Il campo magnetico terrestre
La terra come un dipolo magnetico
Nord
geografico
S
N
Nord
magnetico
Posizione del Polo Nord magnetico
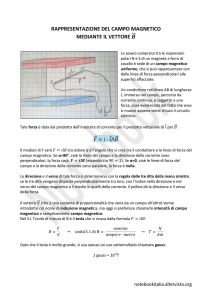
Il vettore campo magnetico
Vettore
magnetico
Il vettore campo magnetico
Vettore
magnetico
Il vettore campo magnetico
Piano de mN
ll’orizzon
te
mN
Componente
orizzontale
Inclinazione
Componente
verticale
Vettore
magnetico
Il vettore campo magnetico
gN
Piano de
ll’orizzon
te
mN
Declinazione
Componente
orizzontale
gE
Inclinazione
Componente
verticale
Vettore
magnetico
Il vettore campo magnetico
Emisfero magnetico Nord
Il vettore
• è inclinato verso il basso
• la componente orizzontale
punta verso il mN
• la componente orizzontale
punta verso il polo
Emisfero magnetico Sud
mN
mN
Il vettore
• è inclinato verso l’alto
• la componente orizzontale
punta verso il mN
• la componente orizzontale
punta verso l’equatore
Il campo magnetico terrestre (CMT): variazioni
Intensità:
30 - 65 µT (30 000-65 000 nT)
Inclinazione:
0° (equatore) - 90° (poli magnetici)
Variazione CMT di
5-10 nT/km e 0.01°/km
Variazioni temporali (giornaliere)
di 50-100 nT e 0.1°
(fino a 150-200 nT e 0.5-1°)
e spaziali (anomalie)
fino a oltre 100,000 nT
(spesso 5-10 % del campo locale)
Il campo magnetico terrestre (CMT): variazioni
Intensità:
30 - 65 µT (30 000-65 000 nT)
E’ possibile identificare
differenti località geografiche
sulla base dei loro parametri
magnetici (ad es. intensità e
inclinazione).
Possibile impiego di stimoli
magnetici per navigare.
Problemi per presenza di
variazioni temporali.
Inclinazione:
0° (equatore) - 90° (poli magnetici)
Variazione CMT di
5-10 nT/km e 0.01°/km
Variazioni temporali (giornaliere)
di 50-100 nT e 0.1°
(fino a 150-200 nT e 0.5-1°)
e spaziali (anomalie)
fino a oltre 100,000 nT
(spesso 5-10 % del campo locale)
La magnetorecezione
Esperimenti di condizionamento magnetico
Tentati ripetutamente, spesso con scarso successo
Buoni risultati nelle api (sensibilità di pochi nT), e (in parte)
in alcuni pesci.
Recenti risultati interessanti nei colombi e in altri uccelli.
Esperimenti di condizionamento magnetico
Colombo viaggiatore Columba livia
(ca. 100 000 nT)
Mora et al. 2004
Esperimenti di condizionamento magnetico
Colombo viaggiatore Columba livia
(ca. 100 000 nT)
Mora et al. 2004
Esperimenti di condizionamento magnetico
Pettirosso
(Erithacus rubecula)
(ca. 85 000 nT)
Kishkinev et al. 2012
Esperimenti di condizionamento magnetico
Pettirosso
(Erithacus rubecula)
Suono + CM
(ca. 85 000 nT)
Kishkinev et al. 2012
Suono
decrescente
+ CM
Solo CM
Esperimenti di orientamento magnetico: metodi
Applicazione di magneti
Albatros urlatore Diomedea exulans
Tartaruga verde Chelonia mydas
Esperimenti di orientamento magnetico: metodi
Uso di bobine magnetiche
Tartaruga liuto
(Dermochelys coriacea)
1.3 m
Lohmann 1991
Esperimenti di orientamento magnetico: metodi
Uso di bobine magnetiche
4.1 m
Tartaruga verde Chelonia mydas
Lohmann et al. 2004
Esperimenti di orientamento magnetico
Tartaruga liuto
(Dermochelys coriacea)
Lohmann 1991
Esperimenti di orientamento magnetico
Tartaruga liuto
(Dermochelys coriacea)
Lohmann 1991
Esperimenti di orientamento magnetico
Pettirosso
(Erithacus rubecula)
Campo magnetico locale
Wiltschko & Wiltschko 1999
Campo magnetico
ruotato di 120°
Il recettore magnetico
e i sistemi di trasduzione
Tre principali meccanismi di trasduzione degli stimoli
magnetici (magnetorecezione) proposti:
1. Magnetorecezione per induzione
2. Magnetorecezione chimica (tramite fotopigmenti)
3. Magnetorecezione tramite particelle ferromagnetiche
(magnetite)
Magnetorecezione per induzione
Informazioni magnetiche percepite da elettrorecettori che
rilevano il campo elettrico indotto dal movimento
dell’animale in un campo magnetico esterno
Il campo elettrico indotto dipende dalla direzione ed intensità
del campo magnetico attraversato e dal rapporto spaziale
tra la direzione di movimento e quella del campo
magnetico terrestre (zero per movimenti allineati con linee
di forza magnetiche, è massimo a 90° dalle esse).
