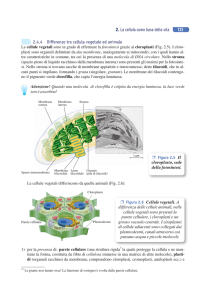
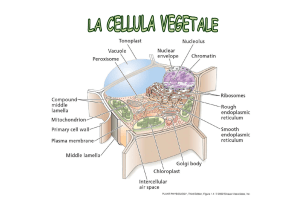
CELLULA VEGETALE
Non dobbiamo immaginarcela come qualcosa in più o in meno rispetto
alla cellula animale, ma come qualcosa di diverso.
Pur in un contesto di analogie morfologiche e metaboliche presenta
aspetti peculiari rispetto a una cellula animale:
9Assorbimento di H2O e soluti tramite pompe protoniche (H+-K+
piuttosto che Na+-K+).
9Fotorespirazione (cloroplasto-mitocondrio-perossisoma) in aggiunta
alla respirazione cellulare.
9Conversione dei lipidi in zuccheri mediante particolari perossisomi,
denominati gliossisomi.
9Formazione di una parete cellulare esterna, più o meno spessa e
rigida!
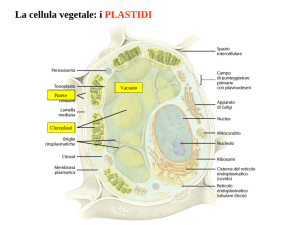
PLASTIDI
Sono organuli cellulari tipici e fondamentali delle cellule vegetali. Non li hanno solo
batteri e funghi eterotrofi e chiaramente tutte le cellule animali.
Non sono fondamentali alla vita della cellula come altri organuli quali i ribosomi ad
esempio, ma per le cellule vegetali sono assolutamente necessari per mantenere il
livello di autotrofia.
Sono coinvolti nei processi di fotosintesi e accumulo.
9Si distinguono in cloroplasti, cromoplasti e leucoplasti.
9Sono delimitati da una doppia membrana; costituiti
internamente da un sistema di membrane dette tilacoidi e da una
sostanza fondamentale detta stroma.
9All’interno delle membrane possono contenere i pigmenti
fotosintetici (clorofilla e carotenoidi).
9Sono organelli semiautonomi simili ai batteri (nucleoide con
DNA circolare; ribosomi 70S inibiti da antibiotici; divisione per
scissione).
ORIGINE ENDOSIMBIONTICA DEI
CLOROPLASTI
¾I presunti antenati dei cloroplasti erano
procarioti fotosintetici che divennero
endosimbionti, ossia viventi all’interno di
cellule più grandi eterotrofe, inizialmente
come prede non digerite o parassiti interni.
¾La cellula ospite sviluppò un sistema di
membrane interne e un citoscheletro che
inglobarono i procarioti fotosintetici isolandoli
all’interno di vescicole.
¾La simbiosi instaurata portò benefici per
entrambi gli organismi e la stretta
interdipendenza portò alla formazione di un
singolo organismo, antenato dei futuri
eucarioti, costituito da parti inseparabili.
da: “Biologia”
(N.A.
Campbell, J.B. Reece)
CLOROPLASTI
Sono gli organuli che effettuano la fotosintesi clorofilliana: immaginate che senza la loro
presenza non sarebbe possibile vivere su questo pianeta.
Sono organuli complessi di colore verde, il verde della clorofilla. Tale colore è una
caratteristica essenziale al ruolo svolto dalla clorofilla nell’assorbire alcune radiazioni
solari per svolgere il processo fotochimico.
Soprattutto nelle cellule dei parenchimi clorofilliani (foglie e giovani rami primari).
- Asse max 4-8 μ, asse min. 1-3 μ,
- tra 60 e 100 ma possono arrivare anche a 200 in una sola cellula.
- Sono capaci di dividersi indipendentemente dalla divisione del nucleo.
