeno. Il
sistema nervoso autonomo ortosimpatico attiverà il fegato affinchè venga rilasciato glucosio nel sangue.
L'ossigeno viene preso attraverso i polmoni e viene attivata la frequenza del respiro dal SNA ortosimpatico.
A questo punto c'è bisogno di mandare maggiore quantità possibile di sangue a cervello e muscoli, il SNA
aumenta dunque frequenza e gittata cardiaca. Questo non basta perchè il sangue tenderebbe a distribuirsi
uniformenente in tutto l'organismo, mentre in una situazione di pericolo è necessario che venga indirizzato al
cervello e i muscoli. Il SNA ortosimapatico fa sì che i vasi si restringono cosicchè il sangue non si dispera a
distretti in cui non è necessario che il sangue arrivi troppo. Contemporaneamente, se da una parte questi vasi
si restringono, dall'altra si dilateranno per portare il sangue a cervello, muscoli e cuore (in quanto
quest'ultimo deve lavorare in maniera particolarmente intensa).
Il sistema orto e parasimpatico sono fatti dai neuroni che si trovano, nel primo caso (ortosimpatico) tutti nel
midollo spinale. Nel caso del parasimpatico, i neuroni che lo formano si trovano in parte nel midollo spinale
e in parte nel tronco encefalico. Nel sistema nervoso ortosimpatico e parasimpatico possiamo identificare due
diversi tipi di neuroni. Il SNA è formato da un neurone che poi va a prendere contatto con un altro neurone:
ci sono quindi due neuroni. Il primo invia il proprio assone a prendere contatto con il secondo; il secondo, a
sua volta, invia l'assone verso le strutture bersaglio. Il corpo cellulare del primo neurone si trova o nel
midollo spinale o nel tronco encefalico. Il secondo neurone ha il corpo cellulare fuori dal midollo spinale o
nel tronco encefalico, nel ganglio del SNA. Il primo neurone verrà quindi chiamato pregangliare e il secondo
postgangliare.
I neuroni pregangliali del SNA orto e parasimpatico liberano il medesimo neurotrasmettitore, l'acetilcolina. I
neuroni postgangliali del SNA parasimpatico continuano a usare l'acetilcolina. Nel caso del SNA
ortosimpatico i neuroni postgangliali usano la noradrenalina. Questo perché se entrambi usassero lo stesso
neurotrasmettitore avremmo esattamente lo stesso effetto sull'organo bersaglio, percui è necessario
differenziare. Abbiamo anche altre differenze importanti, tra cui la lunghezza del neurone pre e
postgangliale. L'assone del neurone postgangliare del SNA parasimpatico è particolarmente lungo, quello del
parasimpatico è corto. I gangli del SNA parasimpatico si trovano più vicini all'organo effetto, mentre quelli
del SNA ortosimpatico sono più vicini al midollo spinale, dove ci sono i neuroni pregangliali. Questo perché
abbiamo una terza differenza tra i due sistemi: l'ortosimpatico, tramite un assone pregangiale, raggiunge
contemporaneamente un numero elevato di neuroni postgangiali. Il sistema parasimpatico innerva un numero
molto più basso di gangli e quindi di organi bersaglio. Questo avviene perchè l'ortosimpatico è un sistema
d'emergenza quindi è molto più vantaggioso che attivi tanti neuroni contemporaneamente in una situazione
di pericolo. Questo spiega le differenze precedenti. Altra differenza: tutti i neuroni pregangliali del sistema
nervoso ortosimpatico hanno il corpo cellulare nella sezione lombare e toracica del midollo spinale. Nel
sistema parasimpatico una parte dei neuroni si trova a livello sacrale e un'altra parte nel tronco encefalico.
L'ipotalamo regola l'omeostasi attraverso il SNA e il sistema
endocrino. L'ipotalamo si trova nel diencefalo ed è formato da
diverse popolazioni di neuroni, alcuni dei quali servono per
regolare il SNA, altri per regolare i comportamenti motivati e
altri per regolare le ghiandole attraverso l'ipofisi. L'ipotalamo
regola le nostre ghiandole attraverso l'ipofisi. L'ipofisi viene
distinta in due diverse parti: ipofisi posteriore o neuroipofisi e
ipofisi anteriore o adenoipofisi. Queste due parti sono molto
diverse tra di loro perchè nella neuroipofisi arrivano gli assoni
dei neuroni ipotalamici, la quale è quindi formata da tessuto
nervoso. Nell'adenoipofisi non abbiamo più neuroni, ma
tassuto ghiandolare. L'ipotalamo è in grado di regolare
entrambe queste parti. Vediamo cosa succede a livello della
neuroipofisi: qui arrivano gli assoni dei neuroni ipotalamici,
formando il tessuto nervoso della neuroipofisici, ramificandosi
e raggiungendo i capillari. Questi neuroni sono chiamati
magnocellulari perché sono molto grandi. Questi neuroni
ipotalamici vanno a produrre determinate sostanze, liberate nei
capillari a livello della neuroipofisi. Liberandole nel sangue,
queste sostanze avranno effetto su organi lontani dalla
neuroipofisi. Queste due sostanze sono l'ossitocina e la
vasopressina. L'ossitocina ha come bersaglio principale, nelle donne, l'utero, in quanto è importante durante
il parto, perchè consente la contrazione della muscolatura liscia dell'utero e quindi l'espulsione del feto. Un
altro bersaglio sono le ghiandole mammarie perchè consente l'espulsione del latte dopo il parto. La
vasopressina invece svolge compiti diversi, chiamato ormone antidiuretico, perchè ha come bersaglio i reni.
Queste due sostanze sono classificate come neurotrasmettitori; altre sostenze possono essere classificate
come ormoni, sostanze che vengono liberate nel sangue e agiscono a distanza. I neurotrasmettitori invece
hanno meccanismi d'azione differenti: sono prodotti localmente, non sono liberati nel sangue e agiscono su
organi bersaglio vicini. Ossitocina e vasopressina sono prodotti da neuroni ma agiscono come ormoni perchè
vengono liberate nel sangue e agiscono a distanza: vengono perciò chiamati neurormoni.
A livello dell'adenoipofisi, formata da tessuto ghiandolare (e non più nervoso), abbiamo vere e proprie
ghiandole che vanno a liberare nel sangue degli ormoni. Nell'adenoipofisi sono prodotti e liberati nel sangue
sei diversi tipi di ormoni. Nell'ipotalamo abbiamo altri neuroni che inviano il proprio assone a un circolo
sanguigno dei vasi che si trovano nell'adenoipofisi, che liberano dei fattori che ragigungono il tessuro
ghiandolare dell'adenoipfisi e servono per stimolare o inibire la produzione degli ormoni a livello
dell'adenoipofisi. Uno di questi ormoni è la prolattina. I neuroni che liberano i fattori hanno corpo cellulare
più piccolo e per questo vengono chiamati parvocellulari.