
1
Agli inizi del 1900, l'istologo S.R. Cayal impiegando la tecnica di colorazione con argento (metodo di golgi)
dimostrò che i neuroni all'osservazione microscopica risultano unità distinte, malgrado l'opinione allora corrente
fosse di ritenere il tessuto nervoso una struttura reticolare continua.
Le prove risolutive della discontinuità fra neuroni e dell'esistenza di zone di interazione tra neuroni diversi, le
sinapsi, si ebbero con l'avvento della microscopia elettronica negli anni '40.
Ben prima, nel 1906, C. Sherrington, considerato il fondatore della moderna neurofisiologia, affermava:
" il neurone stesso é visibilmente un continuum da un capo all'altro, ma non si arriva a dimostrare una continuità
là dove si incontrano neurone con neurone, ossia la sinapsi. Qui si può avere un tipo diverso di trasmissione“.
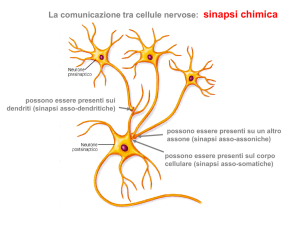
La sinapsi è la regione attraverso la quale un impulso nervoso si trasmette da una cellula ad un’altra. Esistono 2 tipi
di sinapsi: le sinapsi elettriche e quelle chimiche. Nelle sinapsi elettriche due cellule eccitabili comunicano per
passaggio diretto di corrente attraverso le gap-junction. Nelle sinapsi chimiche l’arrivo di un potenziale d’azione
nella cellula pre-sinaptica provoca la liberazione di un neurotrasmettitore il quale determina la risposta elettrica
nell’elemento post-sinaptico.
2
Il trasferimento di segnali elettrici tra cellule eccitabili ha luogo in regioni
specializzate dette sinapsi, la cellula che invia il segnale è detta cellula
presinaptica, la cellula che riceve il segnale è detta cellula postsinaptica. Lo
spazio che fisicamente separa le due cellule comunicanti è lo spazio
intersinaptico o fessura sinaptica.
Nella sinapsi l’efficienza di trasferimento del segnale è garantita sia dalla
stretta vicinanza tra cellula presinaptica e cellula postsinaptica (la fessura
sinaptica varia tra 2-4 a 30-50 nm) sia dall’ampia superficie che le due cellule
utilizzano per la comunicazione.
3
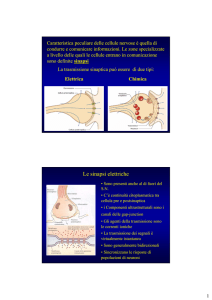
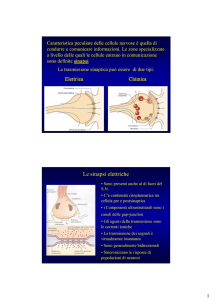
Sulla base delle strategie di comunicazione le sinapsi vengono suddivise in
elettriche e chimiche.
4
Le sinapsi elettriche sono localizzate nel snc, nel tessuto muscolare liscio e
cardiaco e nei tessuti neuroendocrini.
Nelle sinapsi elettriche le cellule sono separate da una fessura sinaptica di 2-4
nm, quindi così stretta da poter essere considerato uno spazio virtuale. Infatti
si può anche affermare che tra le cellule pre- e postsinaptiche ci sia una vera e
propria continuità elettrica garantita dalla presenza di giunzioni comunicanti
(gap junction)
5
Sono delle regioni di membrana contenenti particolari canali acquosi detti
connessoni. Ad ogni connessone della cellula presinaptica corrisponde un
connessone della cellula postsinaptica.
Ogni connessone è costituito da 6 subunità proteiche denominate connessine
che si dispongono in circolo e delimitano un canale acquoso centrale. Le
connessine sono una famiglia di proteine con un PM tra 26 e 56 kDa costituite
da 4 segmenti transmembrana. Strutturalmente ogni polipeptide di connessina
attraversa la membrana plasmatica 4 volte e una alfa elica trans-membrana delimita il
canale acquoso attraverso il quale possono passare molecole di PM non superiore a
1500.
