http://images.google.it/imgres?imgurl=http://www.atlantebotanica.unito.it/photos/Perixilematici4.gif&imgrefurl=http://www.atlante
botanica.unito.it/page.asp%3Fxsl%3Dtavole%26xml%3Dmicroscopica.fusto%26tavola%3DPerixilematici&usg=__MrTYG99m8k
AMRTi9ok3XVGIGuw=&h=400&w=500&sz=80&hl=it&start=13&um=1&tbnid=IQiiyiHdJoeFmM:&tbnh=104&tbnw=130&prev=/images%3
Fq%3Dtessuti%2Bvegetali%2Bfloema%2Bxilema%26hl%3Dit%26cr%3DcountryIT%26um%3D1
Meristematici
Le piante sono degli organismi a crescita indefinita che, a differenza degli animali,
continuano a crescere per tutta la loro vita. Ciò è possibile perchè possiedono dei tessuti
le cui cellule si possono dividere indefinitamente: sono i meristemi. Le cellule derivate
dai meristemi, innanzi tutto crescono di dimensioni (crescita per distensione) e poi si
differenziano, acquisendo forme e funzioni diverse. I tessuti meristematici si possono
dividere in primari e secondari.
I meristemi primari si trovano all'apice dei fusti e delle radici di tutte le piante;
l'allungamento di questi organi dipende proprio dalla produzione ordinata di nuove
cellule nei meristemi apicali e dalla crescita per distensione delle cellule prodotte. Le
cellule dei meristemi primari sono piccole, isodiametriche, con parete sottile; il nucleo
occupa buona parte del volume cellulare e il citoplasma è denso con vacuoli molto
piccoli, numerosissimi ribosomi, ed un corredo completo di organelli (proplastidi,
mitocondri,..).
I meristemi secondari sono localizzati nelle parti legnose dei fusti e delle radici e
quindi si trovano solo nelle Gimnosperme e nelle Dicotiledoni legnose. Essi sono il
cambio cribrovascolare e il cambio subero-fellodermico.
Il primo, più interno, produce i tessuti di conduzione, legno e libro, indispensabili per la
crescita della pianta ed è responsabile dell'aumento in spessore, l'accrescimento
secondario, dei fusti e delle radici.
Il secondo, esterno, produce il sughero, il tessuto di protezione che si sostituisce
all'epidermide nelle zone in struttura secondaria.
Questi due meristemi si definiscono "secondari" perchè derivano, per la maggior parte,
da cellule adulte che hanno riacquistato la capacità di dividersi, e che sono organizzate
in modo diverso rispetto a quelle dei meristemi primari, conservando alcuni caratteri
tipici delle cellule adulte: molte sono allungate e contengono dei grossi vacuoli.
Parenchimatici
Sono sempre costituiti da cellule vive con pareti permeabili e citoplasma organizzato in
modi differenti in relazione alle funzioni che le cellule svolgono: in molti casi (ad es.
nel parenchima fondamentale, clorofilliano, a riserva d'acqua) vi è un vacuolo centrale
che preme il citoplasma e il nucleo contro la parete cellulare, in altri casi, soprattutto in
alcuni parenchimi con funzione di riserva, le sostanze accumulate sono così abbondanti
da riempire completamente la cellula; in questo caso il vacuolo è quasi completamente
assente.
I tipi principali di parenchima sono:
1. Parenchima fondamentale: è costituito da cellule con forma poliedrica e parete
sottile. Il volume cellulare è occupato per la maggior parte da un grosso vacuolo e il
citoplasma è ridotto ad un sottile strato contro la parete cellulare. Si trova in diverse
parti della pianta, ad es. nella corteccia primaria del fusto e delle radici.
2. Parenchima di riserva: può accumulare diversi tipi di sostanze nel citoplasma:
zuccheri e proteine nei vacuoli, amido nei plastidi, oli ecc. Raramente le sostanze di
riserva vengono depositate all'esterno della membrana plasmatica: è il caso delle
emicellulose nell'endosperma di alcuni semi (es. palma da dattero e caffè) che formano
uno strato all'interno della parete cellulare.
3. Parenchima fotosintetico (clorenchima): è costituito da cellule ricche di cloroplasti,
organelli contenenti diversi tipi di pigmenti, tra cui importantissima la clorofilla, di
colore verde. In essi avviene la fotosintesi, processo in cui la luce fornisce l'energia
necessaria per trasformare le molecole di anidride carbonica in zuccheri. Il parenchima
fotosintetico si trova in tutte le parti verdi della pianta ma è particolamente specializzato
nella foglia.
