―Agli stomi è stato affidato il compito di fornire il")
Stomi
Principale strumento
di controllo della traspirazione
RASCHKE (1976)
―Agli stomi è stato affidato il compito di
fornire il cibo mentre impediscono la sete‖
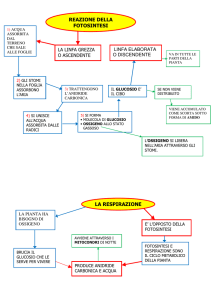
Fotosintesi-traspirazione
“Paradosso” o “Compromesso”
1
Equazione
per la fotosintesi
CO2 + H2O
(CH2O) n + O2
Questa equazione suggerisce:
•Gas, come CO2 e O2, sono importanti nel metabolismo energetico della pianta;
• Le piante devono scambiare i gas con l’ambiente; e
• Per ottenere CO2 devono necessariamente perdere H2O
(traspirare). Quindi, la traspirazione è un “danno” necessario della fotosintesi.
Considerazioni teoriche:
•E’ necessaria una grande area superficiale per un efficiente scambio gassoso;
• Una grande area superficiale offre una grande superficie di disidratazione;
•Ridurre la disidratazione vuol dire porre all’interno dell’organismo la superficie di
assorbimento e quindi la superficie di scambio gassoso;
• Traspirazione/Fotosintesi - Paradosso/Compromesso:
Paradosso: Per ottenere CO2 per la fotosintesi, le piante devono evolvere larghe
e sottili superfici e proteggerle dalla disidratazione
Compromesso: tra la quantità di CO2 assorbita e la quantità di H2O persa per
2
traspirazione
Complicazioni:
•La tendenza di perdere H2O è più grande della tendenza della CO2 a diffondere
nella pianta
Traspiration ratio = mol water transpired /mol CO 2 fixed
Questo rapporto è uguale a 200 cioè circa 200 Kg di H2O traspirata / 1Kg di
biomassa prodotta
Perché?
• Diffusione = gradiente / resistenza
A parità di resistenza per CO2 e H2O, il gradiente per queste molecole non è lo
stesso, il gradiente CO2 est/int è circa 0.36 mmol mol -1 quello per l’H2O è 32
•Diffusione e peso molecolare
Basandosi sul peso molecolare, la tendenza dell’H2O di diffondere fuori dalla foglia
è 1.56 volte più grande della CO2 a diffondere all’interno della foglia
Compromesso regolato dagli stomi
Essi devono quindi : Garantire una via
aperta per l’assorbimento di CO2 e
preservare la pianta da una perdita
eccessiva di H2O
3
Caratteristiche
speciespecifiche
• Numero
•Disposizione
•Taglia
•Forma
•Mobilità
Stomata typical of dicots
Stomata typical of monocots
Potato (Solanum)
Maize (Zea)
4
begonia
banana
5
Adattamenti strutturali per
ridurre la traspirazione
Cripta stomatica
Peli radicali
Quercus sp
Cycas sp
Oleander
6
cripta stomatica
Banksia sp
7
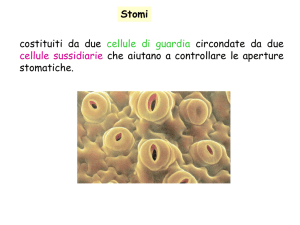
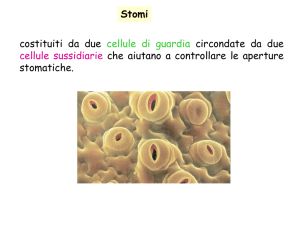
•Due cellule di guardia
Complesso stomatico
• Rima stomatica
•Cellule sussidiarie
Cellule di guardia
Presenti nelle foglie di tutte le piante vascolari e negli
organi di piante più primitive (muschi, epatiche)
A manubrio di ginnasta con
parti terminali bulbose
Graminacee e poche
monocotiledoni (palme)
diversità
morfologica
Reniformi
Dicotiledoni e altre
monocotiledoni
8
Le cellule di guardia sono cellule affiancate
e senza connessioni citoplasmatiche
Cellule epidermiche specializzate
che circondano la rima stomatica
Caratteristiche
delle cellule di
guardia
•piccoli cloroplasti
•Tracce di Rubisco
Scarsa fissazione della CO2
La fotosintesi potrebbe costituire
un sensore per la CO2
Sono strutturate in modo da poter
determinare con il loro
rigonfiamento e svuotamento
apertura e chiusura dello stoma
Mezzo principale di regolazione
