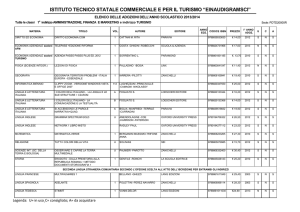
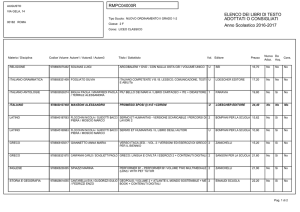
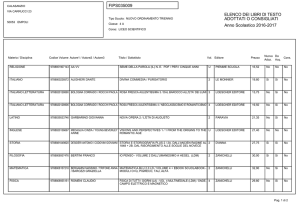
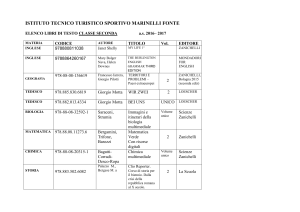
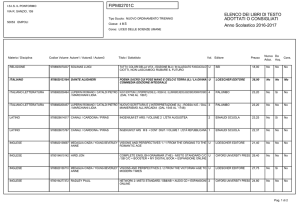
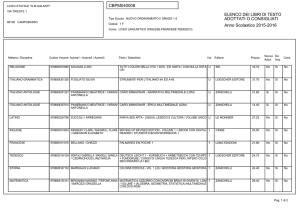
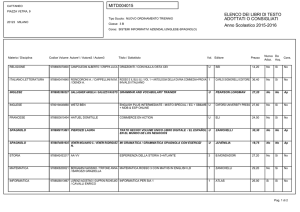
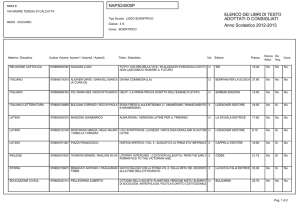
Capitolo 11
Il controllo dell’espressione
genica
Copyright © 2006 Zanichelli editore
La regolazione genica nei procarioti e negli eucarioti
11.1 Le proteine che interagiscono con il DNA
attivano e disattivano i geni dei procarioti in risposta
ai cambiamenti ambientali
Figura 11.1A
Copyright © 2006 Zanichelli editore
Colorizzata SEM 7000×
I primi risultati nel campo del controllo genico furono
ottenuti grazie a esperimenti condotti sul batterio
Escherichia coli.
L’operone del lattosio
• Spesso, nei procarioti, i geni con funzioni
interconnesse sono aggregati in strutture chiamate
operoni.
• Il vantaggio principale che deriva dal raggruppare i
geni in operoni è che l’espressione di questi geni
può essere facilmente coordinata.
Copyright © 2006 Zanichelli editore
Proteine di regolazione si legano a sequenze di
controllo nel DNA e attivano o disattivano gli operoni in
risposta a cambiamenti ambientali. OPERONE
Gene
regolatore Promotore Operatore
Geni per metabolizzare il lattosio
DNA
mRNA
Proteina
Repressore
attivo
L’RNA-polimerasi non può attaccarsi
al promotore
Operone del lattosio disattivato (lattosio assente)
DNA
mRNA
Proteina
Lattosio
L’RNA-polimerasi
si lega al
promotore
Repressore
inattivo
Enzimi per l’utilizzo del lattosio
Figure 11.1B,C Operone del lattosio attivato (il repressore è disattivato dal lattosio)
Copyright © 2006 Zanichelli editore
Altri tipi di operone
• L’operone trp è simile all’operone del lattosio
(operone lac) ma funziona in modo un po’
differente.
• Questo operone controlla la sintesi degli enzimi per
la produzione del triptofano.
Promotore Operatore
Geni
DNA
Repressore
attivo
Repressore
attivo
Triptofano
Repressore
inattivo
Repressore
inattivo
Lattosio
Figura 11.1D
Copyright © 2006 Zanichelli editore
Operone lac
Operone trp
11.2 Il processo di differenziamento dà origine a
una grande varietà di cellule specializzate
• La regolazione dell’espressione genica negli
organismi eucariotici, soprattutto nei pluricellulari,
è più complicata che nei batteri.
• Durante le ripetute divisioni cellulari che portano
uno zigote a diventare un organismo pluricellulare
adulto, le singole cellule vanno incontro al
differenziamento e diventano cellule specializzate
nella struttura e nelle funzioni.
Copyright © 2006 Zanichelli editore
• Differenti tipi di cellule umane producono differenti
tipi di proteine a seconda delle combinazioni di
geni che sono attivi in ciascuna di esse.