Nel tessuto miocardico sono presenti queste giunzioni che assicurano la diffusione del
calcio da una cellula ad un’altra coordinando il battito di tutte le cellule muscolari di
una determinata regione cardiaca.
6
Il processo di apertura del connessone richiede un cambio conformazionale di
tutte le 6 connessine che lo compongono che probabilmente consiste in una
rotazione di tutte le subunità rispetto all’asse maggiore del canale ed è
modulata dal pH intracellulare, dagli ioni Ca2+, da secondi messaggeri e dai
neurotrasmettitori.
Generalmente esse si chiudono in presenza di stimoli che possono
determinare danno cellulare come diminuzione del pH, aumento della
concentrazione intracellulare di Ca2+, variazioni anomale del potenziale di
membrana. Quindi si comportano come vere e proprie porte in grado di isolare
completamente le cellule danneggiate dalle cellule circostanti, preservando
l’integrità del tessuto.
7
Quando connessoni attigui sono aperti vi può essere flusso di corrente tra
cellula pre e postsinaptica. Il risultato è che quando in una cellula presinaptica
insorge un potenziale d’azione si crea una ddp tra gli elementi sinaptici e
quindi una f.e.m. che induce l’insorgenza di correnti elettrotoniche che si
propagano dalla cellula con potenziale più negativo attraverso le giunzioni
comunicanti. Il risultato è una progressiva depolarizzazione della cellula
postsinaptica che se è tale da raggiungere il valore soglia scatena un
potenziale d’azione. Quindi la trasmissione elettrica si basa sulla creazione e
trasmissione di correnti elettrotoniche che sarà tanto più efficiente quanto
minore è la resistenza elettrica. Ciò è garantito dalla stretta vicinanza tra gli
elementi sinaptici, dall’estensione della giunzione comunicante e dall’elevato
numero di connessoni.
Le sinapsi elettriche possono far passare corrente in entrambe le direzioni per
questo.
8
La trasmissione attraverso la sinapsi elettrica è molto rapida perché è il
risultato di un passaggio diretto di corrente dall’elemento presinaptico a quello
postsinaptico. Il ritardo con cui la cellula postsinaptica risponde producendo un
potenziale d’azione è di alcuni decimi di millisecondo. Per questo le sinapsi
elettriche sono localizzate in aree del snc ed in tessuti in cui le cellule devono
generare potenziali d’azione sincroni per assolvere al loro compito.
9
A livello di una sinapsi elettrica la membrana e pre e post-sinaptica sono giustapposte a formare delle giunzioni
comunicanti gap junction
La giunzione elettrica fra due neuroni consente alla corrente del circuito locale prodotta dal potenziale d'azione di
uno dei due neuroni di passare all'altro depolarizzandolo. Sono state descritte sinapsi elettriche che conducono
preferenzialmente in una direzione piuttosto che nell'altra rettificazione
Il passaggio di corrente é consentito dalle gap-junction che formano dei canali di comunicazione tra cellule.
Cellule elettricamente accoppiate possono esser disaccoppiate per chiusura dei canali del connessone a causa di
aumento concentrazione di calcio, protoni ed in risposta a depolarizzazione. Sono presenti a livello del S.N.C., in
cellule non nervose come gli epatociti, le cellule muscolari liscie dell'intestino, le cellule del miocardio sono
accoppiate mediante gap-junction
10
Nel caso delle sinapsi chimiche è presente una fessura sinaptica relativamente
ampia (20-40 nm) e sono presenti specializzazioni strutturali che consentono
di distinguere l’elemento presinaptico da quello postsinaptico. La cellula
presinaptica è caratterizzata da un apparato esocitotico e dalle vescicole
sinaptiche. La cellula postsinatica è contraddistinta, invece, dalla presenza di
molecole recettoriali per il neurotrasmettitore localizzate nella regione della
membrana plasmatica posta di fronte alla cellula presinaptica. Questa
specializzazione dell’elemento pre e postsinaptico rendono la trasmissione
mediante sinapsi chimica unidirezionale.
