ISTITUTO SUPERIORE DI SANITÀ
Il sistema cardiovascolare fetale:
un modello numerico con algoritmi genetici
come metodo di identificazione
Mauro Grigioni, Carla Daniele, Umberto Morbiducci,
Giacomo Di Benedetto, Giuseppe D’Avenio, Vincenzo Barbaro
Laboratorio di Ingegneria Biomedica
ISSN 1123-3117
Rapporti ISTISAN
02/4
Istituto Superiore di Sanità
Il sistema cardiovascolare fetale: un modello numerico con algoritmi genetici come metodo di identificazione.
Mauro Grigioni, Carla Daniele, Umberto Morbiducci, Giacomo Di Benedetto, Giuseppe D’Avenio, Vincenzo
Barbaro
2002, 33 p. Rapporti ISTISAN 02/4
Lo sviluppo della cardiochirurgia fetale, che si pone come frontiera ultima nel trattamento delle malformazioni
cardiache congenite, non può prescindere dalla dettagliata conoscenza della circolazione sanguigna nel sistema
cardiovascolare fetale. Oggetto del presente studio è lo sviluppo e l’identificazione di un modello matematico del
sistema cardiovascolare fetale globale, con il fine di individuare relazioni e differenze esistenti nella circolazione
sanguigna nella parte superiore e inferiore del corpo. La modellizzazione del cuore è stata realizzata mediante due
capacità tempo-variabili, rappresentanti le relazioni pressione-volume di ciascuno dei due ventricoli. Il sistema
vascolare fetale è stato rappresentato mediante due modelli di Windkessel a sei elementi. La identificazione dei
parametri del modello è stata ottenuta facendo uso degli algoritmi genetici, i quali operano seguendo le leggi della
teoria evoluzionistica. Al fine di validare il modello del sistema cardiovascolare fetale umano proposto, i risultati
dello studio numerico eseguito sul modello identificato sono stati confrontati con i dati presenti in letteratura e con
quelli derivanti da misure sperimentali eseguite dagli autori stessi. Ulteriore obiettivo di tale studio, oltre a quello di
fornire uno strumento per studiare la differente distribuzione sanguigna nei vari distretti vascolari nella parte
superiore e inferiore del corpo del feto, è quello di individuare il ruolo dell’istmo aortico; in particolare, esso potrebbe
configurarsi come un valido aiuto nella valutazione del bilancio dinamico durante la assistenza meccanica al circolo.
Parole chiave: Emodinamica fetale, Modello matematico, Algoritmi genetici, Elastanza ventricolare tempovariabile
Istituto Superiore di Sanità
The foetal cardiovascular system: a numerical model with genetic algorithms as identification technique.
Mauro Grigioni, Carla Daniele, Umberto Morbiducci, Giacomo Di Benedetto, Giuseppe D’Avenio, Vincenzo
Barbaro
2002, 33 p. Rapporti ISTISAN 02/4 (in Italian)
The development of foetal cardiac surgery, the ultimate goal in the treatment of congenital cardiac
malformations, needs to be supported by a detailed knowledge of the blood circulation in the foetal cardiovascular
system. This study presents a mathematical model of the foetal human global cardiovascular system, developed with
the aim to clarify the relationships and differences existing between upper and lower body circulation. The heart has
been modelled with two time-varying capacities, each representing the respective ventricle’s pressure-volume
relationship. The foetal vascular system has been represented using two six-element Windkessel models, for the
upper and lower body respectively. The identification of the set of circuital and elastance functions parameters of the
model has been obtained by using genetic algorithms, whose mode of operation follows the laws of the evolutionary
theory. The results of our numerical study on the identified model have been compared with data collected from
measurements and literature, with the aim of validating the proposed global cardiovascular system model of human
foetus. Moreover, this model is intended not only as an instrument to investigate the differences in blood distribution
between the different vascular districts in the upper and lower foetal body but also to identify the role of the aortic
isthmus, the small tract of vessel connecting upper and lower foetal vascular beds; it may also represent a useful tool
in the assessment of the dynamic balance during mechanical assistance to circulation.
Key words: Foetal haemodynamics, Mathematical model, Genetic algorithms, Ventricular time-varying elastance
Il rapporto è accessibile online dal sito di questo Istituto: www.iss.it/pubblicazioni.
© Istituto Superiore di Sanità 2002
INDICE
Introduzione ......................................................................................................................................................... 1
Metodi ....................................................................................................................................................................... 2
Il modello del sistema cardiovascolare fetale.................................................................................................. 2
Identificazione dei parametri del sistema cardiovascolare fetale................................................................ 5
Gli algoritmi genetici.................................................................................................................................... 5
Introduzione agli algoritmi genetici..................................................................................................... 5
Funzionamento degli AG....................................................................................................................... 6
Operatori degli AG.................................................................................................................................. 9
Ottimizzazione di una funzione............................................................................................................ 11
Codifica delle soluzioni negli AG........................................................................................................ 15
Implementazione del metodo di identificazione mediante AG.................................................................... 16
Il problema numerico .......................................................................................................................................... 18
Risultati.................................................................................................................................................................... 19
Discussione .......................................................................................................................................................... 29
Bibliografia ............................................................................................................................................................ 32
i
ii
INTRODUZIONE
Il trattamento chirurgico di malformazioni cardiache congenite in età fetale potrebbe
consentire di prevenire quelle alterazioni che, indotte dalle malformazioni stesse, accompagnano
la crescita dell’individuo. L’approccio a livello fetale potrebbe pertanto permettere di ottenere
risultati migliori di quelli ottenibili attualmente agendo chirurgicamente in età neonatale o
infantile (1). In letteratura sono presenti vari lavori (2-9) a testimoniare come un bypass
cardiaco o cardiopolmonare sia tecnicamente realizzabile su modelli animali di cardiochirurgia
fetale.
Lo sviluppo della chirurgia cardiaca fetale necessita di essere supportato da una dettagliata
conoscenza dell’emodinamica locale in zone e organi specifici nonché delle mutue interazioni
prodotte da alterazioni in territori distali. Le limitate informazioni fisiologiche che attualmente
sono note sulle condizioni della circolazione fetale in aree quali il cervello, la placenta o il
fegato sono inferite dai valori degli ‘indici di resistenza’ dei vasi (10-13). In letteratura sono
presenti diversi modelli della circolazione fetale (14-21): alcuni di tali modelli sono del tipo a
compartimenti (14-20), ma fra questi ve ne sono diversi (15-17) che, non contemplando
compliance vascolari, non sono utilizzabili al fine di indagini di tipo emodinamico; altri modelli
sono stati sviluppati come sistemi di trasporto, associando a valori emodinamici medi il
consumo di ossigeno (18-20). In passato alcuni ricercatori hanno individuato un modello basato
sull’analogia idraulico-elettrica del cuore di feto (21-23), con il fine di studiare gli effetti delle
variazioni del precarico e del postcarico sul cuore stesso: in tali modelli il cuore di feto veniva
considerato come un singolo ventricolo equivalente (22, 23), oppure veniva considerato in toto
(21); la determinazione dei parametri circuitali veniva compiuta sulla base di ipotesi
fluidodinamiche, misure anatomiche ed emodinamiche.
Oggetto di questo studio è la individuazione di un modello dell’intero sistema
cardiovascolare fetale umano, basato sull’analogia idraulico-elettrica, nel quale i comportamenti
dei ventricoli destro e sinistro (entrambi eiettanti nell’aorta) sono rappresentati mediante
compartimenti elettrici separati, uno per ciascun ventricolo appunto. Una volta identificato, il
modello è stato utilizzato per studiare le differenze fra la circolazione relativa alla parte
superiore del corpo e quella relativa alla parte inferiore, nonché per studiare il ruolo svolto
dall’isthmus aorticus nella circolazione fetale. Il comportamento del cuore è stato modellizzato
mediante due capacità tempo-variabili, che stanno a rappresentare i due ventricoli: la legge
biomeccanica che lega la pressione e il volume ventricolare è descritta dalla funzione elastanza
tempo-variabile (la funzione elastanza essendo definita da un set di quattro parametri). La rete
vascolare fetale è stata rappresentata utilizzando due modelli di Windkessel a sei elementi, uno
per la parte superiore del corpo, l’altro per la parte inferiore.
La identificazione dei parametri vascolari e la completa caratterizzazione delle funzioni
elastanza che modellizzano i due ventricoli è stata ottenuta sfruttando la capacità selettiva degli
algoritmi genetici (24, 25), la cui funzione di fitness è stata costruita utilizzando i dati raccolti in
letteratura riguardanti feti umani e feti di pecora (esiste una forte analogia fra il comportamento
dei cuori di feto umano e di feto di pecora) e relativi ai valori massimi della pressione
ventricolare, al volume eiettato, alla distribuzione percentuale di sangue della gittata cardiaca
combinata fetale e alla distribuzione percentuale del ritorno venoso. Il principale risultato
ottenuto in questo studio è stato la identificazione del modello complessivo, in grado di
descrivere adeguatamente il comportamento fisiologico dell’istmo aortico. In questo studio
viene inoltre mostrato un confronto fra misure in vivo (9) e i valori calcolati dal modello
proposto (forma d’onda della pressione ventricolare sinistra e diagramma pressione-volume del
ventricolo sinistro).
1
METODI
In questa sezione vengono descritti un modello matematico dell’intero sistema
cardiovascolare fetale basato sull’analogia idraulico-elettrica, il metodo di identificazione dei
parametri ‘cardiovascolari’ del modello e una descrizione del modo con il quale è stato condotto
lo studio numerico del modello.