4. Parenchima aerifero (aerenchima): è molto diffuso nelle radici e nelle piante
acquatiche, tra le cellule vi sono ampi spazi intercellulari che permettono la circolazione
dei gas.
Inoltre le cellule parenchimatiche possono contenere nel vacuolo sali di acidi organici
(ossalato di calcio), tannini, ecc.
Tegumentali
Rivestono tutta la pianta ed hanno funzione di protezione. I tessuti tegumentali che
rivestono le parti aeree hanno, come funzione principale quella di impedire una
traspirazione troppo elevata, che provocherebbe l'avvizzimento della pianta, perciò sono
costituiti, per la maggior parte, da cellule a stretto contatto tra loro, senza spazi
intercellulari, e hanno le pareti impregnate da polimeri impermeabili: la cutina o la
suberina. Il tessuto tegumentale che riveste le parti non legnose della pianta (fusti
giovani, foglie, frutti, fiori) è l'epidermide, mentre le parti lignificate sono rivestite dal
sughero.
Anche la radice è rivestita, nelle parti non lignificate, da un'epidermide, il rizoderma,
che ha una funzione completamente diversa: quella di assorbire acqua e sali minerali dal
terreno: anche qui le cellule sono strettamente a contatto tra di loro ma hanno parete
molto permeabile e alcune cellule possiedono delle lunghe estroflessioni, i peli radicali,
che servono per aumentarne la superficie assorbente.
Epidermide
Riveste le foglie, i fusti erbacei, i fiori e i frutti; le sue funzioni principali sono il
controllo del movimento dell'acqua tra interno ed esterno della pianta, la protezione
contro altri organismi (es. funghi, batteri ed insetti) e contro agenti non biologici (es.
vento). E' costituita da tre tipi fondamentali di cellule:
1. Cellule ordinarie. Sono cellule vive, strettamente a contatto tra loro in modo da
formare una pellicola impermeabile. Hanno forma variabile, da rettangolare, a
poligonale, lobata o frastagliata, che dipende dalla specie e dall'organo che rivestono.
Generalmente mancano di cloroplasti e sono trasparenti in modo da non ostacolare il
passaggio della luce verso il parenchima clorofilliano sottostante. Le loro pareti cellulari
esterne sono impermeabilizzate dalla deposizione di cutine. Tutte le cellule
dell'epidermide, sono, inoltre, ricoperte da una pellicola continua di cutina, la cuticola.
La cutina, oltre ad essere impermeabile, è in grado di riflettere le radiazioni solari e non
viene digerita da alcun organismo, quindi protegge la pianta da funghi e batteri.
All'esterno della cuticola si trovano sempre delle cere, sotto forma di cristalli di varia
forma, che accrescono l'impermeabilizzazione dell'epidermide. In alcuni casi questo
strato ceroso ha uno spessore notevole, basti pensare alla patina biancastra, la "pruina",
che riveste le prugne e gli acini d'uva!
2. Stomi. Lo strato impermeabile formato dalle cellule ordinarie è interrotto dagli stomi,
dei "buchi" ad apertura regolabile, attraverso i quali avvengono la traspirazione e gli
scambi di gas tra l'interno della pianta e l'ambiente esterno, due processi indispensabili
per la vita della pianta. Ogni stoma è costituito da due cellule di guardia, generalmente
reniformi, che delimitano il poro stomatico (o rima stomatica). Grazie allo spessore non
uniforme della parete cellulare e all'orientamento delle microfibrille di cellulosa, quando
le cellule di guardia assorbono acqua, si allungano e divaricano aprendo il poro
stomatico, viceversa quando perdono acqua lo richiudono. Segnali quali la luce e la
carenza di anidride carbonica nei tessuti fotosintetici danno il via ad un processo che si
conclude con l'apertura dello stoma, mentre il buio, la carenza d'acqua, il calore troppo
elevato ne determinano la chiusura.
3. Tricomi o peli. Comprendono tutte le strutture costituite da una o più cellule che si
proiettano al di fuori del piano dell'epidermide; possono avere funzione protettiva (peli
di rivestimento) oppure produrre sostanze di vario tipo: oli essenziali, sostanze urticanti,
enzimi proteolitici, e così via (peli ghiandolari).
Periderma
Le parti lignificate della pianta sono protette da un tessuto tegumentale secondario, il
periderma, che si forma in seguito all'attività del fellogeno o cambio suberofellodermico che produce
- verso l'esterno il sughero, privo di spazi intercellulari, costituito da cellule rettangolari
in sezione trasversale con parete cellulare suberificata, morte e contenenti aria a
maturità. Il sughero quindi è un tessuto impermeabile ed è anche un isolante termico.
