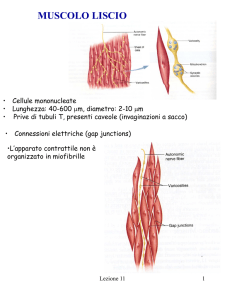
Muscolo liscio
• Arterie
• nell’intestino
• nei bronchioli
• nella vescica
• nell’utero per esempio.
Esso non ha striature poiché manca dei
sarcomeri.
Scorrimento dei filamenti nel
muscolo liscio
I filamenti di actina e di
miosina del muscolo liscio
sono più lunghi di quelli dei
muscoli scheletrico e
cardiaco. Un’altra differenza
tra il muscolo liscio e i
muscoli striati sta nel fatto
che i filamenti di miosina
possiedono teste su tutta la
loro lunghezza.
Nel muscolo liscio la miosina
può scorrere sui filamenti di
actina per un spazio più lungo
perché manca la limitazione
dovuta alla lunghezza del
sarcomero. Di conseguenza i
muscoli lisci possono
accorciarsi di più di quelli
scheletrici.
Classificazione dei recettori
per le catecolamine
Tipo di recettore Bersaglio
Effetto
Agonista
1
Musc. liscio
arterie e vene
Vasocostrizione
Noradrenalina
2
Circolo cutaneo
Vasocostrizione
Noradrenalina
1
Nodo SA
freq. cardiaca
Sist. cond. card. veloc. conduz.
Miocardio di
forza contraz.
lavoro
Noradr. e adren.
Noradr. e adren.
Noradr. e adren.
2
Musc. liscio
arterie e vene
Adrenalina
Vasodilatazione
Due meccanismi di accoppiamento eccitazione-contrazione
Nel primo meccanismo chiamato accoppiamento farmacomeccanico, un farmaco, un neurotrasmettitore o un ormone si lega
ai suoi recettori sulla membrana superficiale (vedi figura sopra) e
attiva la fosfolipasi C
Nel secondo meccanismo, che dipende dalla depolarizzazione della cellula
muscolare liscia, il calcio entra nella cellula dal liquido extracellulare. In alcune
cellule, un farmaco è in grado di aprire i canali del calcio recettore-attivati (vedi
figura a sinistra). In altre cellule, i canali del calcio voltaggio attivati si aprono in
risposta alla depolarizzazione della cellula (vedi figura a destra). In entrambi i
casi, l’ingresso di calcio attraverso questi canali depolarizza al cellula. L’aumento
del calcio intracellulare può attivare la contrazione direttamente ed indurre il
reticolo sarcoplasmatico a rilasciare più calcio, come nel muscolo cardiaco.
Il muscolo liscio può essere diviso in due tipologie principali: multiunitario e unitario (viscerale).
Il muscolo liscio multiunitario ha unità motorie che somigliano a quelle del muscolo scheletrico. La
contrazione del muscolo liscio multiunitario è sotto il controllo nervoso e normalmente non si osservano
contrazioni spontanee. Queste cellule non sono connesse elettricamente, così le contrazioni sono
localizzate. Ci sono solo pochi muscoli lisci multiunitari nei mammiferi, esempi sono i corpi ciliari
(muscoli oculari utilizzati nella messa a fuoco), i vasi deferenti e i muscoli piloerettori che intervengono
nell’erezione pilifera.
Muscolo liscio unitario: muscoli fasici e muscoli tonici
I muscoli lisci unitari (anche noti come muscoli lisci viscerali) sono più presenti rispetto ai muscoli lisci
multiunitari. Esempi di muscoli lisci unitari si trovano nelle pareti dei vasi sanguigni e in organi cavi
come intestino, utero e gli ureteri.
Diversamente dal muscolo liscio multiunitario, le cellule del muscolo liscio unitario sono connesse
elettricamente via giunzioni comunicanti a bassa resistenza. Questo permette che l’eccitazione di una
cellula diffonda alle vicine e quindi che il tessuto operi come una unità funzionale. In questa proprietà è
simile al muscolo cardiaco.
Il muscolo liscio unitario può essere successivamente classificato in muscoli lisci fasici e tonici. I
muscoli lisci fasici, come quelli delle pareti dell’intestino mostrano un comportamento contrattile ritmico
(fasico). Queste contrazioni fasiche sono associate con potenziali d’azione prodotti da cambiamenti
rigenerativi nella permeabilità di canali del calcio voltaggio sensibili. Così queste cellule mostrano
potenziali d’azione dovuti al calcio piuttosto che al sodio come nel caso del nervo e del muscolo. I
muscoli lisci tonici hanno invece una contrazione continua (tono) e si trovano nelle arterie e negli
sfinteri.