Il modello del sistema cardiovascolare fetale
Il cuore di un feto differisce sostanzialmente da quello di un adulto, essendo caratterizzato
dalla presenza di numerosi shunt fra la circolazione arteriosa e venosa (12). In Figura 1 viene
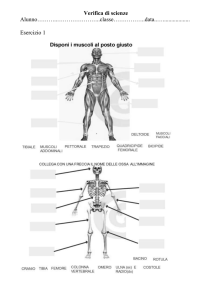
mostrato lo schema di un cuore di feto.
istmo aortico
aorta ascendente
aorta
dotto arterioso
arteria polmonare
tronco polmonare
ventricolo
destro
ventricolo
sinistro
vena cava
superiore
atrio
sinistro
atrio
destro
vena cava
inferiore
setto
interatriale
vena polmonare
aorta discendente
Figura 1. Schema di un cuore di feto
È possibile notare, confrontandolo con un cuore di adulto, la esistenza di due differenze
sostanziali, costituite dalla presenza del dotto arterioso e del setto interatriale. Proprio a causa
della presenza del dotto arterioso, che realizza fisicamente una connessione tra il tronco
polmonare e l’aorta discendente, i due ventricoli eiettano sangue simultaneamente in aorta
discendente: il ventricolo sinistro attraverso il tratto vascolare costituito dall’aorta ascendente e
dall’istmo aortico, il ventricolo destro attraverso il dotto arterioso stesso. Il setto interatriale
2
svolge il ruolo di mettere in comunicazione diretta le due camere atriali fetali, facendo sì che il
sangue venoso che ritorna, attraverso la vena cava inferiore (VCI), nell’atrio destro, fluisca poi
in parte nel ventricolo destro e in parte verso il ventricolo sinistro attraversando il setto
interatriale stesso.
Per quanto riguarda invece il sangue venoso che ritorna verso l’atrio destro attraversi la vena
cava superiore (VCS), esso fluisce quasi interamente nel ventricolo destro.
Nel modello oggetto di questo studio, per caratterizzare il comportamento del ventricolo di
feto si è fatta l’ipotesi che la relazione fra la pressione intraventricolare sinistra P(t) e il volume
del ventricolo sinistro V(t) fosse quella definita da Suga et al. (26), in accordo con l’equazione:
P(t) = E(t) [V(t)-V0]
[1]
in cui il parametro V0 rappresenta il valore di un volume di riferimento senza alcun significato
fisiologico diretto (23) e E(t) rappresenta la funzione elastanza tempo-variabile ventricolare, che
può essere espressa come:
E(t) = Emax · En (t)
[2]
Nella equazione [2] En (t) altro non è se non la funzione elastanza ventricolare normalizzata
rispetto al suo valore massimo di ampiezza ed Emax – il valore massimo di E(t) – rappresenta una
misura della massima capacità contrattile delle fibre miocardiche, che Suga et al. (26) hanno
dimostrato essere indipendente dal battito cardiaco, dal precarico e dal postcarico.
Per la modellizzazione della fase sistolica di ciascuno dei due ventricoli si è fatto uso, per la
legge di variazione di En (t), della relazione definita da Menigault et al. (23), ricavata
interpolando i dati di elastanza normalizzati in ampiezza e in tempo misurati da Suga et al. (26);
tale legge di variazione è rappresentata dalla seguente equazione:
En(
6
5
t
t
t
) = 5,412 ⋅
− 20,066 ⋅
+ 25,542 ⋅
T max
T
max
T
max
T max
t
3
2
4
[3]
t
t
t
− 13,714 ⋅
+ 2,714 ⋅
+ 1, 085 ⋅
+ 0,029
T max
T max
T max
nella quale Tmax costituisce l’intervallo di tempo che intercorre tra l’inizio del ciclo cardiaco e
l’istante in corrispondenza del quale si ha E=E max, ossia l’istante in cui la funzione elastanza
assume il suo valore massimo. La fase del ciclo cardiaco relativa al riempimento ventricolare,
sia per il ventricolo destro che per il sinistro, è stata descritta mediante l’equazione:
P(t) = P0 · exp [(k0 · V(t)]
[4]
in cui P(t) e V(t) in cui rappresentano sempre, ripettivamente, pressione e volume ventricolari,
mentre P0 and k0 sono due parametri costanti. L’equazione [4] descrive la relazione
esponenziale fra pressione e volume ventricolare durante la diastole: tale relazione è stata
definita da Menigault et al. (23) mediante estrapolazione dai diagrammi pressione-volume
misurati da Frank (27) e relativi a ventricolo sinistro. Pertanto, mediante le equazioni [3] e [4] è
stato possibile descrivere la relazione pressione-volume per entrambi i ventricoli durante
l’intero ciclo cardiaco, in seguito alla identificazione, per il ventricolo destro come per il
sinistro, dei valori dei quattro parametri che caratterizzano la funzione ventricolare fetale Emax,
V0 , P0 , k0 . Poiché le equazioni [3] e [4] sono scaturite da correlazioni matematiche su dati
misurati riguardanti il ventricolo sinistro, in questo studio si è fatta l’ipotesi che le medesime
3
leggi biomeccaniche governano il comportamento dei due ventricoli: tale ipotesi scaturisce dalla
considerazione che la natura delle fibre miocardiche che costituiscono i due ventricoli è la
medesima.
Per analogia elettrica, il funzionamento dei due ventricoli è stato studiato considerando
ciascuno di essi come una capacità tempo-variabile accoppiata con il sistema cardiovascolare, il
quale è stato schematizzato mediante tre modelli di Windkessel a due elementi per la parte
superiore del corpo e tre modelli di Windkessel a due elementi per quella inferiore. In questo
modo è stato possibile discriminare fra la rete vascolare superiore e quella inferiore. All’interno
di ciascuna delle due reti vascolari principali, la superiore e la inferiore, sono stati modellizzati i
tre letti vascolari (arterioso, capillare e venoso), ciascuno mediante un modello di Windkessel a
due elementi: ogni coppia (Ri ,Ci ) descrive la resistenza al flusso sanguigno e la capacità relativa
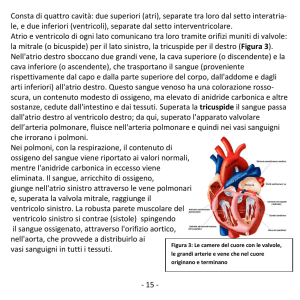
al volume di sangue dell’i-esimo letto vascolare. In Figura 2 è mostrato il modello analogoelettrico del sistema cardiovascolare fetale.
MVD
MVS
AV D
AV S
CS
CD
Resistenza (mmHg ml-1 s)
Capacità (ml mmHg -1)
R1
R2
R3
R4
R5
R6
R7
R8
R9
R10
R11
R12
R13
C1
C2
C3
C4
C5
C6
capacità arteriosa inferiore
capacità capillare inferiore
capacità venosa inferiore
capacità arteriosa superiore
capacità capillare superiore
capacità venosa superiore
CD
CS
MVS
MVD
AVS
AVD
ventricolo dx: capacità tempo-variabile
ventricolo sx: capacità tempo-variabile
valvola atrio-ventricolare sx
valvola atrio-ventricolare dx
valvola aortica ventricolo sx
valvola arteriosa ventricolo dx
resistenza arteriosa inferiore
resistenza capillare inferiore
resistenza venosa inferiore
resistenza arteriosa superiore
resistenza capillare superiore
resistenza venosa superiore
resistenza di ingresso al cuore destro (vista da VCI)
resistenza di ingresso al cuore sinistro (vista da VCI)
resistenza di ingresso al cuore destro (vista da VCS)
resistenza polmonare
res. arteria polmonare principale + dotto arterioso
res. arco aortico + aorta ascendente
resistenza istmo aortico
Figura 2. Modello analogo-elettrico del sistema cardiovascolare fetale
L’istmo aortico, che costituisce il segmento vascolare che unisce i due sistemi vascolari
paralleli perfusi dal ventricolo destro e dal ventricolo sinistro (vedi Figura 1), è stato
4
modellizzato mediante una resistenza (R13 in Figura 2) situata tra l’aorta ascendente e la
discendente. Per quanto riguarda il comportamento compliante dei due atrii, poiché le due
camere atriali nel feto risultano essere comunicanti (vedi Figura 1) per la presenza del setto
interatriale, e in virtù del fatto che la pressione atriale ‘segue’ la pressione venosa (28), si è fatta
l’ipotesi di considerare i due atrii come una singola camera equivalente, la cui capacità è stata
considerata inclusa parte nella capacità del letto venoso superiore (C6 in Figura 2), parte nella
capacità del letto venoso inferiore (C3 in Figura 2).
Il ritorno venoso verso i due ventricoli è stato modellizzato mediante le tre resistenze R9 , R8 ,
R7 (vedi Figura 2). Esse rappresentano rispettivamente la resistenza di ingresso del ventricolo
destro vista dalla circolazione ‘superiore’ (il ritorno venoso proveniente dalla parte superiore del
corpo è interamente diretto verso il ventricolo destro, attraverso la vena cava superiore (28)); la
resistenza di ingresso del ventricolo sinistro vista dalla circolazione ‘inferiore’ insieme con
quella polmonare; la resistenza di ingresso del ventricolo destro vista dalla circolazione
‘inferiore’ (il sangue venoso proveniente dalla parte inferiore del corpo ritorna attraverso la
vena cava inferiore e, a causa della presenza del setto interatriale, si dirige in parte verso il
ventricolo sinistro e in parte verso quello destro; il sangue venoso che ritorna verso il cuore dai
polmoni è invece totalmente diretto verso il ventricolo sinistro) (28).
I vasi connessi direttamente con le uscite dei ventricoli sono stati modellizzati mediante delle
resistenze: l’aorta ascendente con una resistenza (R12 in Figura 2) posta fra il ventricolo sinistro
e la rete vascolare arteriosa superiore; il dotto arterioso mediante una resistenza (R11 in Figura 2)
posta fra il ventricolo destro e la rete vascolare arteriosa inferiore. La circolazione polmonare
fetale è stata rappresentata mediante una resistenza (R10 in Figura 2) che connette il ventricolo
sinistro con il ritorno venoso comprensivo dell’atrio sinistro ‘intrinseco’ e diretto verso il
ventricolo sinistro stesso (come detto in precedenza). Le valvole arteriose e atrioventricolari
sono state modellizzate tramite diodi, facendo l’assunzione che nessuna caduta di pressione e
nessun ritardo temporale potesse essere introdotto dalle valvole stesse (valvole come interruttori
ideali).