- verso l'interno il felloderma, in quantità molto minore rispetto al sughero, costituito da
cellule parenchimatiche.
Meccanici
La rigidità dei tessuti vivi della pianta dipende dal turgore cellulare. Pensate ad una
foglia d'insalata o ad un rametto di basilico: in mancanza d'acqua avvizziscono perchè le
cellule perdono turgore e si afflosciano, come dei palloncini sgonfi. Questo tipo di
sostegno è sufficiente solo in organi molto piccoli e giovani, le piante perciò possiedono
dei tessuti specializzati nella funzione di sostegno e di protezione. Questi sono distinti
in due tipi principali: il collenchima e lo sclerenchima.
Collenchima
E' formato da cellule allungate e vive, con un grosso vacuolo centrale che preme il
citoplasma contro la parete cellulare. Le cellule collenchimatiche si riconoscono
facilmente in sezione perchè hanno una parete primaria in cui si alternano zone spesse,
che contengono grandi quantità di sostanze pectiche, e zone sottili.
Il collenchima è il tessuto di sostegno tipico delle parti giovani dei fusti e delle foglie
che si accrescono per distensione, infatti le sue cellule sono in grado di allungarsi
insieme ai tessuti circostanti e di sostenere l'organo in cui si trovano grazie alle pareti
cellulari ispessite e alla pressione di turgore che si genera al loro interno. Nei fusti
giovani il collenchima è in posizione periferica e può formare uno strato continuo al di
sotto dell'epidermide (es. sambuco) o dei cordoni longitudinali che formano costole
sporgenti: questo accade ad esempio nelle labiate (salvia, menta, ..) dove il fusto ha
sezione quadrangolare per la presenza di quattro cordoni sporgenti di collenchima.
I tipi principali sono:
- angolare
- lamellare.
Sclerenchima
Le sue cellule possiedono una parete secondaria, cioè uno strato di parete all'interno
della parete primaria, che frequentemente subisce un processo di lignificazione. La
presenza di lignina altera le caratteristiche della parete secondaria che diventa più
rigida, robusta e impermeabile.
Le pareti pectocellulosiche sono permeabili all'acqua e alle sostanze disciolte, mentre le
pareti lignificate sono impermeabili: quindi, affinchè le cellule con parete lignificata
possano scambiare tra loro acqua, nutrienti ed altre sostanze, alcune zone della parete
primaria non sono rivestite dalla parete secondaria e formano dei "canali", le
punteggiature, attraverso i quali avviene il passaggio di sostanze tra cellule adiacenti.
Al termine del processo di differenziamento, la maggior le cellule sclerenchimatiche
muore ed il protoplasma degenera.
I tipi principali sono:
- sclereidi = cellule brevi, a volte ramificate
- fibre = cellule lunghe, mai ramificate
Conduttori
Sono tessuti complessi, cioè costituiti da più tipi cellulari. Comprendono lo xilema (o
legno) e il floema (o libro, o tessuto cribroso).
Il primo trasporta a lunga distanza la linfa grezza, costituita principalmente da acqua e
soluti, dalle radici alle foglie, mentre il secondo trasporta la linfa elaborata, costituita da
soluzioni acquose di sostanze organiche (soprattutto zuccheri prodotti dalla fotosintesi,
amminoacidi ed ormoni) dalle foglie al resto della pianta.
Gli elementi cellulari in cui scorre la linfa grezza sono estremamente diversi da quelli in
cui scorre la linfa elaborata. Quelli dello xilema sono cellule morte prive di
protoplasma, con parete ispessita e, in parte, lignificata. Si possono quindi paragonare a
dei tubicini. In essi la linfa viene "risucchiata" verso l'alto dall'evaporazione che si
verifica nelle foglie (come una bibita in una cannuccia) e perciò è sotto tensione.
Gli elementi del floema, invece, sono cellule vive, anche se profondamente modificate,
il trasporto della linfa è, almeno in parte, attivo e la linfa è sotto pressione. Oltre agli
elementi di conduzione, nello xilema e nel floema si trovano: fibre, con funzione di
sostegno meccanico, e cellule parenchimatiche con funzione di riserva di acqua e di
sostanze nutritizie.
Xilema
Gli elementi di conduzione hanno parete primaria rinforzata all'interno da ispessimenti
di parete secondaria lignificata che impediscono agli elementi stessi di collassare a
causa della tensione interna.