11
Gli eventi che si susseguono durante la trasmissione in una sinapsi chimica
possono essere riassunti in tre fasi principali:
1. esocitosi del neurotrasmettitore nella fessura sinaptica
2. attivazione della membrana postsinaptica
3. rimozione del neurotrasmettitore dalla fessura sinaptica
12
La liberazione del neurotrasmettitore nella fessura sinaptica è indotta dal potenziale d’azione
generato dalla cellula presinaptica e dalla variazione della concentrazione intracellulare di
Ca2+. Ciò è reso possibile grazie alla presenza di canali per il Ca2+ voltaggio-dipendenti sulla
membrana dell’elemento presinaptico. In particolare, aprendosi in risposta al potenziale
d’azione questi canali causano un massiccio ingresso di ioni Ca2+ nel citoplasma che da il via
all’esocitosi del neurotrasmettitore contenuto nelle vescicole sinaptiche.
L’esocitosi del neurotrasmettitore avviene solo in aree specializzate della membrana
presinaptica, dette zone attive, dove le vescicole sinaptiche sono guidate dagli elementi
citoscheletrici attraverso un processo Ca2+ dipendente.
La natura chimica dei neurotrasmettitori è diversa a seconda della sinapsi chimica considerata
(acetilcolina, glutammato, glicina, dopamina, acido -amino butirrico, adrenalina, ecc) ma in
tutti i casi il neurotrasmettitore è una molecola con basso PM, permettendo al
neurotrasmettitore di diffondere velocemente nella fessura sinaptica.
13
14
15
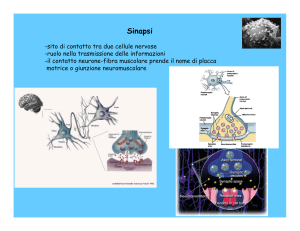
La giunzione neuromuscolare o placca motrice è stata la prima sinapsi dei
vertebrati ad essere studiata in dettaglio.
La trasmissione chimica si produce attraverso una fessura sinaptica, spazio di circa 20-30 nm che
separa le cellule pre e post-sinaptica.La terminazione pre-sinaptica contiene le vescicole sinaptiche di
circa 40 nm di diametro contenenti circa 10.000-50.000 molecole di trasmettitore ognuna.
16
17
18
Nelle sinapsi chimiche il neurotrasmettitore liberato dall’elemento presinaptico evoca
la risposta elettrica.
I neurotrasmettitori (acetilcolina, noradrenalina) contenuti in vescicole (50 nm di
diametro) localizzate in zone attive sotto membrana; i neuropeptidi contenuti in
vescicole + grosse in tutto il terminale assonico
Il potenziale d’azione nell’elemento pre-sinaptico provoca apertura di canali voltaggio
dipendenti del calcio
Entrata di calcio nel citosol
Esocitosi del neurotrasmettitore nel vallo sinaptico
Diffusione del neurotrasmettitore e legame a recettori della membrana post-sinaptica
Variazione della conduttanza della membrana post-sinaptica (EPSP; IPSP)
Recettori per molti neurotrasmettitori sono canali ionici
Neuropeptidi possono essere neuromodulatori
L’azione dei neurotrasmettitori si interrompe quando vengono riassunti dall’elemento
pre-sinaptico
Trasmissione unidirezionale, ritardo sinaptico
19
La membrana delle vescicole sinaptiche contiene le ATPasi di tipo V che
generano basso pH intravescicolare che viene utilizzato per concentrare nelle
vescicole il neurotrasmettitore tramite un sistema di controtrasporto. Le
vescicole sono associate al citoscheletro tramite la sinapsina. I topi knockout
privi del gene della sinapsina sono predisposti ad attacchi epilettici in quanto si
riduce il numero delle membrane che si fondono con la membrana plasmatica.