• A seconda dei geni attivi, ciascuna cellula assume
una specifica struttura e funzione.
Figura 11.2
Cellule muscolari
Copyright © 2006 Zanichelli editore
Cellule del pancreas
Cellule del sangue
11.3 Le cellule differenziate possono conservare
tutto il loro potenziale genetico
Le cellule differenziate esprimono solo una piccola
percentuale dei loro geni.
Radice di una pianta
di carota
Singola cellula
Figura 11.3
Copyright © 2006 Zanichelli editore
Cellule radicali
coltivate in una
soluzione
nutritiva
Le cellule
Germoglio Pianta adulta
si dividono
nel terreno di coltura
11.4 Il modo in cui il DNA si ripiega all’interno dei
cromosomi eucariotici contribuisce a regolare
l’espressione genica
Un nucleosoma è formato da un filamento di DNA
avvolto attorno a un nucleo proteico centrale costituito
da otto istoni.
Copyright © 2006 Zanichelli editore
Spiralizzazione del DNA in un cromosoma eucariotico:
Doppia elica di DNA
(2 nm di diametro)
Istoni
Linker
TEM
«Perle di una
collana»
Nucleosoma
(10 nm di diametro)
Figura 11.4
Copyright © 2006 Zanichelli editore
TEM
Fibra elicoidale compatta Superavvolgimento
(30 nm di diametro)
(300 nm di diametro)
Cromosoma in metafase
700
nm
• Questa fibra elicoidale compatta si avvolge e
ripiega ulteriormente.
• Presumibilmente, la spiralizzazione del DNA
impedisce l’espressione dei geni in quanto non
consente all’enzima RNA-polimerasi (e ad altre
proteine che contribuiscono alla trascrizione) di
prendere contatto con il DNA.
Copyright © 2006 Zanichelli editore
11.5 Nelle femmine dei mammiferi uno dei due
cromosomi X è disattivato in tutte le cellule
Nelle femmine dei mammiferi uno dei due cromosomi X
si presenta fortemente condensato in tutte le cellule
somatiche e quasi del tutto inattivo (disattivazione del
cromosoma X).
Embrione
Divisione
cellulare
e inattivazione
casuale del
cromosoma X
Cromosomi X
Figura 11.5
Copyright © 2006 Zanichelli editore
Allele
per il pelo Allele per
arancione il pelo nero
Due cellule nella
popolazione adulta
X attivo
X inattivo
X inattivo
X attivo
Pelo
arancione
Pelo
nero
11.6 Negli eucarioti la trascrizione è controllata da
complessi aggregati di proteine
• Come i procarioti, anche gli eucarioti utilizzano
proteine di regolazione che, legandosi al DNA,
attivano o disattivano la trascrizione.
• I meccanismi di controllo comprendono proteine
che si legano a segmenti specifici del DNA (con
sistemi più complessi di quelli dei procarioti).
Copyright © 2006 Zanichelli editore
I fattori di trascrizione
Alcune proteine di regolazione, chiamate fattori di
trascrizione, favoriscono l’inizio della trascrizione.
Intensificatori
Promotore
Gene
DNA
Induttori
Fattori di
trascrizione
Altre
proteine
RNA-polimerasi
Ripiegamento
del DNA
Figura 11.6
Copyright © 2006 Zanichelli editore
Trascrizione
La coordinazione dell’espressione genica negli eucarioti
• Negli eucarioti la coordinazione dell’espressione
genica sembra dipendere dalla presenza di una
specifica sequenza enhancer (o di diversi
enhancer) in ogni gene che fa parte dello stesso
«gruppo di lavoro».
• Diverse copie di fattori di trascrizione che
riconoscono queste sequenze di DNA si legano a
esse promuovendo la trascrizione simultanea dei
geni.
Copyright © 2006 Zanichelli editore
11.7 L’RNA eucariotico può essere modificato in vari
modi
• Completata la trascrizione, i segmenti non codificanti
(introni) vengono rimossi grazie al processo di splicing.
• In alcuni casi la cellula svolge lo splicing in maniera
differente e genera diverse molecole di mRNA a partire
dallo stesso trascritto di RNA.
Esone
DNA
Trascritto
di RNA
Splicing dell’RNA
Figura 11.7
Copyright © 2006 Zanichelli editore
mRNA
oppure
11.8 Anche la traduzione e le ultime fasi
dell’espressione genica sono soggette a regolazione
Dopo che l’RNA è stato modificato e trasferito dal
nucleo al citoplasma, avvengono altre forme di controllo
dell’espressione genica:
• demolizione più o meno rapida dell’mRNA;
• attivazione della traduzione;
• modificazione dei polipeptidi tradotti;
• demolizione delle proteine.