Infatti il loro numero può aumentare nel corso dello sviluppo in lunghezza della cellula.
Il loro processo riproduttivo sembra essere dipendente dalla esposizione alla luce, e
dalla presenza di alcuni ormoni del tipo della cinetina.
Le forme sono molto variabili (stella, pallina, nastro spiralato, ecc.), la loro dimensione si
può modificare ciclicamente, si possono spostare ed orientarsi in modo confacente
rispetto alla luce (variano di posizione in base anche alla qualità di luce che arriva).
All’interno del cloroplasto sono visibili delle aree più scure, - maggiore assorbimento di
luce e maggiore densità -, dette grana, che definiscono la porzione stazionaria interna
al cloroplasto, rispetto ad una sua parte mobile. Ciascun cloroplasto è delimitato da una
doppia membrana: la più esterna è facilmente permeabile dai metaboliti, la seconda
invece è più difficilmente attraversabile ed il passaggio dei metaboliti è regolato da
alcuni trasportatori.
I grana sono costituiti da un sistema ordinato di lamelle disposte parallelamente tra loro
ed orientate secondo l’asse maggiore del plastidio. Queste lamelle sono sacculi
schiacciati, delimitati da membrana ai quali è stato dato il nome di tilacoidi.
Per le loro dimensioni si distinguono in due tipi:
tilacoidi dei grana e tilacoidi intergrana (o stromatico).
Gli intergrana sono molto lunghi
longitudinalmente tutto il cloroplasto.
e
percorrono
Quelli dei grana invece sono più piccoli, della forma di una
monetina di 0,3-1 micron, sono sovrapposti l’uno all’altro
come a formare una pila cilindrica detta appunto grana.
Si va dai 40 agli 800 grana per cloroplasto.
Le membrane dei tilacoidi contengono il più dei lipidi di un
cloroplasto, i vari pigmenti fotosintetici, e la metà delle
proteine: infatti qui si svolgono le cosi dette
reazioni luminose della fotosintesi, le reazioni del
trasporto di elettroni fotosintetico (fotosistema I e II)
e la fosforilazione ad esso associata.
All’interno del cloroplasto vi è una matrice chiamata stroma o sostanza fondamentale,
un compartimento idrofilo ricco di proteine enzimatiche (tra le quali anche quelle della
fissazione e riduzione della CO2).
Nello stroma del cloroplasto sono presenti numerosissimi ribosomi, più piccoli di quelli
presenti sul citoplasma. Sono anch’essi in grado si sintetizzare proteine e per alcuni
caratteri che riguardano i meccanismi di assemblaggio polipeptidico, assomigliano molto
ai ribosomi dei batteri. Sottoforma di fibrille sottilissime è presente anche il DNA.
Gli zuccheri che si formano in seguito alla fotosintesi vengono subito traslocati in tutte le
porzioni periferiche della pianta: spesso però la velocità di produzione è troppo elevata
rispetto alle possibilità di trasporto per cui una parte degli zuccheri viene trasformata in
amido primario o di assimilazione.
Solitamente nei cloroplasti
sono presenti uno o più
granuli di amido. Tali
granuli non hanno vita
lunga: di notte vengono
idrolizzati e gli zuccheri che
ne
derivano
vengono
subito
trasportati
nei
parenchimi di riserva. A
volte sono rinvenibili anche
delle gocce lipidiche.
In merito alla struttura interna dei
cloroplasti si è osservato che i
tilacoidi che collegano i vari grana
sono costituiti da un sistema di
canali membranosi, visualizzabile a
livello
tridimensionale.
Diversi
modelli tridimensionali sono stati
proposti per la spiegazione di
queste strutture.
La membrana dei tilacoidi presenta una
struttura granulare, costituita da un
foglio bidimensionale di sottounità,
ciascuna di circa 120-175 Angstrom(Å).
Si pensa che ciascuna di queste
sottounità, di forma romboidale,
potesse
essere
l’espressione
morfologica dell’unità fotosintetica,
detta quantosoma.