A seconda del tipo di ispessimenti, vengono classificati in anulati, spiralati,
scalariformi, reticolati oppure punteggiati (quando la parete secondaria presenta aree
circolari non ispessite che prendono il nome di punteggiature areolate).
Inoltre, gli elementi di conduzione si distinguono in:
Tracheidi o vasi chiusi: sono cellule allungate e affusolate. La linfa, per passare da una
cellula all'altra, deve sempre attraversare la lamella mediana e le pareti primarie delle
cellule contigue.
Articoli dei vasi: sono impilati longitudinalmente a formare le trachee o vasi aperti.
Durante il differenziamento la parete cellulare tra articoli sovrapposti subisce un
processo di lisi e viene eliminata: si forma così una larga apertura, la perforazione, che
può essere semplice, se costituita da un unico foro, o composta se costituita da due o più
fori. All'interno di una trachea la linfa passa senza ostacoli da un articolo all'altro.
Il legno o xilema secondario è un tessuto molto complesso, in cui si distinguono
sempre il sistema longitudinale o assiale che ha funzione di conduzione della linfa
grezza e di sostegno, e il sistema dei raggi parenchimatici che provvede al trasporto
trasversale di sostanze nutritizie ed ha funzione di riserva di carboidrati e sostanze
minerali.
Il sistema longitudinale del legno delle Gimnosperme è molto diverso da quello delle
Angiosperme: nelle Gimnosperme è costituito da un unico tipo cellulare che ha sia
funzione di conduzione, sia di sostegno: le fibrotracheidi con punteggiature areolate
(legno omoxilo); nelle Dicotiledoni, invece, è formato da diversi tipi cellulari: elementi
vascolari, con funzione di conduzione, fibre, con funzione di sostegno e cellule
parenchimatiche con funzione di riserva (legno eteroxilo).
Anche i raggi midollari sono organizzati in modo differente nei due gruppi: nelle
Dicotiledoni hanno altezza e larghezza variabili, da una a numerose cellule, anche nella
stessa pianta, e sono costituite solo da cellule parenchimatiche, mentre nelle
Gimnosperme generalmente sono uniseriati e possono essere formati anche da
fibrotracheidi.
Nelle sezioni trasversali del legno, di solito si notano, anche ad occhio nudo, degli anelli
concentrici; questi si formano perchè il cambio cribrovascolare ha un'attività periodica:
è attivo nei periodi favorevoli alla crescita, mentre è quiescente se il clima è troppo
caldo, freddo o secco. Nelle sezioni microscopiche i singoli anelli sono sempre ben
distinguibili tra loro perchè gli ultimi strati di cellule formati prima della quiescenza
sono costituiti solo da fibre, e costituiscono il cosiddetto legno di chiusura.
Floema
Gli elementi di conduzione hanno una parete primaria che può essere ispessita ma mai
lignificata. Si tratta però di una parete particolare in quanto vi sono plasmodesmi molto
grandi: i pori cribrosi, raggruppati in aree cribrose.
A maturità sono privi di nucleo funzionante e il citoplasma è profondamente modificato.
Gli elementi di conduzione possono essere di due tipi:
Le cellule cribrose (tipiche delle Gimnosperme): sono allungate e affusolate, i pori
cribrosi hanno le stesse dimensioni e la stessa distribuzione in tutte le pareti cellulari.
Gli articoli dei tubi cribrosi (caratteristici delle Angiosperme): sono impilati
longitudinalmente a formare i tubi cribrosi. Gli articoli sovrapposti sono separati tra loro
dalle placche cribrose, attraversate da grandi pori con un sottile strato di callosio tra
parete e membrana plasmatica.
Le placche cribrose possono essere semplici, se possiedono una sola area cribrosa, o
composte. Gli elementi cribrosi sono associati alle cellule compagne, nelle
angiosperme, e alle cellule albuminose nelle gimnosperme, che controllano il
metabolismo degli elementi di conduzione ai quali sono associate e probabilmente sono
implicate nel trasporto di sostanze verso l'interno e verso l'esterno del floema.
Secretori
Sono organizzati in modo molto variabile, possono essere interni o esterni alla pianta,
comprendono singole cellule, canali o cavità, peli secernenti. Alcuni di essi, gli idatodi e
le ghiandole saline, riversano all'esterno acqua e sali minerali in eccesso; altri
sintetizzano ed accumulano o riversano all'esterno prodotti organici di vario tipo
(alcaloidi, oli essenziali, resine, gomme, latici, nettare, enzimi proteolitici, ecc. ), spesso
importanti per le relazioni tra piante e animali.