Copyright © 2006 Zanichelli editore
La degradazione dell’mRNA
• Le molecole di mRNA non sono eterne: nel
citoplasma si trovano, infatti, degli enzimi che
hanno il compito di degradarle.
• Il tempo di sopravvivenza delle molecole di mRNA
è un fattore importante che regola la quantità di
proteine assemblate dalla cellula.
Copyright © 2006 Zanichelli editore
L’innesco della traduzione
• Anche il processo di traduzione dell’mRNA in
polipeptidi offre una possibilità di regolazione
genica.
• Tra le molecole coinvolte nella traduzione vi sono
numerose proteine che hanno la funzione di
regolare l’inizio della sintesi proteica.
Copyright © 2006 Zanichelli editore
L’attivazione delle proteine
S
S
SH
SH
SH
Polipeptide iniziale
(inattivo)
Figura 11.8
Copyright © 2006 Zanichelli editore
Taglio
S
S
Ripiegamento del polipeptide
e formazione dei legami S—S
S
SH
SH
SH
I polipeptidi che si formano dopo la traduzione non
sempre sono già pronti ad agire: spesso devono essere
modificati per diventare funzionali.
S
S S
Polipeptide ripiegato
(inattivo)
S
S
S S
Insulina
(ormone attivo)
La demolizione delle proteine
• Un altro meccanismo di controllo che opera dopo
la traduzione è la demolizione selettiva delle
proteine.
• Alcune proteine che controllano il tasso metabolico
delle cellule vengono demolite in pochi minuti o in
poche ore.
Copyright © 2006 Zanichelli editore
11.9 Una visione d’insieme
dell’espressione genica negli
eucarioti
I molteplici meccanismi che
controllano l’espressione genica
sono analoghi alle valvole di controllo
delle tubazioni.
Figura 11.9
Copyright © 2006 Zanichelli editore
La clonazione degli animali
11.10 Gli animali possono essere clonati tramite
trasferimento nucleare
Nucleo della cellula del donatore
Cellula
del donatore
Impianto della blastocisti Nascita di un clone
in una madre surrogata del donatore
(clonazione riproduttiva)
Accrescimento in coltura
Rimozione
Inclusione del
nucleo di una cellula per produrre una blastocisti
del nucleo
dalla cellula uovo somatica del donatore
adulto
Figura 11.10
Copyright © 2006 Zanichelli editore
Cellule staminali
embrionali si sviluppano
dalla blastocisti
e crescono in coltura
Formazione di cellule
specializzate a partire
dalle cellule staminali
(clonazione terapeutica)
COLLEGAMENTI
11.11 La clonazione terapeutica può produrre cellule
staminali che hanno grandi potenzialità mediche
• La clonazione riproduttiva dei mammiferi è utile per la
ricerca, l’agricoltura e la medicina.
• L’uso di cellule staminali
embrionali è però correlato a
problemi di natura tecnica e di
ordine etico, connessi all’utilizzo
di embrioni umani.
Figura 11.11A
Copyright © 2006 Zanichelli editore
• Le cellule staminali adulte sono cellule indifferenziate
presenti in molti tessuti adulti che sostituiscono le
cellule che non sono in grado di dividersi.
• Anche le cellule staminali adulte possono crescere in
coltura e dare origine a cellule differenziate.
Cellule ematiche
Cellule staminali
adulte
nel midollo osseo
Cellule nervose
Colture di cellule
staminali embrionali
Cellule muscolari del cuore
Figura 11.11B
Copyright © 2006 Zanichelli editore
Diverse condizioni Diversi tipi di cellule
di coltura
differenziate
• Contrariamente alle cellule embrionali staminali, le
cellule staminali adulte si trovano già sulla strada
del differenziamento ed è molto più difficile isolarle
e coltivarle in laboratorio.
• Normalmente le cellule staminali adulte danno
origine solo a un gruppo limitato di tipi di cellule.
• Un terzo modo per ottenere cellule staminali è
quello di prelevarle dal sangue del cordone
ombelicale o dalla placenta al momento del parto.
Copyright © 2006 Zanichelli editore
Il controllo genetico dello sviluppo embrionale
11.12 Le reazioni in sequenza dell’espressione genica
e la comunicazione cellulare dirigono lo sviluppo di un
animale
Le prime intuizioni sulle relazioni esistenti tra espressione
genica e sviluppo embrionale scaturirono studiando i mutanti
dei moscerini della frutta (Drosophila melanogaster).