I muscoli lisci tonici sono ineccitabili elettricamente e le contrazioni toniche non sono associate alla
generazione di potenziali d’azione .
I filamenti sono inseriti su zone di attacco sulla superficie della membrana o su corpi densi che
sono analoghi alle linee Z del muscolo striato. Il contenuto di actina e tropomiosina è circa doppio
rispetto al muscolo scheletrico, ma il contenuto di miosina è 4-5 volte meno rispetto al muscolo
scheletrico. Nondimeno, il muscolo liscio può sviluppare una forza (normalizzata per la sezione
trasversale) circa doppia rispetto al muscolo scheletrico.
Le cellule muscolari lisce sono generalmente più piccole (da 5 a 50 micrometri in diametro) delle
cellule muscolari scheletriche (da 10 a 100 micrometri di diametro). Non hanno i tubuli trasversi. Il
reticolo sarcoplasmatico è meno sviluppato rispetto al muscolo cardiaco o allo scheletrico.
Muscolo liscio unitario: contrazioni spontanee (cellule pacemaker che generano potenziali
pacemaker) o attivate da stiramento
Alcuni muscoli lisci unitari mostrano contrazioni spontanee che sono indipendenti dal controllo
nervoso. Queste contrazioni sono associate a potenziali d’azione generati da attività elettrica
spontanea (pace-making), come nelle regioni degli ureteri. Alcuni muscoli lisci unitari possono
essere attivati dallo stiramento (figura). Lo stiramento causa una depolarizzazione di queste
cellule muscolari lisce che porta alla generazione di potenziali d’azione, che causano contrazione.
Così questi muscoli lisci resistono attivamente allo stiramento o alla distensione (per esempio i
muscoli delle pareti intestinali o della vescica).
La maggiorparte dei muscoli lisci unitari sono sensibili a stimoli la
noradrenalina, adrenalina, l’istamina o gli ormoni. Un esempio relativo agli
ormoni è l’utero che contiene muscoli lisci sensibili ad estrogeni e al
progesterone.
I muscoli lisci unitari non rispondono in modo identico agli stessi agenti
chimici. Piuttosto la risposta dipende dal tipo di recettore di membrana
con cui questi agenti interagiscono e da come questi recettori, a loro volta,
influiscono sulla cellula.
Per esempio, i muscoli lisci intestinali sono innervati sia da fibre nervose
adrenergiche che colinergiche (vedi figura dopo). La stimolazione delle
fibre nervose colinergiche rilascia acetilcolina dalle varicosità che causa
aumento di forza contrattile. La stimolazione delle fibre nervose
adrenergiche rilascia noradrenalina, che induce il muscolo liscio a
rilassarsi.
D'altronde, il muscolo liscio di molti vasi sanguigni è sensibile alla
noradrenalina che deriva dai nervi adrenergici o che circola nel sangue
(rilasciata dalla ghiandola surrene). In questi muscoli lisci, comunque, la
norepinefrina induce un incremento della contrazione dei vasi.
rilasciamento
contrazione
Miosina fosfatasi - Attivata da adrenalina
Recettore per epinefrina
Protein kinasi inattiva
ATP cAMP
Protein kinasi attiva
Miosina fosfatasi
inattiva
Miosina fosfatasi
attiva
rilasciamento
Muscolo Liscio: velocità e durata della
contrazione. La maggior parte della muscolatura liscia si
trova nella parete degli organi cavi. La velocità di contrazione nel
muscolo liscio è molto più bassa che non nei muscoli scheletrico e
cardiaco. A differenza di questi ultimi però
la contrazione può
essere sostenuta
per tempi molto più
lunghi. La tensione
muscolare
mantenuta
costantemente dal
muscolo liscio a
livello misurabile
viene detta tono.
Il risultato di una tale relazione tensione attiva-lunghezza piatta è la risultante di una
forza attiva meglio mantenuta a qualsiasi lunghezza relativa.
La forza attiva nel muscolo liscio, normalizzata per l’area trasversale, è circa doppia
rispetto a quella del muscolo scheletrico, anche se il muscolo liscio contiene quattro
cinque volte meno miosina. Non è chiaro perché i ponti trasversali sono in grado di
sviluppare parecchia forza.
La relazione carico-velocità del muscolo liscio è simile nella forma a quello del
muscolo striato (figura). La massima velocità di accorciamento è dalle 300 alle
500 volte più lenta del muscolo scheletrico a rapida contrazione, in accordo
alla sua bassa velocità di idrolisi dell’ATP della miosina.