Nel modello individuato, la circolazione coronarica non è stata presa in considerazione –
essa corrisponde al 2÷4% della gittata combinata cardiaca fetale (28) –, assumendo il contributo
di quest’ultima come cumulativo con quello eiettato dal ventricolo sinistro verso la parte
superiore del corpo, il quale poi ritorna nel ventricolo destro.
Identificazione dei parametri
del sistema cardiovascolare fetale
La stima dei parametri circuitali e la completa caratterizzazione delle funzioni elastanza che
modellizzano i due ventricoli è stata ottenuta mediante l’utilizzo di algoritmi genetici (AG) (2425).
Gli algoritmi genetici
Introduzione agli algoritmi genetici
La teoria evoluzionistica dei sistemi biologici lega le possibilità di sopravvivenza di un
individuo principalmente alla sua capacità di adattamento all’ambiente (valore adattativo o
fitness) e i mezzi che, in ciascun individuo, caratterizzano tale capacità, risiedono nel suo
patrimonio genetico.
5
La selezione naturale funziona nel seguente modo:
– gli individui di una popolazione con un valore adattativo migliore tendono a sopravvivere
e a riprodursi più a lungo;
– le caratteristiche, codificate nei loro geni, vengono trasmesse alla propria discendenza e
tendono a propagarsi nelle nuove generazioni;
– nella riproduzione sessuale, i cromosomi della prole sono una mescolanza di quelli dei
loro genitori;
– le caratteristiche della prole sono in parte ereditate dai genitori e in parte sono il risultato
di nuovi geni creati durante il processo di riproduzione.
La teoria dell’evoluzione della specie può essere utilizzata per simulare al computer dei
processi di affinamento progressivo delle potenziali soluzioni alle problematiche connesse ai
sistemi complessi, quindi gli algoritmi evoluzionistici, algoritmi legati alla teoria
dell’evoluzione, sono delle ottimizzazioni e ricercano procedimenti ispirati alla genetica e al
processo di evoluzione naturale.
L’algoritmo di calcolo che permette al computer di simulare in poco tempo un processo
evolutivo di molti secoli prende il nome specifico di AG.
La Tabella 1 riporta lo schema dell’analogia computer-natura.
Tabella 1. Analogia natura-computer
NATURA
COMPUTER
Individuo
Soluzione del problema
Popolazione
Insieme delle possibili soluzioni
Fitness
Qualità di una soluzione
Cromosoma
Rappresentazione di una soluzione
(insieme di parametri)
Gene
Parte della rappresentazione di una soluzione
(parametro o“grado” di libertà)
Crescita
Decodifica della rappresentazione di soluzioni
Crossover, Mutazione
Operatori di ricerca
Selezione naturale
Riutilizzo di buone (sotto-)soluzioni
Funzionamento degli AG
Molti problemi per i quali sono stati sviluppati gli AG sono quelli di ottimizzazione, i quali
si presentano frequentemente nelle applicazioni. Dato un difficile problema di ottimizzazione è
possibile trovare un algoritmo efficiente le cui soluzioni sono approssimativamente ottimali.
Gli AG appartengono alla classe degli algoritmi probabilistici, eppure sono molto diversi
dagli algoritmi casuali poiché combinano elementi di ricerca stocastica con quelli di ricerca
guidata.
Un’altra importante proprietà di questi metodi di ricerca basati sulla genetica è che essi
conservano una popolazione di soluzioni potenziali, mentre tutti gli altri metodi trattano singoli
punti dello spazio di ricerca delle soluzioni.
Gli AG compiono inoltre una ricerca multidirezionale trattando una popolazione di
potenziali soluzioni e favoriscono la formazione di informazione e scambio tra tali direzioni.
6
Per poter impiegare gli AG è necessario innanzi tutto stabilire una codifica della soluzione
del problema trattato in termini di stringhe binarie; la ricerca di una buona soluzione diventa
quindi la ricerca di particolari stringhe binarie. Si parte da una popolazione iniziale di individui,
ciascuno dei quali mantiene una soluzione codificata nel suo DNA (stringa binaria) e tale
popolazione è sottoposta a una evoluzione simulata: ad ogni generazione le soluzioni
relativamente “buone” si riproducono, mentre quelle relativamente “cattive” muoiono. Per
distinguere tra le differenti soluzioni si usa una funzione oggettiva (di valutazione) che gioca il
ruolo di ambiente.
La riproduzione delle stringhe avviene, in analogia con gli esseri viventi a riproduzione
sessuata, tramite accoppiamento, cioè tramite la fusione delle due stringhe appartenenti ai
genitori. In pratica ciò si realizza stabilendo uno o più punti in cui spezzare ciascuna delle
stringhe, per combinare poi opportunamente tali parti delle stringhe (crossover). I discendenti
così prodotti non vanno a rimpiazzare i genitori, i quali sono le stringhe che hanno fornito i
migliori valori della funzione di valutazione; essi rimpiazzano invece le stringhe della
precedente generazione che hanno dato valori meno buoni della stessa. In questo modo la
popolazione globale non cambia ad ogni generazione.
Quindi, al fine di evitare la produzione di popolazioni troppo uniformi (che esplorano una
regione ristretta dello spazio delle soluzioni), si produce su una piccola percentuale di individui
una mutazione. Quest’ultima altera arbitrariamente uno o più geni di un cromosoma selezionato,
per mezzo di uno scambio casuale con probabilità pari alla “percentuale” di mutazione PM.
L’intuizione dietro l’operatore mutazione è l’introduzione di qualche variabilità extra nella
popolazione.
Un AG, come ogni programma evoluzionistico, per un particolare problema deve avere le
seguenti cinque componenti:
– rappresentazione genetica per le potenziali soluzioni del problema;
– modo per creare una popolazione di soluzioni iniziali;
– funzione di valutazione che gioca il ruolo di ambiente, valutando le soluzioni in termini
del loro fitness;
– operatori genetici che alterano la composizione dei figli;
– valori per i vari parametri usati dall’algoritmo genetico (es. dimensione della popolazione
e probabilità di applicare operatori genetici, ecc.).
La struttura di un semplice AG è la stessa di ogni programma evoluzionistico (Figura 3).
begin
t=0
inizializza P(t)
valuta P(t)
while (not condizione_di_terminazione) do
begin
t = t+1
seleziona P(t) da P(t-1)
altera P(t)
valuta P(t)
end
end
Figura 3. Struttura semplice di AG
7
In tale algoritmo, P(t) rappresenta la popolazione di individui (soluzioni) all’iterazione t,
ovvero P(t) = {xt 1 , xt 2 , …, xt n }.
Gli AG sono stati introdotti da Holland e l’algoritmo da lui presentato viene riportato in
Figura 4.
begin
t = 0;
P(t) = INIZIALIZZA_POPOLAZIONE( );
repeat
for i=1 to m do
f(Ai) = CALCOLA_FITNESS(Ai);
fitness_medio = CALCOLA_FITNESS_MEDIO (P(t))
for k=1 to m do
begin
Ak = SELEZIONA (P(t))
/* ogni individuo viene selezionato confrontando il proprio valore di fitness con il fitness_medio*/
if (Pc > random(0,1)) then
begin
Ai = SELEZIONA (P(t));
Afiglio = CROSSOVER(Ai, Ak);
if (PM > random(0,1)) then
MUTAZIONE_FIGLIO (Afiglio);
if (PI > random(0,1)) then
INVERSIONE_FIGLIO (Afiglio);
P(t+1)=AGGIORNA_POPOLAZIONE (Afiglio);
end
end;
t = t+1
until(condizione_di_terminazione);
end
Figura 4. Algoritmo di Holland
Con PC si indica la probabilità di applicare l’operatore di crossover, con PM la probabilità di
mutare i geni dell’individuo trattato e con PI la probabilità di invertire la sequenza dell’intero
codice genetico di un individuo.
L’algoritmo di Holland funziona come segue:
sia P(0) la popolazione iniziale generata casualmente e P(t) la popolazione di n individui
all’istante t, l’algoritmo genera iterativamente una nuova popolazione P(t+1) a partire da P(t)
applicando alcuni operatori genetici. Questi operatori modificano degli individui nella
popolazione P(t) ottenendo così una popolazione P(t+1) che contiene un maggior numero di
individui con un valore di fitness migliore.
La condizione di terminazione può essere fissata da:
– un numero massimo di iterazioni, raggiunto il quale l’algoritmo si ferma;
– un controllo sul miglioramento del valore di fitness fornita dalla soluzione corrente;
quando si notano miglioramenti poco apprezzabili delle soluzioni, l’algoritmo viene
interrotto.
8
Operatori degli AG
Gli operatori degli AG sono:
1) Selezione
L’operatore di selezione ha la funzione di scegliere gli individui ai quali devono essere
applicati gli operatori genetici.
La selezione degli individui può essere fatta casualmente, oppure per mezzo del valore di
fitness. In quest’ultimo caso gli individui con valore di fitness migliore hanno una
probabilità più alta di essere selezionati per generare i figli.
Questa operazione di selezione degli individui con valore di fitness più alto, può essere
vista come una metafora della sopravvivenza naturale di Darwin.
Infatti in natura il valore di fitness di un individuo è determinato dall’abilità di superare
tutti gli ostacoli dell’ambiente. La fase di selezione è applicata sulla popolazione
corrente: le stringhe selezionate sono messe in una popolazione intermedia e gli operatori
genetici sono applicati alla popolazione intermedia, per creare la popolazione successiva.
Ottenuta quest’ultima si dice allora che nell’algoritmo genetico è trascorsa una
generazione.
2) Rimpiazzamento
Ci sono due modi principali per rimpiazzare un individuo nella popolazione:
– rimpiazzamento generazionale
La popolazione corrente è rimpiazzata dalla successiva; in questo caso le dimensioni
della popolazione corrente e della successiva devono essere uguali.
– rimpiazzamento a stato continuo
I nuovi individui generati rimpiazzano gli individui peggiori della popolazione
corrente quando i figli hanno un valore di fitness migliore. La popolazione
successiva è identica alla popolazione corrente eccetto per l’ultimo figlio generato:
in letteratura questo modello è conosciuto come modello statico di popolazione.