Tali unità presentano sempre una certa
disposizione ordinata, più o meno
differente a seconda dei tipi di piante.
Queste particelle, quantosomi, sono
immerse in un'unica matrice lipidica del
classico tipo bistratificato: le particelle
sono spesso leggermente più spesse
della membrana fosfolipidica, ed è per
questo che la superficie delle lamelle
appare rugosa.
La composizione chimica dei cloroplasti è stata ottenuta attraverso il loro isolamento:
40-60% di proteine, 20-30% di lipidi, 5-10% di clorofilla (oltre il 75% clorofilla a), 24% di carotenoidi, 2-7% di acidi nucleici, 7-10% di residuo minerale.
Il pigmento più importante presente nei cloroplasti è la clorofilla, che si distingue in
diverse tipologie. Il tipo a, il primo che si è evoluto ed è presente in tutte le alghe e nelle
piante superiori, la clorofilla b invece è presente solo in alcune alghe, le più evolute ed in
tutte le piante superiori, Briofite comprese.
La clorofilla a è costituita da una doppia struttura: un anello porfirinico, costituito da
quattro anelli pirrolici con al centro un atomo di magnesio, ed un composto a 20 atomi
di carbonio di natura alifatica (catena lipidica) detto fitolo.
La b è simile in tutto tranne che per la sostituzione dei gruppi carbossilici (CH3) con dei
gruppi aldeidici sull’anello porfirinico.
Le due clorofilla presentano spettri di assorbimento piuttosto diversi: i picchi di
assorbimento che si raggiungono nelle bande del rosso e dell’azzurro sono sfalsati.
Tali molecole si trovano nelle membrane dei tilacoidi, legate sia alle proteine (anello
porfirinico), sia ai lipidi (fitolo).
da: “Biologia delle piante” (P.H. Raven et alii)
da: “Biologia delle
piante” (P.H. Raven
et alii)
Le clorofilla e i pigmenti carotenoidi si trovano
sulle membrane dei tilacoidi e sono responsabili
della cattura della luce nella fotosintesi. Nei
cloroplasti l’energia solare è convertita in energia
chimica e la CO2 fissata sotto forma di carboidrati,
amminoacidi, acidi grassi; in essi viene accumulato
temporaneamente l’amido (amido primario o
fotosintetico).
Nei cloroplasti oltre alle clorofilla sono presenti anche i carotenoidi: luteina, b-carotene,
a-carotene, violaxantina, neoxantina. Essi si rendono manifesti in autunno, quando la
clorofilla viene distrutta. Hanno il ruolo: a) di proteggere la clorofilla da processi di
fotodistruzione b) assorbire e trasferire energia luminosa alla clorofilla a. Anche i
chinoni, come il plastochinone sono presenti in abbondanza nei cloroplasti, e sono
presenti sia sui tilacoidi che nello stroma. La loro concentrazione sembra essere
notevolmente influenzata dalla intensità luminosa.
PROPLASTIDI
Rappresentano
lo
stadio
giovanile dei plastidi e si
trovano nelle cellule ancora
indifferenziate dei meristemi
primari.
Di
struttura
semplicissima
(dimensioni variabili da 0,5 ad
1µ), sono delimitati da una
doppia membrana, con uno
stroma interno privo di sistema
lamellare.
Posseggono
la
protoclorofilla, collocata in un
complesso proteico.
Essi
sono
capaci
di
sintetizzare
amido,
ma
comunque non producono
zuccheri. Presentano numerosi
ribosomi e fanno normale
sintesi proteica e sintesi di
acidi grassi.
La trasformazione dei proplastidi in cloroplasti è strettamente dipendente dalla luce, ed
avviene a partire dalle cellule meristematiche dell’apice del germoglio. La luce non è
l’unico arbitro a dettare le condizioni, come per esempio nelle cellule dell’epidermide (si
formano si dei cloroplasti, ma molto rudimentali e con scarsissima efficienza
fotosintetica) che ovviamente hanno un ruolo protettivo.