La risposta rilevata dagli elettrorecettori dipende quindi dalla
direzione di movimento: sistema adatto per percepire
direzioni (magnetiche) di movimento (prestazioni di
bussola).
Possibile solo in acqua salata, complicato dalla presenza di
campi elettrici disturbanti prodotti da flussi di acqua non
derivanti da moto (correnti).
Magnetorecezione per induzione: evidenze
Presenza di elettrorecettori specifici nelle ampolle del
Lorenzini negli Elasmobranchi
Magnetorecezione per induzione: evidenze
Presenza di elettrorecettori specifici nelle ampolle del
Lorenzini negli Elasmobranchi
Kalmijn 1978
Magnetorecezione per induzione: evidenze
Presenza di elettrorecettori specifici nelle ampolle del
Lorenzini negli Elasmobranchi
Questi apparati rilevatori di campi elettrici, che servono per
rilevare prede (in base al campo elettrico prodotto),
misurano gradienti di voltaggio in senso dorso- ventrale,
quindi sul piano verticale.
Kalmijn 1978
Magnetorecezione per induzione: evidenze
Presenza di elettrorecettori specifici nelle ampolle del
Lorenzini negli Elasmobranchi
Questi apparati rilevatori di campi elettrici, che servono per
rilevare prede (in base al campo elettrico prodotto),
misurano gradienti di voltaggio in senso dorso- ventrale,
quindi sul piano verticale.
Le ampolle hanno sensibilità elevate (anche 0.010 µV/cm),
che sono tali da poter rilevare correnti indotte anche da
movimenti non particolarmente veloci (pochi m/s).
Scarse prove sperimentali dell’uso di questi elettrorecettori
per rilevare informazioni magnetiche direzionali: in
acquario rilevata capacità di condizionamento magnetico
e di assunzioni di specifiche direzioni nello spazio, ma
interpretazione dei dati controversa e non univoca.
Magnetorecezione chimica
Possibilità che il decorso e i prodotti di reazioni chimiche a
carico di (macro)molecole biologiche possano dipendere
da campi deboli come il CMT, e che questo determini
(almeno) una prima trasduzione dello stimolo magnetico.
Situazione particolarmente adatta nelle reazioni che
coinvolgono la formazione di radicali liberi: Radical Pair
Mechanism.
Magnetorecezione chimica:
il Radical Pair Mechanism
Campi magnetici
(anche deboli)
Trasduzione dello stimolo magnetico (poi amplificata)
Ritz et al. 2000
Magnetorecezione chimica:
il Radical Pair Mechanism
Campi
magnetici
Ritz et al. 2010
Magnetorecezione chimica:
il Radical Pair Mechanism nei fotorecettori
Reazioni adatte per un meccanismo siffatto devono possedere
determinate caratteristiche (range specifico di durata,
molecole con pochi ioni idrogeno, ecc..).
Importanza della disposizione ordinata e rigida delle molecole
interessate all’interno delle cellule e delle cellule stesse,
per garantire la massima efficienza del sistema (anche se
il processo può funzionare anche con recettori disposti
casualmente).
Queste condizioni sono tipicamente realizzate nelle reazioni a
seguito dell’assorbimento di luce, per cui soprattutto nei
fotorecettori retinici.
Magnetorecezione chimica:
il Radical Pair Mechanism nei fotorecettori
Fotoeccitazione
Campi magnetici
(anche deboli)
Trasduzione dello stimolo magnetico (poi amplificata)
Ritz et al. 2000
Magnetorecezione chimica:
il Radical Pair Mechanism nei fotorecettori
Ipotesi di magnetorecezione a carico di fotopigmenti eccitati
proposta più volte (Hong, Leask, Edmonds); attualmente il
meccanismo più accreditato è quello elaborato da Ritz e
collaboratori (Univ. Illinois, USA).
Proposto ruolo dei criptocromi, molecole fotorecettoriali note
nei Mammiferi, le cui proprietà sarebbero ottimali per
fornire informazioni direzionali (bussolari) tramite RPM.
I prodotti delle reazioni con i radicali nei fotopigmenti
differirebbero in differenti parti della retina, producendo
così differenti pattern visivi al variare dell’orientamento
dell’animale (“vision-based magnetic compass”).
Questi fenomeni sarebbero poi processati dalle normali vie
sensoriali visive.