Un’importante differenza tra i due metodi è che nel rimpiazzamento a stato continuo i
nuovi figli sono immediatamente disponibili per la riproduzione, perciò gli AG hanno
l’opportunità di esplorare subito gli individui più promettenti appena essi vengono creati.
Questi due modelli di rimpiazzamento vengono impiegati coerentemente col modello di
generazione adottato: il rimpiazzamento generazionale col modello di generazione
discreto e il rimpiazzamento a stato continuo col modello di generazione continuo.
3) Crossover
L’operatore di crossover, come già visto, prende parti dei due genitori e crea un nuovo
individuo combinando tali parti. In questo modo i figli contengono informazioni
genetiche di entrambi i genitori.
I punti di crossover (di solito due) sono scelti casualmente. I nuovi individui
consisteranno di parti alternate delle stringhe dei genitori.
Nonostante sia casuale, questo scambio di informazione genetica rende gli algoritmi
genetici molto versatili; infatti una parte buona di un genitore può rimpiazzare una parte
meno buona dell’altro genitore, creando un figlio migliore dei genitori (Figura 5 e 6).
9
Genitore 1
Genitore 2
0010011010
1110010001
si estrae una posizione a caso tra 1 e 10,
(per esempio 6) e si pone il punto di separazione
alla sua destra; si ottiene:
0010011010
accoppiando ciascuna metà
con la metà complementare,
si ottiene:
1110010001
0010010001
1110011010
Figlio 1
Figlio 2
Figura 5. Esempio di crossover ad un punto
Genitore 1
Genitore 2
1|010|0101 si estraggono due posizioni a caso 0|011|1110
tra 1 e 8, (per esempio 1 e 4), e si pone il punto
di separazione alla sua destra si ottiene:
1|011|0101
0|010|1110
Figlio 1
Figlio 2
Figura 6. Esempio di crossover a due punti
4) Inversione
L’operatore di inversione prende due punti casuali del cromosoma e inverte la sequenza
di geni tra questi due punti. L’importanza del cambiamento così effettuato dipende dalla
particolare scelta per rappresentare la soluzione (Figura 7).
101| 10 10 |1
101| 01 01 |1
Figura 7. Esempio di inversione
5) Mutazione
L’operatore di mutazione consiste nello scambiare i valori di uno o più geni in un
individuo appena creato; nel caso della codifica binaria della soluzione, si nega il valore
dei bit selezionati per effettuare tale operazione (Figura 8).
10
000110010
000111010
Figura 8. Esempio di mutazione di un gene
L’operatore di mutazione è impiegato principalmente per introdurre delle varianti nelle
soluzioni della popolazione corrente che consentano di sfuggire dagli ottimi locali quando
la maggioranza delle soluzioni converge verso questi ultimi.
Ottimizzazione di una funzione
In questo paragrafo si discutono le caratteristiche basilari di un AG per l’ottimizzazione di
una funzione semplice ad una variabile. Si consideri, per esempio, la funzione definita da:
f ( x ) = x ⋅ sen(10 π ⋅ x ) + 1,0
Il problema consiste nel trovare la x nell’intervallo [-1,2] che massimizzi la funzione f, cioè
trovare x0 tale che f(x0) ≥ f(x) ∀ x ∈ [-1,2] (Figura 9).
Figura 9. Grafico della funzione f(x)
L’analisi della funzione è relativamente semplice. Si determinano gli zeri della derivata prima f ':
f ′( x ) = sen (10π ⋅ x ) + 10π ⋅ x ⋅ cos(10 π ⋅ x ) = 0
Tale formula è equivalente alla seguente funzione (riportata in grafico in Figura 10):
tan(10ð ⋅ x) = −10ð ⋅ x .
11
Figura 10. Grafico della funzione tan(10π
π x) = -10π
πx
La precedente equazione ha un numero infinito di soluzioni:
2i − 1
xi =
+ åi ,
per i = 1,2,...
20
x0 = 0 ,
2i + 1
xi =
− εi ,
per i = −1,−2,... ,
20
dove i termini ε i rappresentano una sequenza decrescente di numeri reali (per i = 1,2,…; e i = 1,-2,…) tende a zero.
È da notare inoltre che la funzione f raggiunge i suoi massimi locali in xi , per i intero dispari
e i minimi locali in xi , per i intero pari. Poiché il dominio del problema è l’intervallo [-1,2], la
funzione f raggiunge il suo massimo in:
x19 =
37
+ å19 = 1,85 + å19 ,
20
dove f(x19 ) è poco maggiore di:
π
f (1,85 ) = 1,85 ⋅ sen18 π + + 1,0 = 2,85 .
2
Si suppone di voler costruire un algoritmo genetico che risolva il problema precedente, cioè
di voler massimizzare la funzione f.
Come cromosoma per rappresentare i valori reali della variabile x si utilizza un vettore
binario. La lunghezza del vettore dipende dalla precisione richiesta che, in questo esempio, è di
6 cifre dopo il punto decimale. Il dominio della variabile x ha lunghezza 3; la precisione
richiesta implica che l’intervallo [-1,2] deve essere diviso in almeno 3x106 intervalli della stessa
dimensione. Ciò significa che sono richiesti 22 bit per il vettore binario (cromosoma), poiché:
2097152 = 2 21 < 3000000 ≤ 2 22 = 4194304 .
12
La conversione di una stringa binaria <b21 b20 …b0 > in un numero reale x dell’intervallo [-1,2]
è diretta e si completa in due passi:
Si converte la stringa binaria <b21 b20 …b0 > da base 2 a base 10:
(< b21b20 ...b0 > )2 = ∑ bi ⋅ 2i
21
i =0
10
= x′ ,
Si trova il corrispondente valore di x:
x = −1 + x′ ⋅
3
,
2 −1
22
dove -1 è il limite sinistro del dominio e 3 è la lunghezza dello stesso.
Per esempio un cromosoma (1000101110110101000111) rappresenta il numero 0.637197, in
quanto:
x ′ = (100010111 0110101000 111) 2 = 2288967
e:
x = −1 + 2288967 ⋅
3
= 0,637197 .
4194303
Naturalmente i cromosomi (0000000000000000000000) e (1111111111111111111111)
rappresentano gli estremi del dominio -1 e 2, rispettivamente.
Il processo di inizializzazione è molto semplice: si crea una popolazione di cromosomi, dove
ogni cromosoma è un vettore binario di 22 bit. Tutti i 22 bit di ogni cromosoma vengono
inizializzati in maniera casuale. La funzione di valutazione eval per vettori binari v è
equivalente alla funzione f:
eval(v) = f(x)
dove il cromosoma v rappresenta il valore reale x. Essa, come già visto, gioca il ruolo di
ambiente, valutando le soluzioni potenziali in termini del loro fitness. Per esempio dati i tre
cromosomi:
v 1 = (1000101110110101000111),
v 2 = (0000001110000000010000),
v 3 = (1110000000111111000101),
che corrispondono ai valori x1 = 0,637197, x2 = 0,958973 e x3 = 1,627888, rispettivamente, la
loro funzione di valutazione li stimerà come segue:
eval(v 1 ) = f(x1) = 1,586345,
eval(v 2 ) = f(x2) = 0,078878,
eval(v 3 ) = f(x3) = 2,250650.
Chiaramente il cromosoma v 3 è il migliore tra i tre cromosomi in quanto la sua valutazione
restituisce il valore più alto.
Durante la fase di alterazione dell’AG si utilizzeranno i due classici operatori genetici:
mutazione e crossover. Come già visto, la mutazione altera uno o più geni (posizioni di un
13
cromosoma) con una probabilità pari alla percentuale di mutazione. Si suppone che sia stato
selezionato il quinto gene del cromosoma v 3 per la mutazione: poiché esso corrisponde allo 0,
verrà cambiato con 1. Così il cromosoma v 3 dopo la mutazione sarà:
v'3 = (1110100000111111000101).
Questo cromosoma rappresenta il valore:
x'3 = 1,721638
e la sua funzione di valutazione è:
f(x' 3 ) = -0,082257.
Ciò significa che da questa mutazione in particolare ne consegue un significante decremento
della qualità del cromosoma v 3 . D’altra parte, se fosse stato selezionato per la mutazione il
decimo gene del cromosoma v 3 , allora:
v''3 = (1110000001111111000101)
i cui valori corrispondenti sono:
x'3 = 1,630818
e:
f(x''3 ) = -2,343555,
e si avrebbe avuto un miglioramento sul valore originale di:
f(x3 ) = 2,250650.
Si illustra ora l’operatore di crossover sui cromosomi v 2 e v 3 . Si suppone che il (solo) punto
di crossover sia stato selezionato casualmente dopo il quinto gene:
v 2 = (00000|01110000000010000),
v 3 = (11100|00000111111000101).
I due “figli” risultanti sono:
v'2 = (00000|00000111111000101),
v'3 = (11100|01110000000010000).
Quest’ultimi vengono valutati in:
f(v’ 2 ) = f(-0,998113-0,082257 f(v ' 2 ) = f( −0,998113) = 0,940865 ,
f(v '3 ) = f(1,666028 ) = 2,459245 .
Si noti che il secondo figlio ha una valutazione migliore di entrambi i genitori.
14
Per questo problema particolare sono stati usati i seguenti parametri: dimensione della
popolazione pop_size = 50, probabilità di crossover P c = 0,25, probabilità di mutazione PM = 0,01.
Di seguito si riportano alcuni risultati sperimentali per un tale sistema genetico. La Tabella 2
infatti fornisce il numero di generazioni per il quale si nota un aumento nella funzione di
valutazione, insieme ai valori della funzione: il miglior cromosoma dopo 150 generazioni è v max
corrispondente al valore 1,850773. Come ci si aspettava, xmax = 1,85 + ε e f(xmax) è poco
maggiore di 2,85.