- I proplastidi che si differenziano dalle cellule del meristema radicale si trasformano
automaticamente in amiloplasti, cioè organuli adibiti alla riserva di amido. Se però le
radici vengono fatte crescere alla luce essi si trasformano in cloroplasti, anche se non
mostrano mai un sistema lamellare così organizzato come quello dei cloroplasti del
fusto.
Il passaggio da proplastidi a cloroplasti rappresenta quindi il raggiungimento della
capacità fotosintetica.
La membrana esterna del cloroplasto presenta un ruolo molto importante nella genesi
del sistema lamellare interno. Essa nell’area a contatto con lo stroma si introflette
originando delle invaginazioni (tilacoidi).
Entrano in gioco la sintesi di numerose proteine che avviene sia nel citoplasma che nello
stroma del plastidio. I lipidi sono anch’essi una componente importante dei tilacoidi, e
sembra che parallelamente alla loro sintesi il plastidio converta la protoclorofilla in
clorofilla.
Infine con la formazione dei grana si raggiunge la tappa conclusiva del differenziamento
delle membrane fotosintetiche = Infatti i tilacoidi contengono solo il fotosistema I,
mentre i grana sia I che il II.
Se i semi di Angiosperme
germinano al buio, si
originano piantine eziolate:
- forti scompensi;
- maggiore crescita degli
internodi;
- assenza di clorofilla.
La protoclorofilla non si
può
trasformare
(per
riduzione, aggiunta di due
atomi
di
idrogeno)
in
clorofilla.
Gli ezioplasti sono comunque più grossi dei
proplastidi. Si differenziano solo alcuni tilacoidi sparsi
ed un corpo di natura cristallina (strutture tubuliformi
disposte in un reticolo cubico o esagonale) detto Corpo
Prolamellare.
Nel CP è contenuta anche la protoclorofilla. Nello
stroma si trovano i ribosomi e qualche fibrilla di DNA.
I proplastidi, differenziandosi
in plastidi adulti presentano
uno sviluppo anormale e
vengono detti ezioplasti.
Essi rimangono organuli
indifferenziati
e
quindi
assomigliano
molto
ai
proplastidi.
Se esposti alla luce essi poi
diventano cloroplasti.
Differenziazione degli ezioplasti in cloroplasti
In alcune specie il CP si forma in condizioni naturali nell’arco di una notte (Cucumis
sativus e Phaseolus vulgaris) e scompaiono di giorno, mentre in molte altre esso non ha
il numero di ore sufficienti per formarsi in maniera naturale.
Illuminando giovani piantine eziolate, nel giro di 2-3 giorni queste diventano verdi e
acquistano la capacità di normale fotosintesi. Il CP si dissolve velocemente e diventa
anche il centro di organizzazione del sistema lamellare operativo dei tilacoidi e grana.
Riportando la piantina al buio si rinnesca il processo inverso, si riforma il CP. Tale
relazione però non produce sempre gli effetti descritti a tutti gli stadi di sviluppo dei
plastidi: nelle fasi di sviluppo giovanile tale reversibilità e bene evidente, ma tende a
ridursi quasi a zero nei plastidi che hanno raggiunto lo stadio maturo.
Sin dalle prime fasi di sviluppo compaiono delle file di vescicole, mentre nei pressi del
CP sono spesso presenti delle gocce lipidiche che verranno poi utilizzate per la
costruzione del sistema di lamelle fotosintetiche.
Leucoplasti
Si indicano così i plastidi incolori di una pianta. Sono organi che hanno principalmente il
ruolo di accumulare sostanze di riserva, e si trovano in particolare nei parenchimi
amiliferi dei semi, della radice (amiloplasti) e del fusto (ezioplasti). Per questo hanno
una elevata capacità sia di sintetizzare amido che, viceversa, di idrolizzarlo.
