12
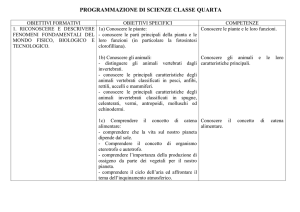
RICONSIDERAZIONI
SULL’EVOLUZIONE
DEI VERTEBRATI
SOMMARIO
12.1
Omeostasi
12.2
Cresta neurale: una novità che ha
permesso l’evoluzione dei
Vertebrati
12.3
Sguardo d’insieme sull’evoluzione
dei Vertebrati
12.1 OMEOSTASI
i
Un concetto fondamentale che si deve tenere in massimo conto per comprendere le differenze anatomiche nei diversi gruppi di Vertebrati e, di conseguenza, la loro filogenesi, è
quello dell’interazione organismo-ambiente. Gli animali devono adattarsi all’ambiente in
cui vivono, ma il termine “adattarsi” suona spesso generico e merita una più compiuta
esplicitazione con esempi adeguati. In molti animali vi è la tendenza a mantenere costanti
alcune caratteristiche fisiologiche. La capacità di mantenere costanti queste caratteristiche
è detta omeostasi. Un semplice esempio di omeostasi è la capacità di mantenere costante,
entro certi limiti, la concentrazione di glucosio nel sangue (glicemia) attraverso l’azione di
due ormoni antagonisti: insulina (ipoglicemizzante) e glucagone (iperglicemizzante). Talvolta, però, si devono affrontare necessità omeostatiche più complesse; inoltre, un tipo di
omeostasi può interagire con un altro in un gioco sempre più complesso.
12.1.1 OMEOSTASI IDRICO-SALINA
Da un certo punto di vista un animale può essere considerato una soluzione acquosa di sostanze organiche ed inorganiche contenuta all’interno di una membrana costituita da epiteli (epidermide, epitelio delle mucose) che la separa dall’ambiente esterno. Questa membrana non è completamente impermeabile e, quindi, permette che avvengano di continuo
scambi con l’ambiente esterno essenziali per la sopravvivenza di un organismo. L’importante è che la concentrazione e la composizione dei liquidi interni dell’organismo non subiscano variazioni e ciò è assicurato dagli organi preposti all’omeostasi idrico-salina. La concentrazione salina e, quindi, l’osmolalità del sangue e dei liquidi interni della maggior parte
dei Vertebrati è compresa tra 250 e 350 mOsm; fanno eccezione i Missinoidi, i Celacanti ed
i Condroitti marini con osmolarità leggermente superiore alle 1000 mOsm. L’osmolarità
dell’acqua di mare è di circa 1000 mOsm e quella dell’acqua dolce intorno a 10 mOsm. È del
tutto evidente che tra i fluidi interni dei Vertebrati acquatici e l’ambiente in cui vivono vi è
un gradiente osmotico che potrebbe portare a disidratazione in molte Specie marine o ad
un eccessivo carico idrico nelle Specie di acqua dolce. Inoltre, la maggior parte dei pesci è
stenoalina, cioè non è in grado di tollerare variazioni della salinità dell’ambiente in cui vive.
VERTEBRATI DI ACQUA DOLCE – Come sappiamo, la superficie corporea dei pesci è poco
permeabile ad acqua e ioni. Vi è, tuttavia, un’area molto delicata in cui gli scambi osmotici
con l’ambiente esterno sono inevitabili: l’epitelio branchiale. Per permettere la respirazione, grandi quantità di acqua devono scorrere sopra l’epitelio branchiale; è proprio a questo livello che l’acqua può entrare per osmosi e gli ioni possono uscire per diffusione. I
fluidi interni tenderebbero a diluirsi in maniera eccessiva se il rene non ristabilisse l’omeostasi. Per compensare l’entrata dell’acqua il rene produce un volume cospicuo di urina ipotonica; i grandi glomeruli dei nefroni permettono un’elevata produzione di ultrafiltrato,
che, però, è isosmotico rispetto al sangue ed ai fluidi interni. Per recuperare i sali, gli ioni
vengono riassorbiti a livello dei tubuli contorti prossimale e distale. Quest’ultimo è impermeabile all’acqua (tratto diluente), di conseguenza l’urina diviene sempre meno concentrata fino a essere iposmotica rispetto ai fluidi interni. Poiché, comunque, alcuni sali vengono persi con l’urina, il pesce dulcacquicolo assume sali attraverso gli alimenti (ecco il
ruolo dell’epitelio intestinale nell’omeostasi) ed, inoltre, speciali cellule localizzate sull’epi342
CAPITOLO 12
Riconsiderazioni sull’evoluzione dei Vertebrati
dermide e sulle branchie, gli ionociti, assorbono contro gradiente ioni Na+ e Cl- dall’acqua
con un notevole costo energetico.