La fosforilazione della sinapsina da parte della CAM-chinasi II a sua volta
attivata dal complesso calcio-calmodulina formatosi in seguto all’entrata di
calcio, provoca la dissociazione delle vescicole sinaptiche dal citoscheletro,
aumentando il numero di vescicole che si può fondere con la membrana
plasmatica.
20
Il rilascio del neurotrasmettitore dalle vescicole sinaptiche si verifica solo in
alcune zone attive della membrana del terminale assonale. Le vescicole
sinaptiche vengono trasportate verso la zona attiva si fondono con la
membrana dell’assone solo quando le proteine delle vescicole interagiscono
con le proteine della membrana cellulare. Il processo coinvolge diverse
proteine, come la sinaptofisina, VAMP e SNAP, Rab3A. La Rab3A è
fondamentale per la localizzazione delle vescicole nella zona attiva. La VAMP
o sinaptobrevina si lega strettamente alla sintaxina e alla SNAP25. Le proteine
SNAP e NSF intervengono nella dissociazione. La tossina botulinica B
costituita da 2 polipeptidi di cui 1 è una proteasi che scinde la VAMP
impedendo il rilascio del neurotrasmettitore e quindi la paralisi. La
sinaptotagmina I che ha quattro siti di legame per il calcio sembra essere il
meccanismo calcio-sensibile che avvia l’esocitosi
21
22
Il neurotrasmettitore può essere ricaptato dal terminale assonico ma anche
alle cellule gliali. Può legarsi al recettore dell’elemento post-sinaptico,
sottoposto all’azione di enzimi o può essere riversato nel sangue come l’ADH
e svolgere la funzione ormonale
23
Il numero di molecole identificate come neurotrasmettitori è superiore al
centinaio ed è in continua crescita.
•I neurotrasmettitori sono sintetizzati nel terminale presinaptico;
•Sono impacchettati in vescicole e rilasciati per esocitosi;
•Diffondono nello spazio intersinaptico e si legano a specifici recettori posti
sulla membrana postsinaptica, alterandone in modo transiente le proprietà
elettriche;
•Si legano con alta selettività ai recettori postsinaptici, attivandoli;
•La loro azione è bolccata da antagonisti recettoriali specifici;
•La loro azione termina per degradazione enzimatica e/o ricaptazione nel
terminale presinaptico o nelle cellule gliali tramite trasportatori.
24
Acetilcolina: idrolizzata a colina e acetato da acetilcolinesterasi presente sulla superficie della
membrana post-sinaptica; l'eserina é un agente anticolinesterasico che ritarda l'idrolisi dell'aceticolina;
Ammine biogene:
Dopamina, norepinefrina (oradrenalina) ed epinefrina (adrenalina) hanno una via biosintetica comune a
partire dalla tirosina; serotonina ed istamina
Aminoacidi: glicina ( di tipo inibitorio), glutammato ed aspartato (eccitarori), acido gamma
amminobutirrico (GABA) con ruolo inibitorio.
Neuropeptidi:
Ormone antidiuretico e oxitocina appaiono coinvolti nei meccanismi della memoria; sostanza p, presente
nei neuroni sensitivi spinali, agisce nei meccanismi nervosi del dolore; livelli elevati di colecistochinina
nel cervello causano sazietà;l'angiotensina stimola il bere; l'ormone LH-rilasciante stimola il
comportamento sessuale
25
Gli enzimi necessari per la sintesi di questi trasmettitori vengono prodotti nel
corpo cellulare e trasportati alla terminazione nervosa mediante trasporto
assonico lento (2). Nella terminazione assonica, i precursori dei
neurotrasmettitori sono trasportati nel citoplasma per mezzo di trasportatori
specifici. In questa sede il neurotrasmettitore a basso PM viene sintetizzato,
impacchettato in vescicole e poi liberato per esocitosi.