degli scambi gassosi
“ valvole di turgore‖
9
Porzioni di pareti
differenzialmente
ispessite (5 µm)
Caratteristiche
strutturali della
parete delle cellule
di guardia
Micellazione radiale
delle microfibrille
di cellulosa
Maggiore ispessimento in
quella parte di parete che
delimita la rima stomatica
Permette alla cellula di
guardia, assorbendo H2O ed
espandendosi, di accrescersi in
lunghezza e non in diametro
Sembra più importante
10
dell’ispessimento delle pareti
Rima stomatica
Cellule sussidiarie
o compagne
È rappresentata da una lunga fenditura
che separa le due porzioni mediane delle
cellule di guardia
Cellule epidermiche differenziate
Aiutano le cellule di
guardia a controllare le
aperture stomatiche
Sono spesso assenti in
specie con stomi
reniforme e sostituite da
semplici cellule
epidermiche
11
Stomi: struttura ideale per regolare gli scambi gassosi
•Molti sono i pori sulla superficie delle piante;
•I pori comprendono una larga area della superficie di una pianta;
•I pori sono piccoli;
•I pori sono perfettamente distribuiti;
•I pori sono perfettamente localizzati;
•Il grado di apertura/chiusura dei pori è strettamente regolato in risposta a
condizioni ambientali.
12
2000-6000 stomi per cm2 = 2-3 % della superficie fogliare
Localizzati spesso solo nella pagina inferiore della foglia ma in molte specie
anche su ambedue le pagine
Da Meidner e
Mansfield, 1968
13
Diffusion from a "Standard Leaf"
Supplied
by Dr. Paul
Birnberg.
Quanto più piccolo è il poro, tanto più alta è l’efficienza
di diffusione per unità di superficie
14
Le molecole di gas diffondono
attraverso il poro formando
Conchiglie di diffusione
Vapore d’H2O diffonde attraverso gli
stomi in tutte le direzioni andando ad
occupare superfici non munite di stomi
Il paradosso dei
pori
Questo mantiene sempre ripido
il gradiente di pressione
di vapore fra l’interno e
l’esterno della foglia
15
Ha lo strato di
diffusione più breve
Il gradiente di
concentrazione più
ripido
La conduttività dipende:
•Dalla distanza che le molecole di H2O devono
compiere per diffondere (strato di diffusione)
•Dall’aria di scambio (superficie poro)
Se la distanza fra i pori è inferiore allo spessore dei loro strati
di diffusione, le emisfere centrate sui pori si sovrapporranno
e formeranno uno strato di diffusione unico.
16
Schema di apertura stomatica (1)
Stomi chiusi
(cellula di guardia flaccide)
Ingresso di H2O dalle cellule sussidiarie
Incremento della pressione
Stomi aperti
(cellula di guardia turgide)
17
Cellule di guardia, Ψ1= -3,5 MPa
Cellule sussidiarie, Ψ2= -1,5 MPa
1 2 3,5 1,5 2,0MPa
diminuzione del Ψw
nelle cellule di guardia dovuta ad una
diminuzione del Ψs
Stoma chiuso
Ψs
-1,9 MPa
Stoma aperto
Ψs
-3,5 MPa
Richiamo d’H2O
nelle cellule di guardia
dalle cellule sussidiarie
adiacenti
18
Aumenta pressione di turgore Ψp
e compensa il minore Ψs
Stoma aperto 3-12 µm di larghezza e 10-40 µm di lunghezza
Aumento del volume da 40 a 100 % a seconda della specie
l’incremento di volume nelle cellule di guardia
è dovuto all’aumento della concentrazione di soluti nel succo cellulare
Schema di apertura stomatica (2)
Stomi chiusi
Aggiunta di soluti
Riduzione del Ψs
(cellula di guardia flaccide)
Riduzione
del Ψw
Ingresso
di H2O
Incremento
della pressione
Stomi aperti
(cellula di guardia
turgide)
19
Quali soluti e
da dove vengono ?