Sono molto più semplici dei cloroplasti. Presentano una doppia membrana lipoproteica,
e nello stroma un ridottissimo sistema di membrane (pochi tilacoidI).
Dal proplastidio aumentano gli enzimi del metabolismo dei carboidrati (aldolasi, 3fosfogliceraldeide-deidrogenasi);
Il più dello stroma di un amiloplasto adulto è occupato da granuli di amido. Come si sa
questo non è un carattere esclusivo degli amiloplasti (vedi cloroplasti) ma qui esso è
accumulato come sostanza di riserva.
Infatti qui esso è chiamato amido secondario, o di accumulo, in quanto deriva dagli
zuccheri provenienti dai cloroplasti, dove temporaneamente di giorno si può accumulare
un po’ di amido primario.
L’amido è la riserva glucidica vegetale per
eccellenza, ed in particolar modo esso è utilizzato
nel periodo della ripresa vegetativa.
I granuli di amido variano molto per numero forma e
dimensioni: nella patata troviamo i più grandi, nella
castagna i più piccoli.
Il centro di formazione dell’amido è chiamato ilo. La
forma piuttosto variabile e può essere posizionato al
centro dei granuli o in posizioni diverse. Se i centri di
accumulo sono due si formano i grani di amido
composti (riso e avena).
L’amido è un polimero del glucosio (legami alpha1-4). Lungo la catena dell’amido vi
sono delle ramificazioni (legami alpha1-6). Infatti la maggior parte degli amidi è
separabile in due componenti: l’amilosio (catena lineare di residui glucosidici alpha1-4)
e l’amilopectina (molecola ramificata di residui glucosidici con legami alpha1-4 e
ramificazioni alpha1-6). L’amilosio, per lo specifico legame alpha1-4 forma delle strutture
ad elica. Le due frazioni di amido si trovano nelle cellule in proporzioni diverse.
Trattando con
iodio, l’amilosio
si colora di blu,
l’amilopectina di
un rosso violaceo
meno intenso.
Amiloplasto con all’interno 2 granuli di amido con ilo asimmetrico; la matrice
è abbondante e la struttura lamellare ridotta.
In alcune Graminaceae e Compositae non si forma amido, ma fruttosani (es.
inulina), molecole più piccole dell’amido formate da catene di fruttosio; si
accumulano nel vacuolo e non nei leucoplasti.
La forma del granulo di
amido
(1-170
µm)
permette di risalire alla
specie che lo ha generato;
ha
quindi
significato
tassonomico
e
farmacognostico.
CROMOPLASTI
Sono plastidi colorati per l’abbondante presenza di pigmenti, tra cui i carotenoidi.
Questi plastidi sono presenti abbondantemente nei fiori e nei frutti.
Sono fotosinteticamente inattivi, e non hanno un particolare ruolo funzionale;
- doppia membrana, e nello stroma troviamo vescicole, tubuli, piccoli cristalli, lamelle,
gocce lipidiche ecc. Non presentando una struttura interna ripetibile.
- I pigmenti che prevalgono sono i carotenoidi, che nella Carota raggiungono il 20-25%
del peso secco; oltre a ciò il 58% è costituito da lipidi, 22% da proteine e 3,3% da RNA.
I cromoplasti derivano il più delle volte, dalla diretta trasformazione di cloroplasti o
amiloplasti (Peperoni, mele, Pomodori, Arance, ecc.), anche se in qualche caso si
originano direttamente dalla differenziazione dei proplastidi.
Una volta che il cromoplasto si è formato, indipendentemente dalla sua provenienza,
non può ritrasformarsi in cloroplasto: ciò confermerebbe la teoria che il cromoplasto
rappresenterebbe lo stato gerontico del plastidio.
Sono organelli sferici (0,5-1,4 μm) circondati da una
membrana singolaPEROSSISOMI
con una matrice interna che contiene
un corpo di natura proteica cristallino (raccolta di
molecole enzimatiche).