Tabella 2. Risultati di 150 generazioni
Numero di generazioni
Funzione di valutazione
1
6
8
9
10
12
39
40
51
99
137
145
1,441942
2,250003
2,250283
2,250284
2,250363
2,328077
2,344251
2,345087
2,738930
2,849246
2,850217
2,850227
Codifica delle soluzioni negli AG
La codifica delle soluzioni di un problema non sono sempre immediate e si possono
presentare diversi casi a seconda dei parametri a cui ci troviamo di fronte:
• Parametri interi
–
Se il valore p varia nell’intervallo {0,1,…..,2 N-1}, allora si può codificare usando una
rappresentazione binaria del tipo:
e =< b 0 , b 1 ,...b N > ,
con N bit b i ∈ {0,1} tale che:
N
p = ∑ b i ⋅ 2 i −1 .
i =1
–
Se p ∈ {M,M+1,…..,M+2N-1}, allora si può codificare p – M nello stesso modo.
–
Se p ∈ {0,1,……,L-1} e non esiste un
N
tale che L = 2N , allora si hanno problemi.
Possibili soluzioni
a) Prendere N = log2 (L)+1 e rimappare ogni stringa
< b 0 , b1 ,...b N >10 ≥ L ,
in p = L-1 (ritaglio).
[Problema: ogni p tra 0 e L-2 è rappresentato da una stringa binaria mentre
p=L-1 è rappresentato da 2N-L stringhe.]
15
b) Prendere N = log2 (L)+1 e rimappare ogni
e10 = <b0 , b1 , … bN >10
in
p=
e10
( L − 1) (riduzione).
2 −1
N
[Problema: alcuni valori di p sono rappresentati da due stringhe binarie
(leggera deviazione).]
• Parametri a valori reali
Possono essere rappresentati numeri di punto fisso o interi con riduzione e
quantizzazione.
Se p ∈ [pmin,pmax] allora e è la rappresentazione binaria della parte intera di
(2N-1)p/(p max-p min).
Per parametri a valori reali c’è un conflitto tra la volontà di voler considerare geni brevi
per una buona convergenza e il bisogno di conoscere il risultato con una certa
precisione.
Possono essere usati alfabeti non binari per calcolare parametri che possono assumere
solo un numero piccolo di valori. I caratteri di tale alfabeto sono rappresentati da singoli
geni (atomi) non divisibili. A volte anche interi o parametri reali sono considerati
“atomici” (simboli di un alfabeto molto grande).
• Vettori di parametri
Sono codificati combinando le codifiche [b 0i ,….., b Ni ] di ogni componente.
Le codifiche di parametri multipli possono essere concatenate:
[b
o intervallate:
1
1
M
M
],
M
1
M
].
,..., b N ,..., b0 ,..., b N
1
0
,..., b0 ,..., bN ,..., bN
[b
0
L’ordine dei parametri in un vettore è molto importante, specialmente per parametri
concatenati.
Implementazione del metodo di identificazione
mediante AG
Per l’identificazione dei parametri del modello del sistema cardiovascolare fetale è stato
utilizzato un AG steady state (non è consentito alla popolazione di aumentare o diminuire nel
passaggio da una generazione alla successiva) con una popolazione costituita da 50 individui
(cromosomi).
Ciascun cromosoma è stato costruito con 27 geni e a ciascun gene è stato consentito di
variare fra un fissato valore minimo e uno massimo.
In Tabella 3 sono riportati i geni e i corrispondenti intervalli di variazione fissati. Gli
intervalli di esistenza per i geni sono stati individuati a partire dai valori emodinamici e dai dati
raccolti in letteratura (23, 29), nonché da misure effettuate dagli autori (9).
16
Tabella 3. Parametri (geni) e corrispondenti intervalli di variabilità
Parametro
Intervallo
di variabilità
Emax (VD)
Emax (VS)
V0 (VD)
V0 (VS)
P0 (VD)
K0 (VD)
P0 (VS)
K0 (VS)
R1
(5 ÷ 18)
(5 ÷ 18)
(-6 ÷ 0)
(-6 ÷ 0)
(0,01 ÷ 0,2)
(0,1 ÷ 1)
(0,01 ÷ 0,2)
(0,1 ÷ 1)
(0,1 ÷ 1)
Parametro
Intervallo
di variabilità
(0,1 ÷ 5)
(0,1 ÷ 5)
(0,1 ÷ 1)
(0,1 ÷ 5)
(0,1 ÷ 5)
(0,1 ÷ 1)
(0,1 ÷ 1)
(0,1 ÷ 1)
(10 ÷ 30)
R2
R3
R4
R5
R6
R7
R8
R9
R10
Parametro
R11
R12
R13
C1
C2
C3
C4
C5
C6
Intervallo
di variabilità
(0,1 ÷ 2)
(0,1 ÷ 2)
(0,1 ÷ 5)
(0,1 ÷ 1)
(0,1 ÷ 1)
(15 ÷ 30)
(0,1 ÷ 1)
(0,1 ÷ 1)
(15 ÷ 30)
VD: ventricolo destro
VS: ventricolo sinistro
Partendo da una popolazione di 50 individui, generata casualmente, ciascuno di tali individui
è stato ‘valutato’ mediante la applicazione di una funzione di fitness (24, 25), la quale
rappresenta l’errore quadratico medio calcolato riferendosi ai valori riportati in Tabella 4. La
scelta effettuata di minimizzare i valori delle grandezze calcolate rispetto alle pressioni
massime, i volume eiettati dai ventricoli e le portate, ha consentito al modello di essere
consistente con una circolazione fetale realistica. I valori delle grandezze utilizzati per la
identificazione sono stati individuati in letteratura (12, 28), ad eccezione del valore massimo di
pressione e del volume eiettato dal ventricolo sinistro, i quali sono stati misurati dagli autori (9).
Tabella 4. Valori delle grandezze emodinamiche utilizzati per costruire la funzione di fitness
Grandezza emodinamica
Valore fisiologico
Fonte
Pressione massima VS
68 mmHg
(9)
Volume Eiettato VS (VEVS)
1,7 ml
(9)
Volume di sangue che ritorna
in VS in in un ciclo cardiaco
1,7 ml
Stimato da VEVS
Distribuzione della Gittata Cardiaca
Combinata (GCC)
25% aorta ascendente
(12)
10% istmo aortico
58% dotto arterioso
7% arteria polmonare
Ritorno sanguigno venoso
75% da circolazione inferiore +
circolazione polmonare
Estrapolato da (12)
25% dalla circolazione superiore
Pressione massima VD
80 mmHg
(12)
Volume Eiettato VD (VEVD)
3,2 ml
Stimato da VEVS
e dalla distribuzione di GCC
Volume di sangue che ritorna
in VD in in un ciclo cardiaco
3,2 ml
Stimato da VEVS
VD: ventricolo destro
VS: ventricolo sinistro
17
Tutte le quantità utilizzate nella funzione di fitness, ad eccezione dei valori massimi di
pressione ventricolare, sono rappresentative dell’intero ciclo cardiaco. Ciò sta a significare che
la procedura di identificazione è stata costruita senza introdurre riferimenti diretti alla
morfologia delle forme d’onda delle grandezze emodinamiche (pressioni, portate e volumi).
Alla fase di valutazione della popolazione corrente segue la fase di selezione di una una
popolazione a partire dalla precedente, selezione operata per mezzo della procedura di selezione
Rank Elitist (25), la quale garantisce la convergenza e protegge da eventuali rischi di
stagnazione (quando tutti gli individui della popolazione presentano la medesima qualità intorno
ad un valore medio, si esercita una piccola ‘pressione selettiva’, ossia le soluzioni migliori sono
favorite soltanto leggermente rispetto a quelle peggiori). Sugli individui che compongono la
nuova popolazione selezionata vengono applicati gli operatori genetici di crossover e mutazione
non uniforme, per creare nuove soluzioni (25). Sulla nuova popolazione generata vengono
ripetuti i passi precedenti. I risultati sono stati ottenuti con numero di generazioni pari a 2000.
Il problema numerico
La soluzione numerica del modello del sistema cardiovascolare fetale di Figura 2 è stata
ottenuta mediante implementazione in linguaggio MATLAB delle equazioni del modello stesso,
ricavate a partire dalla topologia del circuito applicando i metodi classici della teoria dei circuiti,
nonché dell’AG. È stato considerato un ciclo cardiaco di 0,345 secondi – come misurato in (9) –,
suddiviso in 250 incrementi temporali. Il programma ha calcolato, per ciascuno step temporale,
le portate di sangue attraverso resistenze e capacità, i volumi immagazzinati nei vari letti
vascolari e nei ventricoli e le pressioni in tutti i nodi della rete cardiovascolare. Al fine di
assicurare l’ottenimento di una soluzione numerica stazionaria, il programma ha eseguito calcoli
per una durata temporale corrispondente a 20 cicli cardiaci. Tale programma veniva chiamato
dalla routine che implementava l’AG quando quest’ultima doveva eseguire la valutazione, in
modo che potesse fornire i dati per calcolare la funzione di fitness. Dopo 2000 generazioni
create dalla routine che implementava l’AG, è stato identificato il miglior set (in base alle
valutazioni fornite dalla funzione di fitness) di parametri circuitali e di parametri delle funzioni
elastanza con la corrispondente soluzione numerica ottenuta per il modello.
18
RISULTATI
Il primo risultato rilevante ottenuto consiste nella identificazione, effettuata mediante la
applicazione degli AG, della topologia circuitale ipotizzata. Il comportamento fisiologico
dell’istmo aortico è stato infatti riprodotto soltanto in seguito all’applicazione degli AG al
modello matematico rappresentato topologicamente in Figura 2; si è riusciti ad ottenere ciò
soltanto dopo aver inserito nel modello la circolazione polmonare, non essendo altre topologie
più semplici testate in grado di riprodurre numericamente le condizioni fisiologiche.
In Tabella 5 sono riassunti i valori che costituiscono il miglior set di parametri circuitali e di
parametri delle funzioni elastanza identificati.