VERTEBRATI MARINI – I gradienti osmotici cui sono sottoposti i Vertebrati marini sono
l’opposto di quelli visti per i Vertebrati dulcacquicoli. Anche nei Teleostei marini, data
l’impermeabilità della cute, la gran parte dei movimenti osmotici si realizza a livello branchiale con notevole perdita di acqua. Per risparmiare acqua, i glomeruli di questi pesci sono
piccoli o addirittura assenti (di conseguenza, l’ultrafiltrazione è minima), così come il tubulo contorto distale è corto o assente: l’urina prodotta sarà quindi poca e molto concentrata. Per compensare la copiosa perdita di acqua a livello branchiale, i Teleostei marini
sono costretti a bere acqua di mare, ma così facendo introducono molto NaCl. Per compensare questo sovraccarico di sali gli ionociti disseminati sulle branchie e nell’epidermide
pompano attivamente ioni Na+ e Cl- verso l’esterno contro gradiente di concentrazione.
Condroitti e Celacanti, viceversa, rendono minimo il gradiente osmotico mantenendo la
concentrazione osmotica dei fluidi interni leggermente superiore a quella dell’acqua di
mare: essi raggiungono questo scopo trattenendo urea. In questo modo i Condroitti possono assumere acqua a livello branchiale per osmosi, non sono costretti a bere, possono
permettersi un nefrone con glomerulo grande e tubulo contorto distale. Per eliminare l’eccesso di NaCl, che comunque si verifica, anche per via alimentare, sviluppano una ghiandola del sale, un’estroflessione dorsale della cloaca, che secerne un fluido isosmotico rispetto ai fluidi corporei, ma con concentrazioni più elevate di ioni Na+ e Cl-.
L’omeostasi idrico-salina, dunque, si basa sulla regolazione dei flussi di acqua e sali minerali in entrata e in uscita attraverso l’epidermide, l’epitelio branchiale, l’epitelio intestinale ed il rene.
OMEOSTASI IDRICO-SALINA E CATABOLITI AZOTATI – Non va, però, dimenticato che il rene è
l’organo più importante per l’eliminazione dei cataboliti del metabolismo azotato, cioè delle
proteine e degli acidi nucleici. L’omeostasi idrico-salina è strettamente legata alla forma con
cui i diversi Vertebrati eliminano i cataboliti azotati. Quando le sostanze azotate vengono
metabolizzate, l’azoto viene ridotto enzimaticamente a NH3 , sostanza altamente tossica ma
molto solubile in acqua. La maggior parte dei pesci e gli Anfibi larvali eliminano NH3 tal quale
(animali ammoniotelici). Condroitti, Celacanti, Anfibi adulti e Mammiferi, viceversa, sintetizzano urea a partire dall’ammoniaca con un processo che richiede un certo dispendio
energetico. L’ureotelismo presenta però dei vantaggi: la ritenzione di urea (sostanza poco
tossica) da parte di alcuni Vertebrati marini contrasta la disidratazione per osmosi; inoltre
l’urea, essendo meno tossica dell’ammoniaca, può essere concentrata nell’urina consentendo
così il risparmio di acqua. L’ureotelismo è comparso più volte nella storia dei Vertebrati.