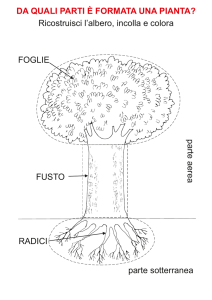
Fusto
Il fusto, oltre a collegare le radici con le foglie mediante i tessuti vascolari, é quella
parte della pianta che produce, sostiene e orienta nello spazio le foglie. Infatti é proprio
nella parte apicale del fusto, e precisamente nel meristema apicale, che si formano le
bozze fogliari, e quindi viene organizzata la distribuzione delle foglie sul ramo.
Nella maggior parte dei casi nei fusti si distinguono chiaramente nodi e internodi, i
nodi sono i punti in cui sono inserite le foglie, mentre gli internodi sono i segmenti
compresi tra due nodi successivi.
Se provate ad osservare un qualsiasi rametto in crescita, vi accorgerete che la lunghezza
degli internodi aumenta man mano che ci si allontana dall'apice: la zona in cui si
verifica questo allungamento é la zona di distensione. In essa le cellule, originate dal
meristema apicale, cessano di dividersi, aumentano di dimensioni e si differenziano,
cioé assumono gradualmente caratteristiche morfologiche e fisiologiche differenti, che
dipendono dalla funzione che avranno nel fusto differenziato.
Alla zona di distensione segue la zona di struttura primaria, costituita da tessuti
differenziati, e caratterizzata dalla presenza dei fasci cribrovascolari, fasci, cioé,
costituiti da floema e xilema.
Nelle piante erbacee (la maggior parte delle Monocotiledoni e alcune Dicotiledoni), una
volta che si é differenziata la struttura primaria, questa viene mantenuta per tutta la vita
della pianta. Nelle piante legnose (alcune Dicotiledoni e tutte le Gimnosperme) invece,
alla zona di struttura primaria segue quella di struttura secondaria, nella quale il
tessuto più abbondante é il legno, che si forma in seguito al differenziamento e
all'attività del cambio cribrovascolare.
Struttura Primaria
Il fusto in struttura primaria é costituito, dall'esterno verso l'interno, da tre parti:
epidermide, corteccia e cilindro centrale (o stele).
La corteccia é costituita prevalentemente da tessuto parenchimatico, le sue cellule più
superficiali, che vengono raggiunte dalla luce, contengono cloroplasti, infatti i fusti
giovani sono verdi e svolgono la fotosintesi. Nella corteccia si trovano dei tessuti
meccanici: il collenchima, in posizione più superficiale e lo sclerenchima, costituito
prevalentemente da fibre, in posizione più profonda. Entrambi possono circondare
completamente il fusto o formare fasci longitudinali.
Frequentemente non c'é un confine netto tra corteccia e il cilindro centrale, che occupa
la maggior parte dello spessore del fusto e contiene i fasci vascolari.
Osservando una sezione trasversale di un fusto primario é abbastanza facile distinguere
una Dicotiledone (o una Gimnosperma) da una Monocotiledone; infatti Dicotiledoni e
Gimnosperme hanno i fasci cribrovascolari disposti secondo un cerchio regolare
(secondo una eustele), mentre le Monocotiledoni hanno fasci sparsi in tutto lo spessore
della stele, sono cioé disposti secondo una atactostele. Un'ulteriore differenza tra i due
gruppi di piante sta nel tipo di fasci vascolari che sono collaterali aperti nelle
Dicotiledoni e Gimnosperme e collaterali chiusi nelle Monocotiledoni (la spiegazione
dei due termini si trova nelle didascalie alle figure).
Struttura Secondaria
Nelle Dicotiledoni legnose e nelle Gimnosperme lo strato di cellule procambiali dei
fasci vascolari riprende l'attività meristematica e dà origine al cambio intrafasciale;
contemporaneamente le cellule parenchimatiche poste tra i singoli fasci si
sdifferenziano e iniziano anch'esse a dividersi dando origine al cambio interfasciale. In
questo modo si forma un cambio cribrovascolare continuo, circolare, che inizia a
produrre il legno internamente e il floema secondario, in quantità minore, esternamente.
Poiché il cambio si forma tra lo xilema e il floema, il midollo e le porzioni xilematiche
dei fasci vascolari rimangono al centro del fusto in struttura secondaria.