Occhio
SEM 50×
Antenna
Figura 11.12A
Copyright © 2006 Zanichelli editore
Capo di un moscerino della frutta
normale
Zampa
Capo di un moscerino della frutta mutante
Un gradiente di espressione
genica controlla lo sviluppo del
moscerino della frutta a partire
dalla cellula uovo fecondata:
Figura 11.12B
Copyright © 2006 Zanichelli editore
• Un gene omeotico (detto anche omeogene) è il
gene di controllo principale che regola una serie di
altri geni adibiti allo sviluppo del piano strutturale di
un organismo.
• Un gruppo di geni omeotici dei moscerini della
frutta ordina alle cellule dei segmenti del capo e
del torace (la parte centrale del corpo) di formare
rispettivamente le antenne e le zampe.
Copyright © 2006 Zanichelli editore
11.13 Sequenze di trasduzione del segnale
trasformano i messaggi ricevuti dalla membrana
plasmatica in reazioni di risposta all’interno della
cellula
• Uno dei fattori più importanti nello sviluppo di un
organismo è la comunicazione tra cellule, un
meccanismo con cui certe proteine o altri tipi di
molecole portano i messaggi dalle cellule che li
trasmettono alle cellule (bersaglio) che li ricevono.
• Questo consente di coordinare meglio le attività
cellulari in un organismo adulto.
Copyright © 2006 Zanichelli editore
Una sequenza di
trasduzione del segnale
trasforma un segnale che
arriva sulla membrana di
una cellula bersaglio in
una risposta specifica
all’interno della cellula.
Cellula che trasmette il segnale
Molecola segnale
1
Membrana
Recettore
plasmatica
proteico
3
2
Cellula bersaglio
Ripetitori proteici
Fattore di
trascrizione
(attivato)
4
Nucleo
DNA
5
mRNA
Trascrizione
Nuova
proteina
6
Figura 11.13
Copyright © 2006 Zanichelli editore
Traduzione
11.14 I geni che sono alla base dello sviluppo sono
molto antichi
I geni omeotici del moscerino della frutta contengono
sequenze di nucleotidi, chiamate homeobox, che sono
molto simili in molti tipi di organismi eucarioti.
Cromosoma del moscerino Cromosomi del topo
Embrione di moscerino (10 ore)Embrione di topo (12 ore)
Figura 11.14
Copyright © 2006 Zanichelli editore
Moscerino adulto
Topo adulto
Le basi genetiche del cancro
11.15 Il cancro si può scatenare a causa di mutazioni
di geni che controllano la divisione cellulare
• Le cellule tumorali, che si dividono in modo
incontrollato, derivano da mutazioni in geni codificanti
per proteine che influiscono sul ciclo cellulare.
• Le cellule possono diventare cancerose se il loro ciclo
è alterato a causa dell’espressione di oncogèni di
origine virale o dovuti a mutazioni causate da agenti
cancerogeni.
Copyright © 2006 Zanichelli editore
Le mutazioni genetiche possono
provocare il cancro
L’apoptosi (morte cellulare programmata) è un meccanismo molto
importante per la prevenzione della cancerogenesi. Nel cancro la
cellula perde il controllo del ciclo cellulare a causa di mutazioni di due
tipi di geni:
• i protoncogeni, che codificano per proteine che promuovono il ciclo
cellulare e inibiscono l’apoptosi;
• i geni soppressori dei tumori, che codificano per proteine che
inibiscono il ciclo cellulare e favoriscono l’apoptosi.
18
Sylvia S. Mader Immagini e concetti della biologia © Zanichelli editore, 2012
I proto-oncogèni
• I proto-oncogèni sono geni che possono essere
trasformati in ocogèni da una mutazione nel DNA.
• I proto-oncogèni codificano per i fattori di crescita (le
proteine che stimolano la divisione cellulare) e per altre
proteine che li regolano.
DNA del Proto-oncogène
Copie multiple
Mutazione
del gene
all’interno del gene
Oncogène
Proteina iperattiva
Figura 11.15A in quantità normale
Copyright © 2006 Zanichelli editore
Proteina normale
in eccesso
Il gene si è spostato verso un nuovo locus del
DNA e viene regolato da nuovi geni di controllo
Nuovo promotore
Proteina normale
in eccesso
Gli oncosoppressori
Il cancro può essere indotto anche da alterazioni dei
geni (detti oncosoppressori) i cui prodotti inibiscono la
divisione cellulare.