Giocano un ruolo importante nella fotorespirazione, un
processo che consuma O2 e libera CO2 (il contrario di
quanto avviene durante la fotosintesi). Nelle foglie verdi
sono strettamente associati ai mitocondri e ai cloroplasti.
Alcuni perossisomi (gliossisomi) contengono gli enzimi
necessari per la conversione dei grassi di riserva in
saccarosio durante la germinazione di molti semi.
da: “Biologia delle piante” (P.H. Raven et alii)
IL VACUOLO
Sono dei contenitori presenti all’interno della cellula solitamente di forma tondeggiante
delimitati da una doppia membrana lipoproteica chiamata tonoplasto.
Il tonoplasto è un tipo di membrana asimmetrica (proteine-lipidi-proteine) in quanto la
porzione proteica interna è più grossa di quella esterna. In entrambe le superfici, sono
presenti alcune particelle (60-150 Angstrom Å), la cui funzione non è ancora
perfettamente nota, ma si pensa siano proteine strutturali ed enzimatiche, importanti
nella regolazione del passaggio dei vari metaboliti.
E’ esclusivo delle cellule vegetali, ed in merito alla sua origine, si differenzia da
dilatazioni del reticolo endoplasmatico, ipotesi attualmente più accreditata.
Nelle cellule meristematiche il vacuolo è assente o poco sviluppato, mentre nelle cellule
che iniziano ad accrescersi nella prima fase di distensione alcune porzioni del reticolo
endoplasmatico iniziano a rigonfiarsi, prendendo il nome di provacuoli.
Quando inizia questa dilatazione, i ribosomi iniziano un processo di disancoramento
dalla membrana. Ciò non è ancora chiaro come avvenga, ma di fatto, quando il vacuolo
si è differenziato, non rimangono più ribosomi ancorati al tonoplasto.
Nel periodo giovanile, i vacuoli iniziano a differenziarsi in numero elevato ma con scarsa
rilevanza; nella fase di accrescimento per distensione acquisiscono sempre maggiori
dimensioni, tendono a fondersi tra loro e ad assorbire una notevole quantità di acqua. A
distensione terminata, il numero dei vacuoli e ridottissimo: molto spesso ne rimane uno
e di grosse dimensioni (anche l’80-90% delle dimensioni interne della cellula).
Qui si evidenzia un aspetto molto importante: la cellula vegetale aumentando di
dimensioni non sintetizza nuovo plasma, ma aumenta il volume dei suoi vacuoli. Nella
fase di crescita per distensione le cellule vegetali hanno solitamente un ritmo di sviluppo
notevole: non essendoci una corrispondente sintesi di plasma, il vacuolo si accresce in
proporzione anche per evitare che si creino vuoti d’aria.
- Un altro aspetto funzionale della presenza dei vacuoli sta nel fatto che essi aiutano a
mantenere un rapporto proporzionale tra superficie cellulare e plasma: il volume della
cellula aumenta in virtù dei vacuoli, il plasma rimane stabile ma confinato a contatto con
la membrana e quindi in grado di effettuare adeguati scambi con l’esterno.
- Oltre a ciò, non essendo sviluppato in questo tipo di cellule un sistema di escrezione
dei metaboliti secondari come negli animali, tali sacchi sono anche visti come magazzini
di derivati metabolitici secondari o di rifiuto della cellula vegetale.
- Essi funzionano anche come osmometri ed hanno così un ruolo importantissimo nella
regolazione dell’assorbimento di H2O nella cellula. Oltre a ciò, con i processi osmotici il
tonoplasto viene tenuto in tensione trasferendo alla cellula la necessaria turgidità.
- In quanto la parete cellulare è abbastanza rigida, di conseguenza nella cellula
vegetale si instaurano forti pressioni idrostatiche. Nel vacuolo possono essere
accumulate anche sostanze di riserva e proteine enzimatiche.