Man mano che procede la crescita secondaria, il cambio viene spostato verso l'esterno
dal legno che esso stesso ha prodotto, quindi aumenta di circonferenza e tutti i tessuti al
suo esterno (libro secondario e tessuti di origine primaria: floema dei fasci vascolari,
parenchima corticale ed epidermide) vengono stirati e tendono a strapparsi. Il floema
non viene lacerato grazie alla formazione del parenchima di dilatazione: le cellule dei
raggi parenchimatici aumentano di dimensioni ed effettuano numerose divisioni
cellulari consentendo un graduale allargamento del raggio che assume forma a V in
sezione trasversale.
L'epidermide può assecondare l'aumento di circonferenza anche per alcuni anni, poiché
le cellule epidermiche possono riprendere a dividersi, ma prima o poi viene lacerata. Il
corpo secondario della pianta viene quindi protetto da un tessuto tegumentale
secondario, il periderma, che si forma in seguito all'attività del cambio suberofellodermico.
Foglia
Le funzioni principali della foglia sono la fotosintesi, e quindi la sintesi di sostanze
organiche a partire dall'anidride carbonica, e la traspirazione, essenziale per la risalita
della linfa grezza nei vasi del legno. Per questo motivo in genere le foglie sono laminari
e appiattite: in questo modo la loro superficie é molto elevata rispetto al volume, e una
superficie ampia favorisce sia gli scambi di gas con l'esterno, sia l'evaporazione
dell'acqua. Le foglie però possono essere spesse, cilindriche quando é essenziale per la
vita della pianta limitare la traspirazione: é il caso, ad esempio, delle foglie aghiformi
del pino e di altre conifere.
Le foglie originano da bozze fogliari che si formano nella parte basale dell'apice
meristematico del fusto (vedi istologia, tessuti meristematici), nelle prime fasi della
crescita tutte le cellule dell'abbozzo si dividono intensamente, successivamente le
divisioni proseguono in alcune zone ben determinate, che fanno si che la foglia assuma
la sua forma definitiva, infine le cellule crescono per distensione e la foglia raggiunge le
sue dimensioni definitive.
Le foglie sono rivestite dall'epidermide: si distinguono un'epidermide ventrale (o
adassiale) e una dorsale (o abassiale). Tra le due epidermidi vi é il mesofillo, costituito,
almeno in parte, da parenchima fotosintetico, in cui corrono le nervature, costituite dai
fasci vascolari avvolti da una guaina e, a volte, accompagnate da tessuti di sostegno.
Radice
Le funzioni principali delle radici sono l'assorbimento di acqua e sali minerali dal
terreno e l'ancoraggio della pianta al substrato. Anche nelle radici, come nei fusti, si
distingue una zona meristematica apicale, cui segue una zona di distensione e quindi di
struttura primaria. Questa é la zona della radice specializzata nell'assorbimento ed é
costituita da tre regioni:
- l'epidermide (o rizoderma) caratterizzata dalla presenza di peli radicali
- la corteccia di solito molto sviluppata, costituita da tessuto parenchimatico, che
frequentemente accumula granuli d'amido. Gli spazi intercellulari sono generalmente
più ampi che nel fusto e mancano i tessuti di sostegno. Lo strato più interno della
corteccia, a contatto con la stele, é l'endoderma. Questo é costituito da un unico strato
di cellule, prive di spazi intercellulari, con ispessimenti di parete nastriformi, le strie di
Caspary, nelle pareti radiali e trasversali. In corrispondenza di questi ispessimenti il
plasmalemma aderisce strettamente alla parete cellulare che è suberificata o lignificata,
e quindi idrofoba. L'endoderma é importante per il passaggio delle soluzioni dalla
corteccia alla stele: esse, infatti, vengono assorbite dall'epidermide e possono
attraversare la corteccia liberamente passando nelle pareti cellulari che sono permeabili,
tuttavia non possono attraversare l'endoderma, perché vengono bloccate dalle bande di
Caspary. Per entrare nella stele, i soluti devono necessariamente attraversare il
citoplasma delle cellule endodermiche, e quindi venire selezionati dalle membrane
cellulari.
- il cilindro centrale (o stele) è costituito da uno strato esterno, il periciclo,
parenchimatico e generalmente uniseriato la cui proliferazione dà origine alle radici
laterali, e dai tessuti conduttori immersi in un tessuto parenchimatico.
Nelle Dicotiledoni e Gimnosperme lo xilema e il floema formano una struttura raggiata
con arche xilematiche e arche floematiche alternate tra di loro, il loro numero é basso,
quasi sempre inferiore a 6. In alcuni casi le arche xilematiche confluiscono al centro
della stele, si ha allora una actinostele, in altri casi, invece, il centro della stele é
occupato dal midollo, parenchimatico. Anche nelle Monocotiledoni vi sono arche
xilematiche e floematiche alternate, però il loro numero é molto più elevato: sono
generalmente 15-30 ed é sempre presente midollo al centro della stele.