Forse si sviluppò una prima volta nei pesci di acqua dolce per evitare la disidratazione osmotica quando conquistarono i mari. In altri casi l’ureotelismo può aver rappresentato un preadattamento alle terre emerse, rendendo possibile trattenere l’azoto in forma poco tossica fino
a che non fosse disponibile una quantità di acqua sufficiente per la sua escrezione. La maggior
parte dei sauropsidi converte l’ammoniaca in acido urico (uricotelici), un adattamento importante per animali che devono risparmiare acqua: l’acido urico è, infatti, poco solubile in
acqua. L’urina dei sauropsidi è una soluzione moderatamente diluita di acido urico e di altri
sali, isosmotica rispetto ai liquidi interni. L’acido urico tende, però, a precipitare sotto forma
di cristalli di urati nei dotti escretori, nell’uretere e nella vescica. Quando l’acido urico preci12.1
Omeostasi
343
pita dalla soluzione, l’urina diviene meno concentrata, quindi si libera acqua che può ancora
essere riassorbita; l’escrezione di cataboliti azotati sotto forma di acido urico comporta, dunque, un ulteriore risparmio di acqua. L’uricotelismo si è, però, probabilmente instaurato per
un’altra necessità dei sauropsidi, legata alla riproduzione. Questi Vertebrati, infatti, si riproducono mediante uova cleidoiche in cui l’embrione è isolato dall’ambiente esterno non
solo da gusci protettivi, ma anche dagli annessi embrionali: in un ambiente così chiuso l’ammoniaca porterebbe rapidamente a morte l’embrione. L’uricotelismo permette, invece, di accumulare i prodotti del catabolismo azotato embrionale in forma semisolida e inattiva nel
sacco dell’allantoide senza pericolo di intossicazione per l’embrione.
MAMMIFERI – Il rene dei Mammiferi (così come, in parte, quello degli Uccelli) deve affrontare il problema del risparmio idrico e, contemporaneamente, quello di eliminare
enormi quantità di metaboliti azotati (prevalentemente sotto forma di urea) che derivano dal
tachimetabolismo connesso all’omeotermia. Per far ciò il rene di Mammifero deve filtrare
ogni giorno un volume molto elevato di sangue. In un uomo si calcola che vengano filtrati
circa 1500 litri di sangue al giorno (1 litro al minuto) con produzione di circa 170 litri di ultrafiltrato. Poiché la quanatità di acqua introdotta è pari a circa 2 litri/giorno (inclusa l’acqua
metabolica e quella introdotta con il cibo), se ne deduce che il rene del Mammifero deve riassorbire quasi tutto l’ultrafiltrato ed emettere solo 2 litri di urina al giorno per mantenere
l’equilibrio idro-salino. Per poter far fronte all’imponente ultrafiltrazione il rene dei Mammiferi è costituito da milioni di nefroni. L’ultrafiltrato riceve una prima rielaborazione a livello del tubulo contorto prossimale, ove viene operato un intenso riassorbimento attivo di
ioni cloro e sodio; grazie alla rimozione di NaCl, l’acqua può fluire attraverso l’epitelio del
tubulo verso l’esterno per osmosi ed il volume dell’ultrafiltrato diminuisce di circa un terzo.
Un’ulteriore rielaborazione avviene a livello dell’ansa di Henle, la novità anatomica introdotta proprio dai Vertebrati omeotermi. Il ramo discendente dell’ansa, rivestito da epitelio
sottile, passa attraverso tessuti con osmolarità crescente e l’acqua viene persa (circa 25%)
dall’ultrafiltrato per diffusione passiva. Lungo il percorso nel ramo ascendente dell’ansa non
vi è riduzione importante del volume, bensì un riassorbimento attivo di NaCl cosicché
l’urina che arriva al tubulo contorto distale risulta iposmotica rispetto ai tessuti circostanti e,
quindi, perderà acqua per osmosi con una riduzione del volume che può arrivare al 20%. La
fase finale della formazione dell’urina si realizza a livello dei tubuli collettori che sono permeabili all’acqua. Il riassorbimento idrico a livello di questa zona del rene è sottoposto a controllo ormonale: l’ormone antidiuretico ipofisario (ADH), a seconda delle necessità dell’animale, può aumentare e facilitare il riassorbimento attraverso l’epitelio dei collettori, portando il volume finale dell’urina a circa l’1% dell’ultrafiltrato glomerulare.
La chiave di volta per comprendere la perfetta struttura anatomica di questo rene in relazione alla sua efficienza consiste nel fatto che l’ansa di Henle ed i dotti collettori si spingono entro tessuti con osmolarità crescente. È, inoltre, fondamentale l’organizzazione
strutturale tra l’ansa di Henle ed i vasi che la circondano (vasi retti); questi elementi creano
una serie di tubi paralleli in cui il flusso dell’urina avviene in controcorrente rispetto al
flusso del sangue nei vasi (flusso in controcorrente). Il NaCl secreto dal ramo ascendente
dell’ansa diffonde nei tessuti della regione midollare aumentandono l’osmolalità e questi
sali in eccesso vengono distribuiti dal flusso in controcorrente in modo da mantenere un
forte gradiente osmotico all’interno della zona midollare che facilita il riassorbimento passivo di acqua (Figura 12.1).