Muscolo cardiaco
Le cellule cardiache sono
elettricamente accoppiate tra loro
attraverso dischi intercalati che
contengono regioni a bassa
resistenza chiamate giunzioni
comunicanti. Sincizio elettrico
Giunzioni cellulari
Le cellule cardiache si collegano le
une alle altre tramite giunzioni,
regioni specializzate dette dischi
intercalari.
Sono
membrane
interdigitate
collegate da due diversi tipi di
giunzioni:
• desmosomi che trasferiscono la
tensione generata in una cellula a
quelle adiacenti
• gap junctions che permettono il
passaggio di cariche
Si possono identificare i miofilamenti, le caratteristiche bande A ed I e le
linee M e Z. I miofilamenti cardiaci sono della stessa lunghezza dei
miofilamenti scheletrici, cosicché la relazione forza attiva lunghezza è la
stessa del muscolo scheletrico.
I tubuli trasversi si trovano anche nel cuore di mammifero, ma non nella
stessa quantità nell’atrio e nel ventricolo. Come nel muscolo scheletrico, il
reticolo sarcoplasmatico forma delle strette connessioni con le membrane
del tubulo a T ma, contrariamente a quello che succede nel muscolo
scheletrico, il reticolo sarcoplasmatico forma tali connessioni (dette
connessioni superficiali) anche con la superficie sarcolemmale.
Le cellule muscolari cardiache hanno un diametro tra i 20-30 micrometri
La conduzione elettrica nel tessuto
cardiaco
Potenziali nei muscoli scheletrico e cardiaco
Il muscolo cardiaco non può essere tetanizzato. Nel muscolo cardiaco
comunque, il potenziale d’azione è abbastanza lungo rispetto alla scossa.
Così la membrana può essere stimolata ancora una volta solo dopo che la
forza della scossa semplice è decaduta quasi del tutto.
il fattore di amplificazione
del rilascio di calcio da
calcio indotto varia a
seconda della specie
La relazione forza-velocità del muscolo cardiaco è simile a quello del
muscolo scheletrico, eccetto che la velocità massima di accorciamento è
più bassa (figura). Questo è in accordo con una velocità più bassa della
ATPase della miosina cardiaca e con la più lenta velocità dei cicli dei ponti
trasversali.
Gli elementi passivi del tessuto connettivo nel muscolo cardiaco,
comunque, sono più rigidi e la relazione tra la lunghezza e la forza passiva
è spostata a sinistra rispetto al picco della curva lunghezza-forza attiva
Il muscolo cardiaco dipende pesantemente dal
metabolismo aerobico e pertanto contiene molti
mitocondri e molta mioglobina. La mioglobina
conferisce al cuore il suo caratteristico colore rosso.
Come studiato in precedenza, il metabolismo
aerobico è più efficiente della glicolisi nel produrre
ATP. I problemi sorgono quando
l’approviggionamento di ossigeno è scarso. Questo
ed altre difficoltà insorgono quando il flusso di
sangue che irrora una regione del cuore è scarso o
bloccato completamente. La contrazione si
indebolisce rapidamente o si ferma del tutto, e se il
flusso del sangue è ristretto per abbastanza tempo,
le cellule del miocardio moriranno.
La conduzione elettrica nel tessuto
cardiaco
Tessuti del cuore
Il ruolo del Ca2+ nel PDA delle cellule pacemaker
Classificazione dei recettori
per le catecolamine
Tipo di recettore Bersaglio
Effetto
Agonista
1
Musc. liscio
arterie e vene
Vasocostrizione
Noradrenalina
2
Circolo cutaneo
Vasocostrizione
Noradrenalina
1
Nodo SA
freq. cardiaca
Sist. cond. card. veloc. conduz.
Miocardio di
forza contraz.
lavoro
Noradr. e adren.
Noradr. e adren.
Noradr. e adren.
2
Musc. liscio
arterie e vene
Adrenalina
Vasodilatazione
Influenza del SN sulla frequenza
cardiaca
La stimolazione simpatica (noradrenalina) e l’adrenalina circolante
depolarizzano le cellule pacemaker aumentando la velocità di depolarizzazione
(incremento del flusso di ioni attraverso i canali If e per il Ca2+).
Raggiungendo più rapidamente la soglia aumenta la frequenza (azione
cronotropa positiva).
La stimolazione parasimpatica (Ach) iperpolarizza le cellule pacemaker e
rallenta la depolarizzazione (aumento della permeabilità per il K+ e diminuzione
per il Ca2+).
Raggiungendo più lentamente la soglia rallenta la frequenza (azione cronotropa
negativa).