Ipotesi amido-zucchero (1908)
La quantità di amido diminuiva durante l’apertura stomatica, la sua idrolisi in zuccheri
solubili erano responsabili della diminuzione del Ψs e dell’apertura stomatica. A stomi
chiusi l’amido era sintetizzato ed il destino dello zucchero non era chiaro: era
utilizzato come substrato per la respirazione oppure estruso nell’apoplasto.
Quest’ipotesi fu abbandonato con la scoperta del K+.
Il saccarosio delle cellule di guardia
diventa il soluto osmoticamente attivo
dominante a seguito dell’efflusso di K+
Apertura giornaliera all’alba
associata primariamente
ad accumulo di K+
Chiusura associata
primariamente
a diminuzione di saccarosio
20
K+
agente osmotico
primario
Trasporto di Ioni K+ dalle cellule
sussidiarie alle cellule di guardia
0,5 M K+sono sufficienti a far
diminuire il Ψs di circa 2 MPa
Esperimenti su cellule
di guardia di Vicia
faba prive di cellule
sussidiarie
mostrano
che le cellule di
guardia si aprono solo
se alla soluzione è
aggiunto K+
21
In alcune specie (Allium cepa) il
flusso di K+ è accompagnato da
ClEntra per bilanciare
elettricamente l’ingresso
del K+ e fuoriesce con il K+
In altre specie lo ione K+
è neutralizzato dallo ione
malato2Deriva dall’idrolisi dell’amido
contenuto nei cloroplasti delle cellule
di guardia
Le proporzioni di Cl- e
malato2-per bilanciare
gli ioni K+dipendono
dalle condizioni
ambientali
In cella climatica:
malato2In serra:
Cl22
Schema di apertura stomatica (3)
stoma chiuso (GC flaccide) → saccarosio/potassio/malato/cloro → basso Ψs →
riduzione Ψw → ingresso H2O (osmosi) → aumento pressione → stoma aperto (GC
turgide)
Per chiudere stoma, processo inverso
Comunque, poiché l’assorbimento del potassio è associato con l’apertura degli
stomi nella mattina, mentre la perdita di saccarosio è maggiormente associata
con la chiusura nel pomeriggio
Schema di apertura stomatica finale (giorno/notte)
stoma chiuso (GC flaccide) → assorbimento potassio e cloro, sintesi malato →
basso Ψs → riduzione Ψw → assorbimento H2O (osmosi) dalle cellule sussidiarie →
aumento di pressione → stoma aperto (GC turgide) → ||||| → riduzione
saccarosio (potassio, cloro, malato) → Ψs aumento → Ψw aumento → perdita H 2O
→ riduzione pressione → stoma aperto(GC flaccide)
23
CELL WALL
MEMBRANE
Gas Exhange (e)
Stomatal swelling
A. Guard-cell symplast
accumulate solutes from guardcell apoplast.
B. Water flows into guard cells
osmotically.
C. Radial micellation of
cellulose microfibrils prevents
increase of cell diameter.
E. Water influx increases pressure, but
water is incompressible, so guard-cell
volume increases. The increase results
from stretching of the dorsal wall.
D. Inner wall is strong and
cannot be stretched.
24
Qual’ è il meccanismo del flusso degli ioni K + e Cl- e malato2- ?