Questi neurotrasmettitori vengono liberati molto velocemente, entro 1 ms
dall’arrivo del potenziale d’azione.
Generalmente le vescicole contenenti il neurotrasmettitore a basso peso
molecolare sono ancorate alla membrana presinaptica, per cui un aumento di
Ca localizzato in prossimità dei canali del Ca ne favorisce il rapido rilascio.
26
27
28
29
I neuropeptidi sono contenuti in vescicole di dimensioni maggiori (60-120 nm
di diametro). Sono sintetizzati come pre-propeptidi nel reticolo endoplasmatico
ruvido dove la sequenza segnale che indica il peptide da secernere viene
rimossa. Il propeptide risultante viene impacchettato in vescicole a livello
dell’apparato del Golgi. All’interno della vescicola avviene la completa
maturazione del neurotrasmettitore. Le vescicole vengono poi trasportate al
terminale assonico mediante trasporto assonico rapido.
A differenza dei trasmettitori a basso PM, quelli peptidici sono degradati
enzimaticamente e non sono ricaptati da trasportatori.
Le sostanze peptidiche che agiscono come neurotrasmettitori sono centinaia,
tra cui: oppiacei endogeni (endorfine, encefaline, dinorfine) che mediano
l’analgesia.
30
31
32
33
34
Recettori per i neurotrasmettitori
Recettori per GABA e glicina presentano significative omologie con quello nicotinico dell'acetilcolina pur
avendo funzioni inibitorie, mediando l'ingresso di cloro in cellula.
Sono stati identificati due tipi di recettori per GABA, denominati GABAA (identificate finora 5 diverse
sub-unità, che combinandosi fra loro possono formare canali omeomerici) e GABAB (quest'ultimo
modula indirettamente un canale che non é parte integrante del recettore)
I recettori GABA sono sede di azione di due importanti classi di sostanze farmacologiche:
1) le benzodiazepine (diazepam) farmaci anti ansia
2) barbiturici
attivano l'apertura del canale GABA dipendente.
35
La dopamina, la noradrenalina e l’adrenalina sono catecolamine che hanno in
comune la stessa via biosintetica che parte dall’aminoacido tirosina. La
Tirosina viene convertita a L-dopa dalla tirosina idrossilasi. L-Dopa è poi
convertita a dopamina da una specifica decarbossilasi; nei neuroni
dopaminergici la via biosintetica termina con questa reazione. I neuroni
noradrenergici possiodono un altro enzima la dopamina-beta-idrossilasi che
trasforma la dopamina in noradrenalina che rappresenta il principale
neurotrasmettitore dei neuroni sinaptici post-gangliari.
La serotonina è presente in numerosi neuroni che partecipano alla regolazione
della temperatura.
36
La velocità di trasmissione di una sinapsi chimica dipende dal tipo di molecole
recettoriali della membrana postsinaptica. Quando il neurotrasmettitore si lega
ai cosiddetti recettori ionotropi, quindi a canali ligando-dipendenti, la variazione
del potenziale della membrana postsinaptica inizia nel momento in cui i canali
si aprono. La trasmissione sinaptica è quindi veloce.
Se il neurotrasmettitore si lega ad un recettore metabotropo, una cascata di
eventi intracellulari si interpone tra il legame del neurotrasmettitore e la
variazione del potenziale della membrana postsinaptica ed il segnale elettrico
viene generato in ritardo dalla cellula postsinaptica. Il risultato è una
trasmissione sinaptica lenta.