344
CAPITOLO 12
Riconsiderazioni sull’evoluzione dei Vertebrati
Figura 12.1
Osmoregolazone renale
nel Mammifero
12.1.2 OMEOSTASI TERMICA
Le cellule di tutti gli organismi viventi sono dei sofisticati laboratori in cui sostanze chimiche diverse vengono continuamente trasformate attraverso reazioni metaboliche catalizzate
da enzimi. Dal momento che la temperatura influenza la velocità delle reazioni chimiche, la
sua importanza nei processi vitali è evidente. La velocità delle reazioni chimiche raddoppia
o triplica per un aumento di 10°C di temperatura e ciò significa che anche il metabolismo
può variare di questo ordine di grandezza per variazioni di temperatura. Dovrebbe quindi
essere vantaggioso per un animale riuscire a mantenere la temperatura corporea costante e
sufficientemente elevata da consentire reazioni chimiche veloci. È intuitivo che un animale
che possa permettersi le stesse capacità di movimento, attenzione, risposte agli stimoli di
giorno e di notte, d’estate e d’inverno indipendentemente dalla temperatura dell’ambiente
in cui vive, avrà dei vantaggi rispetto ad un animale la cui attività si innalza o si riduce in
funzione delle variazioni di temperatura esterna.
Tra i Vertebrati solo Uccelli e Mammiferi sono in grado di mantenere costante (e sufficientemente elevata) la temperatura corporea. Questo ha fatto sì che questi animali siano
12.1
Omeostasi
345
Penne - peli pannicolo adiposo sottocutaneo
Eterodonzia
Polmone
parenchimatoso
Cuore
quadricamerato
Ghiandole
salivari anteriorizzazione
digestione
Triturazione
cibo
Ghiandole gastriche
sviluppate
Ventriglio
Succo enterico
Villi
Rene:
midollare
con anse
di Henle
Isolamento termico
Digestione e assorbimento cibo
Assorbimento O2
Eliminazione scorie metaboliche
Trasporto nutrienti e scorie
Figura 12.2
Adattamenti legati
all’omeotermia
stati in grado di colonizzare anche le zone più inospitali ed estreme per valori di temperatura: dai ghiacci polari ai deserti più caldi. Uccelli e Mammiferi vengono classificati, per
questa loro capacità, come animali omeotermi in contrapposizione a tutti gli altri Vertebrati indicati come eterotermi. Alcuni studiosi utilizzano un’altra terminologia – endotermi (Uccelli e Mammiferi) ed ectotermi (gli altri Vertebrati) focalizzando l’attenzione
sulla sorgente di energia utilizzata per la termoregolazione. Gli ectotermi acquistano calore
da sorgenti esterne, ad esempio esponendosi al sole o riposando su rocce calde; gli endotermi mantengono alta e costante la loro temperatura corporea basandosi sulla produzione
metabolica di calore. Un animale endotermo può cambiare la quantità di calore che produce variando la velocità del metabolismo; in questo modo può mantenere costante la propria temperatura corporea in condizioni ambientali differenti regolando la produzione di
calore in modo tale che essa risulti pari alla perdita di calore del proprio corpo (omeostasi
termica). La produzione di calore non è, però, limitata al metabolismo basale (Figura 12. 2):
aumento di calore si verifica anche durante la digestione del cibo, dovuto al consumo di
energia legato ai processi di assimilazione, così come l’attività dei muscoli scheletrici, soprattutto durante la locomozione, è un’altra importante fonte di calore. Poiché gli endotermi vivono spesso in ambienti in cui la temperatura esterna è inferiore alla loro temperatura corporea, tendono a perdere calore verso l’ambiente; controbilanciare la perdita di calore è importante per ridurre il costo energetico dell’omeostasi termica. Uccelli e Mammiferi utilizzano il piumaggio ed il pelo come isolanti contro la perdita di calore. La possibilità
di formare un cuscinetto d’aria stagnante sopra la superficie corporea è un metodo per
creare uno strato isolante che impedisce il trasferimento di calore per conduzione. I peli o
le penne intrappolano l’aria e formano un cuscino di spessore variabile, in quanto può es346
CAPITOLO 12
Riconsiderazioni sull’evoluzione dei Vertebrati
sere regolato alzando o abbassando le penne o i peli. In alcuni Mammiferi l’aumento eccessivo della temperatura corporea è accompagnato da ipersecrezione delle ghiandole sudoripare: l’evaporazione del sudore provoca un abbassamento della temperatura corporea; altri
ansimano, respirando in modo rapido e poco profondo, in modo da aumentare l’evaporazione dell’acqua attraverso l’apparato respiratorio. Un altro modo per coibentare il corpo è
quello di fornirgli un cuscinetto di tessuto isolante, quale è il tessuto adiposo: ampi strati di
pannicolo adiposo nel tessuto sottocutaneo proteggono dalla perdita di calore. La vasodilatazione e la vasocostrizione della cute sono altri espedienti utilizzati per controllare la perdita di calore superficiale: la vasodilatazione è associata a perdita di calore, mentre la vasocostrizione riduce la perdita di calore attraverso la cute.