Accumulo di K+
cellule di guardia
Aumento di pH
Cellule sussidiarie
Esterne
Terminale
Epidermiche
si comportano all’opposto
25
Uso sperimentale delle luci blu e rossa per studiare le vie
osmoregolative associate ai movimenti stomatici
Attivazione H+-ATPasi
Estrude protoni H+
Δ pH
Em
Forza proton motrice
―pmf‖
Forza guida per il trasporto passivo secondario
del K+ attraverso canali regolati dal voltaggio
26
Ingresso di K+
Fuoriuscita di H+
Alcalinizzazione
del citoplasma
Gradiente di pH
0,5-1 unità
pH alcalino attiva
l’enzima PEPc
citoplasma
Simporto H+- Cl-
Favorisce la produzione
di acidi organici per le
specie che utilizzano il
malato come controione
E’ stato recentemente evidenziato che l’influsso di K+ potrebbe essere
anche energizzato da un simporto H+- K+ ad alta affinità
Apertura stomi: Processo attivo
che necessita di ATP per
il funzionamento della pompa protonica
Respirazione
Fosforilazione fotosintetica
27
Gas Exchange (g)
Ion Transport—stomatal opening
Proton extrusion makes
membrane potential (MP) more
negative and acidifies
apoplast.
Water influx
Potassium uptake.
Thermodynamics: MP
Mechanism: MP & wall
acidification activate the Kin
channel
Inside
cell
Membrane
28
Gas Exchange (j)
Ion Transport—stomatal closing
A. Anion efflux shifts the
membrane potential to a less
negative position.
B. Potassium efflux.
Thermodynamics: MP
Mechanism: MP
the Kout channel
Inside
cell
activates
Membrane
29
Fattori di controllo/regolazione della funzionalità delle cellule
di guardia
• Intensità e qualità della luce
le cellule di guardia
funzionano come
valvole idrauliche
sensibili a molteplici
fattori ambientali
• Concentrazioni intracellulari di CO2
•Ormoni
•Temperatura
•Umidità relativa
•Vento
Luce
Segnale ambientale dominante che
controlla i movimenti stomatici
In generale: luce = apertura; buio = chiusura (tranne per le piante CAM)
30
Regolazione temporale
delle aperture stomatiche
piante C3 –C4
Apertura alle prime
luci dell’alba
Chiusura al tramonto
Favorisce la fotosintesi
spesso a spese di
un’elevata traspirazione
Previene inutile perdita
di H2O
Gli stomi si aprono non appena i flussi fotonici incidenti aumentano e viceversa
Giorno
Notte
Questo deve interfacciarsi con la disponibilità di H2O
31
Buona disponibilità idrica
( zone temperate,
caldo-umide)
Scarsa disponibilità idrica
( zone calde o mattinate
molto calde)
Piante CAM
Pianta scambia H2O per i prodotti della
fotosintesi senza disidratarsi con gli stomi aperti
durante
la
mattinata
per
permettere
un’efficiente fotosintesi
Stomi si apriranno di meno o addirittura
resteranno chiusi
Piante adattate al deserto con fotosintesi CAM, gli stomi
sono aperti di notte e chiusi di giorno
Questo permette a queste piante di
mantenere basso il rapporto di traspirazione
Piante C3 = 500
Piante C4 = 250
Moli di H2O traspirata
Moli di CO2 assorbita
Piante CAM = 50
Significato ecofisiologico
32
Aumento di flussi fotonici incidenti
Radiazione fotosinteticamente attiva (400-700 nm)
Stomi aperti
Intensità minima per
apertura stomi
Da 1/1000 a 1/30 della piena luce solare
effetto indiretto della luce
+ luce + fotosintesi
+ consumo di CO2
33
Esiste una risposta
specifica della luce
Espressione di due sistemi di
fotorecettori distinti
Dipendente dalla fotosintesi
delle cellule di guardia (luce rossa)
Guidata da una risposta
alla luce blu
Componente non fotosintetica
Luce blu
Luce rossa
•Meccanismo on-off
•Attivazione molto veloce
•Meno efficiente
•Tempo di latenza 25sec
•Pochi minuti per max velocità
•15 volte più efficiente
Rigonfiamento delle
cellule di guardia e
apertura stomatica
34
Luce blu
Luce rossa
Illuminazione solare
luce rossa e/o luce blu
zeaxantina
Attivazione pompa
H+-ATPasi
È modulata dal cloroplasto delle cellule di guardia
Bloccato da inibitori della fotosintesi
Stimolazione differenziale di carotenoidi e
clorofille selettivamente eccitati
dall’eccitazione di questi pigmenti si attivano vie di regolazioni
35
O2
Basso livello di O2 stimola apertura degli stomi
CO2
Pressione parziale di CO2 negli
spazi intercellulari del mesofillo fogliare
Ogni fattore ambientale che aumenta la velocità di fotosintesi
Consumo di CO2 e quindi abbassamento della pCO2 interna
Bassa pCO2 interna
Apertura degli stomi
A bassi livelli di irradianza la pCO2 interna può costituire il principale fattore di
controllo ma ad alti livelli di irradianza più importante è il ruolo della luce
36
37
H2O
Proteggere contro un’eccessiva perdita di H2O
Ci sono due meccanismi attraverso i quali la perdita di H2O regola la chiusura
stomatica, uno passivo ed uno attivo.