Tabella 5. Parametri del modello: miglior set identificato
Parametro
R1
R2
R3
R4
R5
R6
R7
R8
R9
Valore stimato
0,01 mmHg*s/ml
1,01 mmHg*s/ml
2,94 mmHg*s/ml
0,68 mmHg*s/ml
3,91 mmHg*s/ml
4,57 mmHg*s/ml
0,33 mmHg*s/ml
0,33 mmHg*s/ml
0,67 mmHg*s/ml
Parametro
R10
R11
R12
R13
C1
C2
C3
C4
C5
Valore stimato
20,01 mmHg*s/ml
0,93 mmHg*s/ml
0,28 mmHg*s/ml
1,37 mmHg*s/ml
0,66 ml/mmHg
0,54 ml/mmHg
21,99 ml/mmHg
0,27 ml/mmHg
0,59 ml/mmHg
Parametro
Valore stimato
C6
Emax (VD)
V0 (VD)
Emax (VS)
V0 (VS)
P0 (VD)
K0 (VD)
P0 (VS)
K0 (VS)
28,04 ml/mmHg
17,40 mmHg/ml
-2,01 ml
13,18 mmHg/ml
-1,59 ml
0,03 mmHg
0,87 ml-1
0,14 mmHg
0,42 ml-1
VD: ventricolo destro
VS: ventricolo sinistro
La Tabella 6 mostra un confronto fra i valori di pressione e portata calcolati nei ventricoli e
nei vari letti vascolari, insieme con i valori di stroke volume generati dal modello funzionale e i
corrispondenti valori misurati in vivo (raccolti in letteratura e misurati dagli autori).
Con il modello caratterizzato dal set di parametri identificato, è stato calcolato un valore di
pressione massima nel ventricolo sinistro pari a 65,24 mmHg, che risulta essere molto vicino al
corrispondente valore di 68 mmHg (9), nonché un valore di pressione massima nel ventricolo
destro pari a 83,56 mmHg; anche tale valore calcolato mostra un buon accordo con la
corrispondente quantità (80 mmHg), misurata da Rudolph (12).
Il valore calcolato di stroke volume eiettato dal ventricolo sinistro è risultato pari a 1,91 ml,
mentre per il ventricolo destro è stato calcolato un valore pari a 2,83 ml (rispettivamente, circa il
40% e il 60% della gittata cardiaca combinata). I valori fisiologici di stroke volume che si
desiderava riprodurre (vedi Tabella 4, contenente i dati utilizzati per costruire la funzione di
fitness) sono pari a 1,7 ml per il ventricolo sinistro, misurato da Grigioni et al. (9), e 3,2 ml per
il ventricolo destro (valore stimato a partire dallo stroke volume eiettato dal ventricolo sinistro,
misurato da Grigioni et al. (9), e dalla distribuzione della gittata cardiaca combinata che,
utilizzando i dati riportati da Rudolph (12, 28), è stata utilizzata per la costruzione della
funzione di fitness (vedi Tabella 4), valori che rappresentano rispettivamente circa il 35% e il
65% della gittata cardiaca combinata, in accordo con quanto riportato da Rudolph (28). A tal
riguardo, è possibile notare come i valori calcolati dal modello costituiscano una leggera
sottostima dello stroke volume ventricolare destro e una leggera sovrastima di quello del
ventricolo sinistro.
19
Tabella 6. Confronto fra grandezze fisiologiche e grandezze misurate dal modello
Parametro
Valore fisiologico
Rif.
Risultato modello
Pressione massima VS
68 mmHg
(9)
65,24 mmHg
Volume Eiettato VS (VEVS)
1,7 ml
(9)
1,91 ml
Distribuzione della Gittata
20÷22% parte superiore del corpo (28)
Cardiaca Combinata (GCC)
(testa, collo, cervello)
2÷4% circolazione coronarica
∼10% attraverso l’istmo aortico
verso l’aorta discendente
55÷60% attraverso il dotto arterioso
verso l’aorta discendente
5÷10% polmoni
29,65% parte superiore del
corpo (testa, collo, cervello) +
circolazione coronarica
10,65% attraverso l’istmo aortico
verso l’aorta discendente
53,47% attraverso il dotto
arterioso verso l’aorta
discendente
6,23% polmoni
Ritorno sanguigno venoso
65÷70% dalla VCI
∼8% dalle vene polmonari
22÷25% dalla VCS
(28)
74,87% da VCI
+ vene polmonari
25,13% dalla VCS
Pressione massima VD
80 mmHg
(28)
83,56 mmHg
Volume Eiettato VD (VEVD) 3,2 ml
*
2,83 ml
Distribuzione
del volume di sangue
13% rete arteriosa
7% rete capillare
75% rete venosa
5% cuore
(31)
(23)
11,86% rete arteriosa
9,77% rete capillare
76,31% rete venosa
2,06% cuore
Volume di sangue
105 ml/kg
(30)
103 ml/kg
Rif. Riferimento bibliografico
* Valore stimato a partire dallo stroke volume eiettato dal ventricolo sinistro [Grigioni et al. (9), e dalla distribuzione
della gittata cardiaca combinata [Rudolph (12, 28)].
VD: ventricolo destro; VS: ventricolo sinistro
La Figura 11 mostra i diagrammi pressione-volume, calcolati su un ciclo cardiaco, del
ventricolo destro e del ventricolo sinistro; nella medesima figura è riportato il diagramma
sperimentale pressione-volume del ventricolo sinistro, relativamente alla fase sistolica, così
come misurato mediante cateterizzazione (9) in condizioni fisiologiche: dal confronto fra i
diagrammi pressione-volume calcolato e misurato per il ventricolo sinistro si osserva come il
modello sottostimi leggermente il volume ventricolare sinistro di fine diastole, ma non quello di
fine sistole. La gittata cardiaca combinata calcolata è risultata distribuita per circa il 60% in
aorta discendente (diretta verso la parte inferiore del corpo, che comprende placenta, fegato,
reni), mentre il 10,65% ha attraversato l’istmo aortico e il 53,47% il dotto arterioso. La parte
superiore del corpo (che comprende testa, collo, cervello), insieme con la rete vascolare
coronarica ha ricevuto il 29,65%, mentre il rimanente 6,23% della gittata cardiaca combinata è
stato distribuito ai polmoni. I valori calcolati mostrano un buon accordo con i range fisiologici
relativi alla distribuzione percentuale del sangue, così come misurata da Rudolph et al (28) e
riassunta in Tabella 5. Il valore calcolato di volemia del feto è risultato essere di 103 mlkg-1 , che
mostra un ottimo accordo con il valore di 105 mlkg-1 misurato in (30). Il volume di sangue
calcolato si è distribuito per l’11,86% nella rete arteriosa, per il 9,77% nella rete capillare, per il
76,31% nella rete venosa, mentre il rimanente 2,06% è stato distribuito al cuore. Anche questi
valori calcolati si mostrano in buon accordo con i corrispondenti dati, calcolati e sperimentali
(23, 31), riassunti in Tabella 6 (ad eccezione del cuore, 2,06% il valore calcolato, contro lo
sperimentale 5%: ma se si considera che il modello non contempla la circolazione coronarica,
allora anche per il sangue distribuito al cuore si ottiene un buon accordo).
20
Pressione (mmHg)
VENTRICOLO DESTRO
CALCOLATO
VENTRICOLO SINISTRO
MISURATO
VENTRICOLO SINISTRO
CALCOLATO
Volume (ml)
Figura 11. Diagrammi pressione-volume
Pressione (mmHg)
Sulla base delle ipotesi fatte nel modellizzare le resistenze di ingresso dei ventricoli, la
distribuzione percentuale calcolata del ritorno venoso è risultata così ripartita: il 25,13%
proveniente dalla circolazione ‘superiore’ (ritorno dalla VCS); il 74,87% proveniente da
circolazione ‘inferiore’ e polmonare. Dai valori riportatati in Tabella 5, si nota come la
distribuzione percentuale calcolata del ritorno venoso si mostri in ottimo accordo con i valori
sperimentali forniti da Rudolph et al (12).
Le Figure 12, 13, 14, 15, 16 mostrano, rispettivamente, le forme d’onda calcolate (attraverso le
capacità) della pressione intraventricolare destra, della pressione in aorta discendente (pressione
prossimale), della pressione arteriosa nella parte ‘inferiore’ del corpo, della pressione capillare
nella parte ‘inferiore’ del corpo e della pressione venosa nella parte ‘inferiore’ del corpo.
Tempo (ms)
Figura 12. Pressione ventricolo destro
21
Pressione (mmHg)
Tempi (ms)
Pressione (mmHg)
Figura 13. Pressione aorta discendente
Tempi (ms)
Pressione (mmHg)
Figura 14. Pressione letto arterioso inferiore
Tempi (ms)
Figura 15. Pressione letto capillare inferiore
22
Pressione (mmHg)
Tempi (ms)
Figura 16. Pressione letto venoso inferiore
Le Figure 17, 18, 19, 20 mostrano, rispettivamente, le forme d’onda calcolate della portata
eiettata dal ventricolo destro, della portata in aorta discendente, della portata nel dotto arterioso
e della portata diretta verso il circolo polmonare.
Volume (ml)
Flusso (ml/s)
flusso
(ml/s)
volume
(ml)
Tempi (ms)
Figura 17. Flusso e volume ventricolo destro
23
Flusso (ml/s)
Tempi (ms)
Flusso (ml/s)
Figura 18. Flusso in aorta discendente
Tempi (ms)
Flusso (ml/s)
Figura 19. Flusso nel dotto arterioso
Tempi (ms)
Figura 20. Flusso in arteria polmonare
24
Flusso (ml/s)
Tempi (ms)
Figura 20. Flusso in arteria polmonare
Le Figure 21, 22, 23, 24, 25, mostrano, rispettivamente, le forme d’onda calcolate (attraverso
le capacità) della pressione intraventricolare sinistra, della pressione in aorta ascendente
(pressione distale), della pressione arteriosa nella parte ‘superiore’ del corpo, della pressione
capillare nella parte ‘superiore’ del corpo e della pressione venosa nella parte ‘superiore’ del
corpo.