SUCCO VACUOLARE
E’ detto succo vacuolare perchè contiene la maggior parte del liquido cellulare; può
essere incolore, ma spesso è colorato perché ricco di flavonoidi, ed il pH è solitamente
acido.
In esso possono trovarsi una varietà molto ampia di sostanze: sali, proteine flavonoidi,
alcaloidi, sali inorganici, antociani, ecc. Ciò dipende molto dal tipo di cellula che
trattiamo (che funzione presenta), dalla specie e dall’ambiente. Tra le sostanze di
riserva, nel seme, sono presenti concrezioni ottenute per disidratazione dette granuli
aleuronici (essi contengono riserve fosfate – fitina – e proteine).
- idrati di carbonio, da mono a polisaccaridi, liquidi. Glucosio nell’uva, fruttosio nelle
pesche, Il saccarosio è abbondante nella barbabietola .
- sali inorganici, e tale aspetto dipende molto anche dall’ambiente in cui la pianta vive.
Cloruro di sodio, in ambienti salini, Calcio nei terreni calcarei.
Tra gli scarti o scorie del metabolismo abbiamo:
- Antociani, flavonoli e flavoni, importanti per la colorazione dei petali e dei frutti.
- Alcaloidi, Molti di essi sono delle basi, e si trovano sottoforma di sali di acidi organici.
Tra questi: Cocaina, papaverina, teina, morfina, caffeina, ecc.
- Acidi organici, come l’acido malico e l’acido citrico.
- Tannini: si accumulano spesso in cellule specializzate, colore brunastro. Sono
prodotti di polimerizzazione dei fenoli.
- Terpeni: sono composti derivati dell’isoprene. Si ritrovano in particolari cellule di
alcune specie alle quali conferiscono particolari aromi (es. pepe, zenzero, alloro).Tra i
terpeni sono rinvenibili anche le resine: esse tuttavia sono sostanze molto complesse,
che sono disciolte in terpeni.
Molto spesso nei vacuoli si accumula l’ossalato di calcio. Questo sale è
insolubile in acqua, per cui è sotto forma solida con strutture cristalline. La
cellula vuole spesso eliminare dall’ambiente metabolico l’acido ossalico e
acido ossalacetico, inibitore il primo di diversi enzimi plasmatici e il secondo
della respirazione cellulare.
Fenomeni osmotici:
- Sono determinanti per le cellule vegetali per l’assorbimento dell’acqua. Il sistema
vacuolare permette tali fenomeni, particolarmente importanti nella fase di crescita per
distensione. I fenomeni osmotici sono fenomeni di diffusione.
- Particelle di dimensione atomica o molecolare sono in continuo movimento: la loro
velocità è regolata dalla pressione e dalla temperatura. Spesso si creano delle
condizioni per imprimere direzioni preferenziali: le molecole tendono a spostarsi da zone
dove sono più concentrate a zone dove sono meno concentrate.
- Il movimento quindi delle molecole da zone a maggior concentrazione verso zone a
minor concentrazione prosegue sino a quando la sua concentrazione non diviene
uniforme in tutto l’ambiente. Tale fenomeno è detto di Diffusione. Una volta raggiunto
l’equilibrio le molecole non smettono comunque di muoversi, ma non hanno più una
direzione preferenziale.
- Molto più formalmente il movimento di particelle da zone a maggior concentrazione
verso zone a minor concentrazione, viene indicato anche come movimento che segue
un gradiente di potenziale chimico per una determinata specie chimica. Il movimento
delle molecole prosegue sino a quando il potenziale chimico dell’intero dipartimento
diventa uniforme.
Il potenziale chimico dell’acqua è influenzato dalla temperatura, dalla pressione e
dalle particelle e molecole in essa disciolte.
OSMOSI: la presenza delle particelle di soluto abbassa il potenziale chimico dell’acqua.