Regolare la temperatura corporea ad un valore elevato produce indubbi benefici, ma ha
un costo energetico molto alto. L’animale omeotermo può essere paragonato ad una stufa
in cui la produzione di calore è proporzionale al combustibile che viene bruciato. Bruciare
combustibile significa ossidarlo con una reazione chimica velocissima; non basta, dunque,
il combustibile: è necessaria una adeguata quantità di comburente, cioè di ossigeno. Anche
negli animali la produzione di calore è dovuta a reazioni di ossidazione del combustibile
(cibo), ma la reazione è più lenta perché controllata enzimaticamente.
Un animale omeotermo avrà bisogno di molto più combustibile e di molto più ossigeno
di un animale eterotermo per poter mantenere l’omeostasi termica, in altri termini il suo
metabolismo di base dovrà essere molto elevato (tachimetabolismo). Molti dei suoi apparati ed organi dovranno modificarsi per adattarsi a questa nuova condizione: l’apparato digerente dovrà garantire una maggiore efficienza nella digestione e nell’assorbimento degli
alimenti (triturazione del cibo, aumento della superficie intestinale assorbente); l’apparato
respiratorio dovrà essere in grado di fornire una adeguata quantità di ossigeno per ossidare
il cibo introdotto; l’apparato circolatorio dovrà garantire una rapida distribuzione del materiale energetico a tutte le cellule dell’organismo. L’elevato metabolismo di questi animali
produrrà anche grandi quantità di cataboliti tossici (CO2 e cataboliti azotati, soprattutto);
l’anidride carbonica verrà eliminata da un efficiente polmone, mentre i cataboliti azotati
avranno bisogno di un rene altamente specializzato per la loro eliminazione. In particolare,
il rene dovrà provvedere ad ultrafiltrare grandi quantità di sangue, aumentando il numero
di nefroni; ciò porterà, come conseguenza, alla necessità di rielaborare profondamente l’ultrafiltrato in modo da eliminare i cataboliti, ma riassorbire quasi tutta l’acqua. Ecco, quindi,
che omeostasi termica ed omeostasi idrico-salina devono trovare (e come abbiamo visto,
trovano) un modo per non intralciarsi a vicenda.
12.2 CRESTA NEURALE: UNA NOVITÀ CHE HA PERMESSO
L’EVOLUZIONE DEI VERTEBRATI
i
Come più volte accennato nei vari capitoli di questo libro, la cresta neurale è una popolazione di cellule assolutamente unica nel suo genere: si struttura durante la formazione del
tubo neurale come cordone compatto di cellule al confine tra neuroblasto ed epiblasto ma
le cellule componenti perdono in seguito adesione e vanno a costituire una popolazione di
cellule libere e migranti, destinate a raggiungere, secondo schemi ben determinati e nei
tempi opportuni, diversi organi in formazione, ove andranno a differenziare una moltitudine di derivati dalle più disparate caratteristiche citologiche e funzionali (Figura 12.3).
12.2
Cresta neurale: una novità che ha permesso l’evoluzione dei Vertebrati
347