In Figura 21 viene mostrata anche la forma d’onda della pressione ventricolare sinistra
misurata mediante cateterizzazione dagli autori (9).
pressione
calcolata
Pressione (mmHg)
pressione
misurata
Tempi (ms)
Figura 21. Pressione ventricolo sinistro (calcolata e misurata)
25
Pressione (mmHg)
Tempi (ms)
Pressione (mmHg)
Figura 22. Pressione aorta ascendente
Tempi (ms)
Pressione (mmHg)
Figura 23. Pressione letto arterioso superiore
Tempi (ms)
Figura 24. Pressione letto capillare superiore
26
Pressione (mmHg)
Tempi (ms)
Figura 25. Pressione letto venoso superiore
Le Figure 26, 27, 28 mostrano, rispettivamente, le forme d’onda calcolate della portata
eiettata dal ventricolo sinistro, della portata in aorta ascendente e della portata nell’istmo
aortico.
Le Figure dalla 11 alla 28 mostrano come il modello numerico, con il set di parametri
individuato, sia in grado di calcolare forme d’onda delle grandezze emodinamiche che risultano
essere molto simili alle corrispondenti a quelle fisiologiche. Una conferma di ciò la si può
ottenere dal confronto fra la forma d’onda della pressione ventricolare sinistra calcolata e quella
misurata (Figura 21), nonché dal confronto fra i diagrammi pressione-volume, calcolato e
misurato, relativi al ventricolo sinistro (Figura 11).
volume
(ml)
Tempi (ms)
Figura 26. Flusso e volume ventricolo sinistro
27
Flusso (ml/s)
Volume (ml)
flusso
(ml/s)
Flusso (ml/s)
Tempi (ms)
Flusso (ml/s)
Figura 27. Flusso aorta ascendente
Tempi (ms)
Figura 28. Flusso istmo aortico
28
DISCUSSIONE
Il principale obiettivo del presente studio è la costruzione di un modello matematico del
sistema cardiovascolare fetale, in grado di riprodurre le forme d’onda delle grandezze
emodinamiche nei differenti distretti vascolari. Nella definizione del modello, è stata posta una
particolare attenzione nel considerare le caratteristiche uniche della circolazione fetale,
rappresentate nello specifico dal dotto arterioso, dal setto interatriale e dalla presenza dell’istmo
aortico; quest’ultimo costituisce un importante sito vascolare, poiché unisce circolazione fetale
‘superiore’ e ‘inferiore’ consentendo ad una parte del sangue eiettato dal ventricolo sinistro di
fluire in aorta discendente (Figura 1). La identificazione del set di parametri del modello è stata
effettuata mediante la applicazione degli AG. Gli AG sono poco onerosi dal punto di vista
computazionale e non risultano limitati da ipotesi restrittive sullo spazio di ricerca (ovvero
continuità, derivabilità, ecc.). Inoltre essi non hanno bisogno di un gran numero di informazioni
ausiliarie per lavorare propriamente. L’utilizzo degli AG ha consentito di individuare, fra le
infinite soluzioni del problema, il set di parametri (Tabella 5) più plausible per rappresentatività,
mediante la applicazione di una funzione di fitness costruita in modo tale da rappresentare
l’errore quadratico medio calcolato con riferimento ai valori riportati in Tabella 3.
La topologia circuitale proposta (Figura 2) è riuscita a riprodurre con successo, in seguito
alla fase di identificazione dei parametri basata sull’uso degli AG, il comportamento fisiologico
dell’istmo aortico soltanto quando è stata esplicitamente modellizzata la circolazione polmonare
(i polmoni nel feto ricevono il 5-10% della gittata cardiaca combinata). Sono state infatti
studiate topologie più semplici, ma questi modelli semplificati non si sono rivelati capaci di
riprodurre numericamente le condizioni fisiologiche.
La funzione cardiaca è stata caratterizzata descrivendo la relazione fra pressione e volume
ventricolari tramite il concetto di elastanza (26). Dalla equazioni [1] e [4] appare evidente come
un approccio di questo genere sia imprescindibile da una accurata identificazione dei parametri
(Emax, V0 ), e (P 0 , k0 ). Come notato da Ménigault et al. (22), un infinito numero di tali coppie di
parametri risulta compatibile con i valori fisiologici di pressione, volume e portata. Tuttavia, in
base ai dati sperimentali disponibili in letteratura riguardanti la stima dell’elastanza di feti di
pecora, è possibile caratterizzare propriamente il ventricolo sinistro mediante i valori Emax=12
mmHg/ml, V0 =-4 ml (29). In passato questi valori sono stati utilizzati come riferimento anche
per il ventricolo destro (23), avendo assunto per le due camere ventricolari un uguale
funzionamento. In questo studio non è stato imposto alcun valore a ciascuna delle due coppie
(Emax, V0 ), ma è stato semplicemente fissato un range di valori ammissibili per tali parametri
(necessario all’AG per la identificazione).; inoltre la stima dei due valori di Emax è stata eseguita
in maniera indipendente per ciascuno dei due ventricoli.
È noto che il cuore di feto lavora molto vicino ai limiti superiori della sua capacità
funzionale (32), con il ventricolo destro sufficientemente forte da soddisfare la domanda
fisiologica postnatale. Inoltre alla gittata cardiaca combinata i due ventricoli contribuiscono per
una percentuale di circa il 34% per quanto riguarda il ventricolo sinistro e del 66% per quanto
riguarda il destro. Sulla base di quanto testè affermato, l’ipotesi di uguali valori di E max per i due
ventricoli sembrerebbe discutibile. I valori delle coppie (E max, V0 ) ottenuti al termine della fase
di identificazione appartengono al range fisiologico (13,18 mmHg/ml, -1,59 ml per il ventricolo
sinistro; 17,40 mmHg/ml, -2,01 ml per il ventricolo destro). Per quanto riguarda il ventricolo
sinistro, il valore di E max stimato presenta un ottimo accordo con quello determinato
sperimentalmente (12 mmHg) (29); per quanto riguarda il ventricolo destro la mancanza di dati
sperimentali relativi all’E max impedisce di poter eseguire un confronto con il valore stimato.
29
Il set di parametri del modello individuato tramite gli AG ha consentito di calcolare forme
d’onda di pressione e portata con morfologia molto simile a quella delle grandezze
emodinamiche misurate sperimentalmente (come mostrano le Figure 11-28); tale buon accordo
tra grandezze calcolate e misurate risulta particolarmente evidente confrontando (Figura 11) i
diagrammi pressione-volume, quello calcolato con quello misurato da Grigioni et al. (9), relativi
al ventricolo sinistro (nel confronto, occorre considerare che la misura sperimentale, a causa del
non perfetto accoppiamento fra le dimensioni del catetere e del ventricolo sinistro, potrebbe
essere affetta da polarizzazione), nonché confrontando (Figura 21) le forme d’onda della
pressione ventricolare sinistra, calcolata e misurata da Grigioni et al. (9).
Nel modello, il comportamento dei due ventricoli è duale: mentre per il ventricolo sinistro la
pressione massima calcolata risulta sottostimata e il volume eiettato sovrastimato rispetto ai
corrispondenti dati sperimentali (rispettivamente 65,24 contro 68 mmHg e 1,91 contro 1.7 ml,
come riportato in Tabella 6), per il ventricolo destro si ottengono ripettivamente una sovrastima
e una sottostima per la pressione massima e il volume eiettato (83,56 contro 80 mmHg e 2,83
contro 3,2 ml, come riportato in Tabella 6).
Le Figure 13, 14 mostrano come la pressione in aorta discendente (pressione prossimale) sia
molto simile a quella della rete arteriosa inferiore. In maniera similare, le Figure 22, 23
mostrano come la pressione nel tratto iniziale dell’aorta ascendente sia più elevata della
pressione arteriosa della parte superiore del corpo. La pressione capillare inferiore risulta più
elevata di quella capillare superiore (Figure 15, 24), mentre la pressione venosa risulta la
medesima sia per la parte inferiore che per quella superiore (Figure 16, 25). Pertanto, le
maggiori perdite di carico vengono calcolate dal modello nella parte inferiore del corpo, tenendo
ivi conto in maniera corretta della presenza della placenta.
Uno studio numerico del sistema cardiovascolare del feto umano è stato presentato da
Menigault et al. (23): in esso gli autori hanno rappresentato la rete vascolare mediante un
modello di Windkessel a sei elementi e il cuore fetale è stato considerato comportarsi come un
singolo ventricolo equivalente (assumendo che i due ventricoli lavorano in parallelo); in tale
studio (23), i valori delle capacità che costituiscono il modello sono stati calcolati a partire da
dati presenti in letteratura, sotto l’ipotesi di laminarità del flusso sanguigno. Nel modello
oggetto di questo studio, al contrario, tali parametri sono stati stimati mediante gli AG, senza
fare alcuna ipotesi sul tipo di flusso. La somma delle capacità dei letti arteriosi inferiore e
superiore C1 e C 4 (0,93 ml/mmHg) risulta molto simile al valore della capacità del letto arterioso
trovato da Menigault et al. (0,9 ml/mmHg) (23). Il medesimo accordo è stato trovato per quanto
riguarda le capacità capillari (1,13 contro 0,75 ml/mmHg) e venose (50,03 contro 51
ml/mmHg).
Nel sistema vascolare dell’adulto, la compliance venosa è in massima parte rappresentata
dalla rete di piccole vene e venule costituenti appunto il letto venoso (33). Nel feto di pecora, gli
elevati valori di pressione sistemica media misurati (34, 35) sembrano confermare quanto
accade per l’adulto, suggerendo che nel feto la frazione maggiore di compliance venosa sia
allocata nella regione in cui la pressione venosa è più elevata, ossia più in prossimità del letto
capillare piuttosto che in prossimità dell’atrio. Nel modello qui presentato, il ruolo compliante
di questi distretti vascolari è interpretato dai parametri capacitivi C6 (regione superiore) e C3
(regione inferiore) della Figura 2: come mostrato in Tabella 5, tali valori capacitivi sono i più
elevati tra quelli del set dei parametri del modello, in accordo con (33-35).