Il potenziale dell’acqua pura a pressione atmosferica è uguale a zero (Ψ0), e quindi
l’acqua con soluto avrà un valore di negativo (Ψ-a). l’acqua si muove per compensare
questo (ΔΨ).
Il movimento delle molecole d’acqua (solvente) attraverso una membrana
semipermeabile è detto fenomeno osmotico. Un sistema osmotico deve avere due
caratteristiche:
a) presenza di una membrana semipermeabile;
b) presenza di un dispositivo che consenta lo sviluppo di pressione (nell’esempio visto è
dato dalla pressione della colonna d’acqua).
Nelle cellule vegetali invece la pressione si sviluppa per la presenza di un involucro
elastico e resistente che è la parete cellulare. Lo sviluppo di pressione infatti
permetterà di controbilanciare il flusso di acqua verso il compartimento dove è meno
concentrata proprio perchè tale aumento di pressione farà innalzare il potenziale
chimico dell’acqua sino a portarlo a zero.
In termini pratici si usa definire la pressione osmotica come quella forza che deve
essere applicata sulla superficie di una soluzione (separata dal solvente puro da
una membrana semipermeabile) tale da controbilanciare il deficit di potenziale
rispetto al solvente ed impedirne conseguentemente l’ingresso per diffusione nel
compartimento della soluzione.
Come sappiamo all’interno delle cellule viventi numerosi organelli e comparti sono sede
di fenomeni osmotici: mitocondri, plastidi, reticolo endoplasmatico. Tuttavia, nelle
cellule vegetali, il vacuolo è quello che presenta i fenomeni più importanti per la vita
della cellula, e per la sua presenza diventa un osmometro.
Plasmolisi
Se messa a contatto con una soluzione isotonica, l’acqua entra ed esce dalla cellula in
ugual misura, ma se messa a contatto con una soluzione ipotonica, l’acqua in entrata è
superiore a quella di uscita, la cellula si rigonfia e si sviluppa la pressione osmotica a
causa della presenza della parete.
Una fuoriuscita d’acqua si avrà quando la cellula è posta a contatto con una soluzione
ipertonica, provocando così al suo interno un restringimento del circostante citoplasma
ed un progressivo distacco di questo dalla parete cellulare. Tale fenomeno prende il
nome di plasmolisi. Se poi tale tessuto di cellule plasmolizzato viene successivamente
immerso in acqua pura, la cellula riacquisisce al suo interno nuova acqua, si rigonfia il
vacuolo e si distende il plasma, dando origine al fenomeno della deplasmolisi.
Cellule normali
Cellule plasmolizzate
In questi due fenomeni il plasmalemma ed il tonoplasto si comportano rispetto ai soluti
in maniera diversa:
Il plasmalemma solitamente lascia passare facilmente anche gli ioni di Sali inorganici
mentre nel tonoplasto questo avviene più difficilmente o almeno con maggiore lentezza.
Questo comporta a volte che in fenomeni di plasmolisi si verifichi contemporaneamente
un certo rigonfiamento del plasma.
La parete cellulare controlla i fenomeni osmotici: nel caso visto di un tessuto messo a
contatto con una soluzione ipotonica, essa tende a regolare la pressione osmotica,
ovvero imprime al plasma quella pressione che serve ad un certo punto ad innalzare il
potenziale dell’acqua della cellula sino ad uguagliarlo a quello della soluzione ipotonica
esterna, di modo che cessi il flusso di acqua dall’esterno all’interno.
A questo punto la cellula è detta turgida, perché la pressione di turgore (PT) è in questa
fase identica alla pressione osmotica (PO). La tensione di assorbimento (TA) della
cellula, che è data dalla differenza dei due suddetti valori (TA=PO-PT) è ora uguale a
zero.
Il turgore è determinante per la vita della delle cellule e della pianta: le cellule si
accrescono solo quando sono turgide; gli stomi si aprono solo quando le cellule di
guardia sono turgide.