•Controllo idropassivo – Una semplice riduzione di turgore (fogliare – cellule di
guardia), dovuta a perdita di H2O, che porta alla chiusura che deterrmina la
chiusura dello stoma. La pianta non sta “intenzionalmente” chiudendo lo stoma,
ma è una semplice conseguenza della siccità.
•Controllo idroattivo – questo meccanismo è uno col quale la pianta realmente
sembra monitorare il suo stato idrico. Quando il Ψw cade sotto il valore critico, si
innescano una cascata di eventi che determinano la chiusura degli stomi.
Presumibilmente la pianta sta misurando la pressione (turgore)e quindi sintetizza
e rilascia un “antitraspirante‖ che è traslocato verso le cellule di guardia per
determinare la chiusura.
38
Acido Abscissico
ABA
Ormone che controlla la chiusura degli stomi
ABA è sintetizzato a livelli bassi
continuamente nelle cellule del mesofillo e
accumulato principalmente nei cloroplasti
In condizioni di stress idrico
Concentrazioni da 7 a 13 µM
aumenta di circa 50 volte
39
ABA
Gli stomi si chiudono quando
il Ψw fogliare decresce…
40
Chiusura dello stoma è
associata con un
incremento di contenuto di
ABA
No H2O
Maize
leaves
No H2O
ABA sembra coinvolto nella
chiusura degli stomi anche prima
che si crei una variazione nello
stato idrico fogliare
41
42
43
ABA prodotto nelle radici disidratate degli
starti superiori del suolo e trasportato nelle
foglie può rendere capaci le piante di
“anticipare” la condizione di stress idrico e
indurre la chiusura degli stomi (feed
forward response).
Ri-distribuzione dell’ABA
dallo xilema durante lo
stress idrico
44
Gas Exchange
ion transport—ABA action
ABA
Prodotto nelle radici e trasportato
nelle foglie, o prodotto dalle
stesse foglie o dalle cellule di
guardia
Attiva i flussi primari di Ca++
• inibisce la pompa protonica
• inibisce fuoriuscita di H+
•depolarizzazione della
membrana
Inside
cell
ABA activates the Kout channel via
cytosolic alkalinization.
• blocca canali ingresso di K+
• favorisce canali di uscita di K +
Membrane
Modified from Outlaw WH, Jr. Integration of cellular and physiological
45
functions of guard cells. CRC Crit Rev Plant Sci 22: 503-529
Stato di siccità del suolo e disidratazione
parziale del sistema radicale
è necessario stress
idrico fogliare ?
“sistema di allarme precoce”
Traslocazione dell’ABA
dalle radici al germoglio
attraverso il flusso traspiratorio
Chiusura degli stomi
Auxine-citochinine
fusicoccina
Apertura degli stomi
Attiva pompa H+-ATPasi
Azione dei diserbanti di tipo auxinico
Variano da una specie all’altra
•Soglia
•Rapidità
•modo
di risposta
Variano nell’ambito della stessa specie
Variano con il grado di adattamento all’ambiente
Nell’ambito della stessa foglia (età, condizione 46
di
sviluppo, posizione del germoglio)
 ―Agli stomi è stato affidato il compito di fornire il")