Struttura Primaria
Nelle Monocotiledoni e nelle Dicotiledoni erbacee, la radice non ha una crescita
secondaria e quindi mantiene la struttura primaria per tutta la sua lunghezza. In una
radice di questo tipo, all'apice radicale, alla zona di distensione e a quella di struttura
primaria con funzione assorbente, segue una zona di struttura primaria
"invecchiata" che ha soprattutto la funzione di trasportare la linfa grezza dalle parti più
giovani dell'apparato radicale verso il fusto, e la linfa elaborata in senso opposto. In
questa zona, anche se l'organizzazione dei tessuti resta invariata, le cellule subiscono
delle modificazioni:
1. I peli radicali degenerano e, al di sotto dell'epidermide, si può differenziare un
esoderma, cioé uno strato di cellule con pareti suberificate;
2. Le pareti delle cellule endodermiche vengono impermeabilizzate dalla deposizione di
una lamella di suberina, successivamente vengono ispessite per apposizione di uno
strato cellulosico e lignificate. In sezione trasversale le pareti di queste cellule hanno
ispessimenti ad "U" o ad "O". Solo alcune cellule endodermiche mantengono la struttura
con le strie di Caspary e costituiscono i punti di permeazione.
3. Le cellule degli strati esterni della corteccia e il parenchima del cilindro centrale in
cui sono immersi i cordoni vascolari possono subire un processo di ispessimento e
lignificazione della parete cellulare. Tutte queste modificazioni servono soprattutto per
bloccare il rilascio verso il terreno delle soluzioni trasportate dai sistemi vascolari.
Struttura Secondaria
Nelle Dicotiledoni legnose e nelle Gimnosperme il cambio cribrovascolare origina
dalle cellule parenchimatiche poste tra le arche xilematiche e quelle floematiche, e da
quelle del periciclo esterne alle arche xilematiche, che si sdifferenziano e riprendono a
dividersi.
All'inizio il cambio ha quindi forma ondulata ma in corrispondenza delle arche
floematiche produce il legno più velocemente che nelle altre zone e viene spinto verso
l'esterno. In questo modo assume rapidamente una forma circolare e inizia a produrre
legno secondario verso l'interno e floema secondario verso l'esterno. Quando
l'accrescimento secondario prosegue per un certo tempo, la struttura della radice diventa
simile a quella del fusto secondario, e a volte non è semplice distinguere i due organi tra
loro.
Riproduzione
Nelle Spermatofite (Gimnosperme e Angiosperme) e nelle altre piante terrestri, la
riproduzione sessuale é legata ad un ciclo vitale aplodiplonte in cui si alternano una
generazione aploide (il gametofito) e una diploide (lo sporofito). Il ciclo si può
schematizzare in questo modo:
- i gametofiti (aploidi) producono i gameti (aploidi),
- due gameti si fondono dando origine allo zigote (diploide),
- dallo zigote si forma lo sporofito (per divisioni mitotiche),
- nelle strutture riproduttive dello sporofito (diploide) si formano le spore aploidi (per
meiosi)
- le spore danno origine ai gametofiti (per mitosi), che producono i gameti ... e il ciclo
si chiude.
..... ma che cosa sono in concreto i gametofiti e gli sporofiti? Nelle Angiosperme e nelle
Gimnosperme gli sporofiti sono le "piante" formate da radici, fusti e foglie; i
gametofiti sono invece molto piccoli, microscopici: i gametofiti maschili sono i granuli
di polline germinati, quelli femminili sono racchiusi negli ovuli, all'interno delle
strutture riproduttive femminili (i coni femminili nelle Gimnosperme, i carpelli dei fiori
nelle Angiosperme).
Le Spermatofite sono le uniche piante in cui la riproduzione sessuale ha come risultato
la produzione del seme ("Spermatofita" significa appunto "pianta che produce semi"),
costituito da una piantina in miniatura, quasi completamente disidrata e quiescente,
l'embrione, provvisto delle riserve nutritive necessarie per la germinazione, è protetto
dai tegumenti.
L'"invenzione" del seme ha avuto una grandissima importanza nella diffusione delle
Spermatofite in ogni ambiente della Terra; il seme, infatti, ha la funzione di propagare la
specie sia nello spazio, grazie a diversi meccanismi che ne facilitano la dispersione (ali,
peli, uncini, ecc.), sia nel tempo, dato che può rimanere vitale per lunghi periodi, anche
in condizioni ambientali avverse, grazie allo stato di latenza in cui si trova l'embrione.