Un confronto fra pressioni e volumi medi calcolati nel nostro studio e i corrispondenti dati
presenti in letteratura è reso assai difficoltoso dal fatto che questi ultimi si riferiscono quasi
esclusivamente o a singoli organi oppure a regioni vascolari nella loro globalità (ovvero rete
arteriosa, capillare o venosa in toto), mentre in questo studio tali regioni sono state considerate
non come un uno, ma separate in un’area inferiore e una superiore.
30
Gli autori ritengono importante sottolineare come il modello elaborato calcoli un valore di
volemia fetale (103 ml) in ottimo accordo con i dati sperimentali (105 ml) (30), senza aver fatto
alcuna ipotesi di partenza sul valore della volemia stessa. L’aspetto peculiare del modello è la
presenza dell’istmo aortico, elemento di connessione fra la circolazione fetale superiore e
inferiore; la scelta di modellizzare tale segmento vascolare mediante un elemento resistivo ha
consentito di ottenere la distribuzione di flussi attesa.
Il modello di sistema cardiovascolare fetale oggetto del presente studio può rappresentare il
passo preliminare nella direzione della realizzazione di uno strumento che risulti utile per
investigare i potenziali feedback che potrebbero aver luogo fra i vari distretti vascolari a causa
di una qualche alterazione avvenuta in qualcuno di essi; tale modello potrebbe risultare utile
anche nella interpretazione funzionale di alcuni fenomeni fisiologici osservati di azionereazione, nonché nella simulazione di procedure cliniche come i bypass cardiaci fetali. Il
modello potrebbe essere utilizzato come strumento per identificare dati insufficienti o non
esistenti e per studiare e interpretare un gran numero di esperimenti di fisiologia fetale: ciò
potrebbe rivelarsi utile per la conoscenza del ruolo e dell’importanza che i singoli parametri
hanno nel determinare la emodinamica fetale.
31
BIBLIOGRAFIA
1. Hanley FL. Fetal cardiac Surgery. Adv Card Surg 1994;4:47-74.
2. Sakata M, Hisano K, Okada M, Yasufuku M. A new artificial placenta with a centrifugal pump: longterm total extrauterine support of goat fetuses. J Thorac Cardiovasc Surg 1998;115:1023-31.
3. Reddy VM, Liddicoat JR, Klein JR, McElhinney DB, Wampler RK, Hanley FL. Fetal cardiac bypass
using an in-line axial flow pump to minimize extracorporeal surface and avoid priming volume. Ann
Thorac Surg 1996;62:393-400.
4. Reddy VM, Liddicoat JR, Klein JR, Wampler RK, Hanley FL. Long-term fetal outcome after fetal
cardiac bypass: fetal survival to full term and organ abnormalities. J Thorac Cardiovasc Surg
1996;111:536-44.
5. Hanley FL. Inhibition of fetal stress response improves cardiac output and gas exchange after fetal
cardiac bypass. J Thorac Cardiovasc Surg 1994;107:1416-22.
6. Assad RS, Lee FY, Bergner K, Hanley FL. Extracorporeal circulation in the isolated in situ lamb
placenta: hemodynamic characteristics. J Appl Physiol 1992;72:2176-80.
7. Fenton KN, Heinemann MK, Klautz RJM, Liddicoat JR, Hanley FL. Inhibition of fetal stress
response improves cardiac output and gas exchange after fetal cardiac bypass. J Thorac Cardiovasc
Surg 1994;107:1416-22.
8. Vedrinne C, Tronc F, Martinot S, Robin J, Garhib C, Ninet J, Lehot JJ, Franck M, Champsaur G.
Effects of various flow types on maternal hemodynamics during fetal bypass: is there nitric oxide
release during pulsatile perfusion?. J Thorac Cardiovasc Surg 1998;116(3):432-9.
9. Grigioni M, Carotti A, Daniele C, D’Avenio G, Morbiducci U, Iannace E, Albanese S B, Costa D,
Formigari R, Ferretti E, Di Donato R M. Extracorporeal circulation in ewe’s foetus: towards a
reliable foetal cardiac surgery protocol. Int J Artif Organs 2000;23:189-98.
10. Durand A, Vieyres PP, Patat F, Descamps PH, Grègoire JM, Pourcelot D, Pourcelot L. Etude des
paramàtres hémodynamiques influençant la mesure d’index de résistance placentaire le long de
l’artàre ombilicale. JEMU 1992;13:188-94
11. Vieyres PP, Durand A, Patat F, Descamps PH, Grègoire JM, Pourcelot D, Pourcelot L. Influence of
the measurement location on the resistance index in the umbilical arteries: a haemodynamic
approach. J Ultrasound Med 1991;10:671-5.
12. Rudolph AM. Distribution and regulation of blood flow in the fetal and neonatal lamb. Circ Res
1985;57(6):811-21.
13. Tranquart F, Collt M, Arbeille P H. Hémodynamique aortique cérébrale et rénale foetale. In:
Pourcelot L (Ed.). Dynamique cardio-vasculaire foetale et néonatale Echographie-Doppler. Paris:
Masson; 1991. p. 51-66.
14. Huikeshoven F J, Coleman T G, Jonsma H W. Mathematical model of fetal cardiovascular system:
the uncontrolled case. Am J Physiol 1980;239:317-25.
15. Veth AFL, van Bemmel JH. The role of placental vascular bed in the fetal response to cord
occlusion. In: Longo LD, Reneau DD (Ed.). Fetal and newborn cardiovascular physiology. New
York: Garland; 1978. p. 579-604.
16. Gibbons DT, Johnson F, Wright JTM. ‘A model of the fetal circulation during labor. In: Longo LD,
Reneau DD (Ed.). Fetal and newborn cardiovascular physiology. New York: Garland; 1978. p.60514.
17. Allen WW, Power GG, Longo LD. Fetal O2 changes in response to hypoxic stress: a mathematical
model. J Appl Physiol: Respirat Environ Exercise Physiol 1977;42:179-90.
32
18. Allen WW, Power GG, Longo LD. Fetal oxygen responses to hypoxia: a mathematical model. In:
Longo LD, Reneau DD (Ed.). Fetal and newborn cardiovascular physiology. New York: Garland;
1978. p. 551-60.
19. Cameron JM, Reneau DD, Guilbeau EJ. Multicomponent analysis of the fetal system. In: Longo LD,
Reneau DD (Ed.). Fetal and newborn cardiovascular physiology. New York: Garland; 1978. p. 497550.
20. Huikeshoven FJ, Hope ID, Power G, Gilbert RD, Longo L. Mathematical model of fetal circulation
and oxygen delivery. Am J Physiol 1985;249:192-202.
21. Pennati G, Bellotti M, Fumero R. Mathematical modelling of the human foetal cardiovascular system
based on Doppler ultrasound data. Med Eng Phys 1997;19:327-35.
22. Ménigault E, Vieyres P, Lepoivre B, Berson M, Durand A, Pourcelot D, Tranquart F, Pourcelot L.
Modelisation hemodynamique du coeur foetal humain: etude physiologique. In: Societé de
Biomécanique, Actes du 20éme Congrés.
23. Ménigault E, Vieyres P, Lepoivre B, Durand A, Pourcelot L, Berson M. Fetal heart modelling based
on a pressure-volume relationship. Med Biol Eng Comput 1997;35:715-21.
24. Goldberg E. Genetic algorithms in search, optimization and machine learning. New York: Addison
Wesley, 1989.
25. Poli R. Introduction to evolutionary computation. Notes from the lessons. Birmingham: School of
Computer Science, University of Birmingham; 1996.
26. Suga H, Sagawa K, Shoukas AA. Load independence of the instantaneous pressure-volume ratio of
the canine left ventricle and effects of epinephrine and heart rate on the ratio. Circ Res 1973;32:31421.
27. Frank O. Zur Dynamic des Herzmuskels. Ztschr Biol 1885;14:370-447.
28. Rudolph AM. Congenital diseases of the heart. Chicago: Year Book Medical Publisher Inc;1974.
29. Kirkpatrick SE, Friedman WF. Myocardial determinants of fetal cardiac output. In: Longo LD,
Reneau DD (Ed.). Fetal and newborn cardiovascular physiology. New York: Garland; 1978. p. 36984.
30. Dubois O, Fievet P, Berquin JL. La variabilité des données biologiques di nouveau-né au cours des
premières vingtquatre heures. In: Dubois O, Renaud E (Ed.). Médecine périnatale 4ème journée
nationale. Tours 1974. p. 261-94.
31. Kayser C. Physiologie: les grandes fonctions. Paris: Editions Medicales Flammarion;1963.
32. Rudolph AM, Heyman MA. The circulation of the fetus in utero: methods for studying distribution of
blood flow, cardiac output and organ blood flow. Circ Res 1967;21:163-84.
33. Green JF. Determinants of systemic blood flow. In: Guyton AC, Young DB (Ed.). Cardiovascular
physiology III. Vol. 18. Baltimore (MD): University Park Press; 1979. p. 33-65.
34. Gilbert RD. Determinants of venous return in fetal lamb. Gynecol Invest 1977;8:233-45.
35. Gilbert R D. Control of fetal cardiac output during changes in blood volume. Am J Physiol (Heart
Circ Physiol 7) 1980;238:H80-6.
33
Presidente dell’Istituto Superiore di Sanità
e Direttore responsabile: Enrico Garaci
Coordinamento redazionale:
Paola De Castro e Sandra Salinetti
Stampato dal Servizio per le attività editoriali
dell’Istituto Superiore di Sanità, Viale Regina Elena, 299 - 00161 ROMA
La riproduzione parziale o totale dei Rapporti e Congressi ISTISAN
deve essere preventivamente autorizzata.
Reg. Stampa - Tribunale di Roma n. 131/88 del 1° marzo 1988
Roma, marzo 2002 (n. 1) 3° Suppl.
La responsabilità dei dati scientifici e tecnici
pubblicati nei Rapporti e Congressi ISTISAN è dei singoli autori