Gene oncosoppressore
Proteina
normale
Divisione
cellulare
controllata
Figura 11.15B
Copyright © 2006 Zanichelli editore
Gene oncosoppressore mutato
Proteina
alterata
Divisione
cellulare
incontrollata
11.16 Le proteine codificate dagli oncogeni e gli
oncosoppressori alterati interferiscono con le
normali sequenze di trasduzione del segnale
I prodotti normali degli oncogèni e dei geni soppressori
sono proteine coinvolte nelle sequenze di trasduzione
del segnale.
Copyright © 2006 Zanichelli editore
Le proteine degli
oncogèni possono
stimolare la
sequenza di
trasduzione del
segnale:
Fattore
di crescita
Recettore
Cellula bersaglio
Ripetitore
proteico
iperattivo
Prodotto normale
del gene ras
(prodotto da un
oncogène)
inv ia i segnali
autonomamente
Ripetitori
proteici
Fattore di
trascrizione
(attivato)
DNA
Nucleo
Figura 11.16A
Copyright © 2006 Zanichelli editore
Proteina che
stimola
la divisione cellulare
Trascrizione
Traduzione
Le proteine degli
oncosoppressori
possono inibire la
sequenza di
trasduzione del
segnale:
Fattore che
inibisce la
crescita
Ripetitori
proteici
Fattore di
trascrizione
(attivato)
Recettore
Fattore di trascrizione
non funzionante
(prodotto dal gene
oncosoppressore p53)
non può attivare la
trascrizione.
Prodotto normale
del gene p53
Trascrizione
Traduzione
Figura 11.16B
Copyright © 2006 Zanichelli editore
Proteina
che inibisce
la divisione cellulare
Assenza delle proteine
(divisione cellulare
non inibita)
11.17 Lo sviluppo del cancro ha origine da
mutazioni genetiche multiple
Perché si abbia un completo sviluppo del cancro, è
necessario che si verifichi più di una mutazione nelle
cellule somatiche.
Copyright © 2006 Zanichelli editore
Le mutazioni che conducono all’insorgenza di un tumore
possono accumularsi in una linea di cellule somatiche:
Cromosomi
1
mutazione
2
mutazioni
Cellula normale
4
mutazioni
Cellula maligna
Figura 11.17B
Copyright © 2006 Zanichelli editore
3
mutazioni
Il cancro
procede
lentamente e
diventa
maligno
gradualmente
La cancerogenesi,
ossia lo sviluppo di un
tumore maligno, richiede
l’intervento di numerose
mutazioni; il processo
risulta quindi graduale.
22
Sylvia S. Mader Immagini e concetti della biologia © Zanichelli editore, 2012
Le cellule del tumore
rilasciano fattori di crescita
che promuovono
l’angiogenesi, ossia la
formazione di nuovi vasi
sanguigni.
Le cellule tumorali
invadono anche i vasi
linfatici e sanguigni, e
vengono così trasportate
ad altre parti del corpo.
Quando le cellule
cancerose danno origine a
nuovi tumori in distretti
lontani dal tumore
originario, si dice che il
cancro è in metastasi.
23
Sylvia S. Mader Immagini e concetti della biologia © Zanichelli editore, 2012
COLLEGAMENTI
11.18 Evitare l’esposizione agli agenti cancerogeni
può ridurre il rischio del cancro
• Gli agenti che causano il cancro, cioè i fattori che
alterano il DNA e rendono una cellula cancerosa,
sono detti cancerogeni.
• Evitare l’esposizione agli agenti cancerogeni e
scegliere altri stili di vita può aiutare a ridurre il
rischio di cancro.
Copyright © 2006 Zanichelli editore
Fattori di rischio del cancro nei paesi sviluppati:
Tabella 11.18
Copyright © 2006 Zanichelli editore
La terapia del cancro tende a colpire
le cellule malate in modo selettivo
La diagnosi del cancro richiede un’attenta valutazione della salute
generale e un esame medico del paziente (esami del sangue e delle
urine, indagini per immagini, biopsia, esami endoscopici e chirurgici,
test genetici).
L’asportazione chirurgica è indicata per i cancri in situ, ma visto il
rischio di lasciare alcune cellule malate, gli interventi sono spesso
preceduti e/o seguiti da chemioterapia e/o radioterapia.
La chemioterapia è il trattamento del cancro con farmaci e tende ad
agire in modo selettivo sulle cellule cancerose.
La radioterapia si basa sull’uso di radiazioni ionizzanti che
colpiscono con forte energia le cellule cancerose danneggiandole o
distruggendole.
24
Sylvia S. Mader Immagini e concetti della biologia © Zanichelli editore, 2012