37
Molti neurotrasmettitori generano risposte rapide e di breve durata aprendo
canali ionici (potenziale sinaptico veloce che dura pochi millisecondi). Alcuni
neurotrasmettitori innescano risposte lente e durature attivando sistemi di II
messaggeri. Queste risposte comprendono sia l’apertura che la chiusura di
canali ionici (potenziale sinaptico lento che può durare anche secondi o minuti)
ma non si limitano a questo, infatti possono modificare proteine esistenti o
determinare la sintesi di nuove proteine
38
39
40
41
Pur essendo le sinapsi chimiche più lente di quelle elettriche e sottoposte al
fenomeno dell’affaticabilità della sinapsi che si verifica quando la quantità
di neurotrasmettitore è insufficiente rispetto a quello richiesto, la scelta
evolutiva ha premiato soprattutto le sinapsi chimiche negli organismi
superiori e questo essenzialmente per tre cose:
1. AMPLIFICAZIONE DEL SEGNALE: l’esocitosi
anche di un’unica vescicola sinaptica garantisce
la liberazione di migliaia di molecole di
neurotrasmettitore e l’attivazione di migliaia di
recettori a livello postsinaptico. A differenza delle
sinapsi elettriche, la trasmissione intercellulare
mediante sinapsi chimica può avvenire anche tra
cellule di diverse dimensioni.
2. MANTENERE O INVERTIRE IL SEGNO DEL
SEGNALE ELETTRICO PRESINAPTICO: il
42
neurotrasmettitore rilasciato da una
depolarizzazione presinaptica può generare nella
cellula postsinaptica o una depolarizzazione o
una iperpolarizzazione a seconda del tipo di
canale postsinaptico che il neurotrasmettitore
apre.
3. SOMMAZIONE TEMPORALE O SPAZIALE:
potenziali postsinaptici ravvicinati in termini di
tempo o di spazio possono sommarsi a livello
postsinaptico generando un segnale elettrico
complessivo più efficace dei singoli segnali.
42
43
Recettore nicotinico per l'acetilcolina
E' stato il primo canale ionico a essere purificato, sequenziato, ricostituito in doppi strati sintetici,
registrato il segnale elettrico di un singolo canale; ne é stato clonato e sequenziato il gene; con tecniche
di microscopia elettronica e diffrazione dei raggi x é stata determinata la forma e la probabile
disposizione delle sue cinque sub-unità nella membrana.
Ciò é stato reso possibile dalla grande disponibilità del recettore nell'organo elettrico delle torpedini, e
dall'uso dell'alfa-bungarotossina (BuTx), fluorescente o radiomarcata, che si lega specificamente ed
irreversibilmente al recettore. Non é molto selettivo e si lascia attraversare, allo stato aperto da Na+, K+
e anche dal Ca2+.
44
45
46
47
E' stato possibile dimostrare l'esistenza del potenziale post-sinaptico o potenziale di placca (giunzione
neuromuscolare) utilizzando il curaro (d-Tubocurarina) che blocca i recettori nicotinici per l'acetilcolina,
ed impedisce l'apertura dei canali post-sinaptici.
Il potenziale d'inversione
Il potenziale d'inversione di un mediatore rappresenta il massimo potenziale di membrana prodotto
dall'apertura dei canali attivati da quel mediatore;
superato il quale mediante tecniche sperimentali, l'attivazione di tali canali produrrà una corrente di
direzione opposta.
esso dipende:
1) dalle conduttanze relative dei canali attivati per diversi ioni permeanti (selettività ionica del canale)
2) potenziali d'equilibrio degli ioni permeanti (basati sui loro gradienti di concentrazione)
Il concetto di potenziale d'inversione é essenziale per distinguere tra azioni sinaptiche eccitatorie ed
inibitorie; la sua determinazione sperimentale al variare delle concentrazioni di una determinata specie
ionica ci fornisce l'indicazione sulla composizione ionica di una determinata corrente.
48
49
50
51
52
53
54
55








![2Trasmissione sinaptica [modalità compatibilità]](http://s1.studylibit.com/store/data/004212424_1-cd4014b0b5591f026d28cd110d4215cc-300x300.png)