Gimnosperme
La riproduzione sessuale nelle Gimnosperme é un processo con caratteristiche diverse
nei diversi gruppi. In questo atlante viene descritta, come esempio, la riproduzione
sessuale delle Conifere. Si tratta di un processo complesso, che in alcune specie richiede
circa due anni per completarsi. Nel pino, e nella maggior parte delle altre conifere, gli
apparati riproduttivi sono i coni (o strobili), sempre unisessuali, generalmente portati
dalla stessa pianta: negli strobili maschili si differenziano i granuli di polline, in quelli
femminili gli ovuli contenenti le cellule uovo (i gameti femminili).
I granuli di polline vengono prodotti in grandissima quantità, trasportati dal vento e
catturati da sostanze vischiose prodotte dagli ovuli, quindi penetrano al loro interno. La
fecondazione non avviene immediatamente, in questo stadio infatti negli ovuli non é
ancora avvenuta la divisione meiotica che porta alla formazione delle spore femminili
(le megaspore); solo circa un mese dopo l'impollinazione, a partire da una cellula
madre vengono prodotte quattro megaspore di cui una darà origine al gametofito
femminile, l'endosperma primario, il cui sviluppo richiede più di un anno. Nel
frattempo i granuli pollinici germinano e producono il tubetto pollinico che si accresce
nei tessuti dell'endosperma portando i gameti maschili (i nuclei spermatici) fino alla
cellula uovo. Avviene la fecondazione, si forma lo zigote che dà origine all'embrione,
che nelle conifere possiede numerosi cotiledoni (o foglie embrionali); l'endosperma
funziona come tessuto nutritivo, il tegumento diventa duro e si trasforma nel
rivestimento del seme. Contemporaneamente lo strobilo femminile si accresce, diventa
legnoso e si trasforma nella pigna. A maturità le squame della pigna si divaricano e
viene liberato il seme, spesso provvisto di un'ala che ne facilita la dispersione.
Angiosperme
La principale caratteristica che distingue le Angiosperme dalle Gimnosperme é la
produzione del fiore che é costituito da elementi sterili (i sepali e i petali) e da elementi
fertili, normalmente presenti nello stesso fiore: gli stami in cui si differenziano i granuli
di polline, e il carpello (o i carpelli): foglie modificate ripiegate su se stesse, in cui si
distinguono, dall'apice verso la base, lo stimma, lo stilo e l'ovario, che racchiude gli
ovuli. Quindi, nelle Angiosperme, gli ovuli non sono semplicemente "portati" da
squame, come accade nelle Gimnosperme, ma sono racchiusi all'interno dell'ovario che,
durante la maturazione del seme, si trasforma nel pericarpo del frutto. Questa é una
importante differenza rispetto alle Gimnosperme (le piante "a semi nudi"): infatti il
frutto é un organo esclusivo delle Angiosperme, molto importante ai fini della
dispersione dei semi ad opera del vento, dell'acqua o di animali.
Il processo riproduttivo delle Angiosperme é simile a quello delle Gimnosperme, con
alcune differenze:
- il granulo pollinico completamente germinato (il gametofito maschile) contiene solo
tre nuclei anziché quattro come nelle Gimnosperme, una cellula vegetativa e due cellule
spermatiche;
- il polline non viene disperso solamente dal vento, ma alla sua dispersione partecipano
numerosi animali, tra cui soprattutto gli insetti;
- il polline non entra mai all'interno dell'ovulo ma aderisce alla superficie dello stimma,
quindi, in alcune specie, il tubetto pollinico deve allungarsi in modo considerevole
attraverso lo stilo per giungere agli ovuli;
- il gametofito femminile maturo, il sacco embrionale, é costituito solamente da 8
nuclei, e contiene un'unica cellula uovo che non é racchiusa nell'archegonio;
- avviene il processo della doppia fecondazione: alla fecondazione partecipano entrambi
i nuclei spermatici del granulo pollinico: uno feconda la cellula uovo dando origine allo
zigote, l'altro invece si fonde con due nuclei (i nuclei polari) che si trovano al centro del
sacco embrionale, e si forma l'endosperma triploide che servirà come tessuto di riserva
per lo sviluppo dell'embrione;
- gli embrioni delle Angiosperme possono avere uno o due cotiledoni (foglie
embrionali), rispettivavente nelle Mono- e nelle Dicotiledoni, che possono funzionare
come organi di riserva o come austori durante la germinazione dell'embrione.