Magnetorecezione chimica:
il Radical Pair Mechanism nei fotorecettori
Campi geomagnetico
in Illinois (USA)
50 µT; incl. – 68°
Ritz et al. 2000
Evidenze di magnetorecezione tramite fotorecettori
Esperimenti di orientamento con luci a differenti lunghezze d’onda
Esperimenti di orientamento magnetico
Pettirosso
(Erithacus rubecula)
Campo magnetico locale
Wiltschko & Wiltschko 1999
Campo magnetico
ruotato di 120°
Evidenze di magnetorecezione tramite fotorecettori
Effetti di luci a differenti lunghezze d’onda sull’orientamento dei Passeriformi migratori
Visible light
Pettirosso
(Erithacus rubecula)
Wiltschko et al. 2010
Evidenze di magnetorecezione tramite fotorecettori
Effetto dell’intensità della luce sull’orientamento dei Passeriformi migratori
Test a differenti intensità di luce (misurate in quanti/m2/s)
Pettirosso
(Erithacus rubecula)
Luce verde a 565 nm
Wiltschko & Wiltschko 2005
Evidenze di magnetorecezione tramite fotorecettori
Orientamento dei pettirossi al buio
Luce verde
Pettirosso
(Erithacus rubecula)
Buio
Stapput et al. 2008
Evidenze di magnetorecezione tramite fotorecettori
Casi di corretto orientamento magnetico al buio
Tartaruga liuto
(Dermochelys coriacea)
Lohmann 1991
Evidenze di magnetorecezione tramite fotorecettori
Effetti di campi oscillanti a radio frequenze sull’orientamento dei Passeriformi migratori
Campo RF a 1.315 MHz (in luce verde a 565 nm)
Pettirosso
(Erithacus rubecula)
Thalau et al. 2005
Evidenze di magnetorecezione tramite fotorecettori
Ruolo dei Criptocromi
I criptocromi sono una classe di fotopigmenti nel range blu-verde che
possono formare radicali liberi dopo fotoeccitazione (le rodopsine non
lo fanno)
I criptocromi sono quindi le uniche molecole fotoeccitabili presenti nella
retina dei Vertebrati che possono soddisfare i requisiti previsti dal
RPM.
Tramite tecniche di biologia molecolare l’espressione dei geni per i
criptocromi (4 tipi) è stata rilevata nella retina del beccafico, nel gallo
domestico e nel pettirosso. Sono espressi soprattutto in coni sensibili
al luci violette o ultraviolette, localizzati vicino ai dischi di membrana.
Ruolo dei criptocromi nella magnetorecezione
Orientamento magnetico notturno
Beccafico
(Sylvia borin)
Espressione dei criptocromi nella retina
Strato nucleare esterno
Strato plexiforme esterno
Strato nucleare interno
Displaced ganglion cells
Strato plexiforme interno
Strato cellule gangliari
Mouritsen et al. 2004
Ruolo dei criptocromi nella magnetorecezione
Displaced ganglion cells
Strati nella retina dei Vertebrati
Ruolo dei criptocromi nella magnetorecezione
Ruolo dei Criptocromi
Strato nucleare esterno
Strato plexiforme esterno
Strato nucleare interno
Displaced ganglion cells
Strato plexiforme interno
Strato cellule gangliari
Mouritsen et al. 2004
Ruolo dei criptocromi nella magnetorecezione
Ruolo dei Criptocromi
Strato nucleare esterno
Strato plexiforme esterno
Strato nucleare interno
Displaced ganglion cells
Strato plexiforme interno
Strato cellule gangliari
Strato nucleare esterno
Strato plexiforme esterno
Strato nucleare interno
Strato plexiforme interno
Strato cellule gangliari
Mouritsen et al. 2004
Ruolo dei criptocromi nella magnetorecezione
Displaced ganglion cells
Mouritsen et al. 2004
Evidenze di magnetorecezione tramite fotorecettori
Ruolo dei Criptocromi
I criptocromi sono una classe di fotopigmenti nel range blu-verde che
sembrano poter formare radicali liberi dopo fotoeccitazione (le
rodopsine non lo fanno)
I criptocromi sono quindi le uniche molecole fotoeccitabili presenti nella
retina dei Vertebrati che possono soddisfare i requisiti previsti dal
RPM.
Tramite tecniche di biologia molecolare l’espressione dei geni per i
criptocromi (4 tipi) è stata rilevata nella retina del beccafico, nel gallo
domestico e nel pettirosso. Sono espressi soprattutto in coni sensibili al
luci violette o ultraviolette, localizzati vicino ai dischi di membrana.
Nel beccafico i prodotti di questi geni sono espressi durante
l’orientamento magnetico notturno e sono localizzati nel citosol
delle cellule gangliari retiniche.
Le cellule gangliari retiniche (dislocate o meno) che esprimono i criptocromi
nei migratori sono quelle attive di notte, durante l’orientamento
magnetico migratorio.
Evidenze di magnetorecezione tramite fotorecettori
Ruolo del Cluster N
Le cellule gangliari retiniche sono in connessione con
un’area telencefalica (Cluster N, parte del Wulst
visivo), che si attiva durante la ‘visione notturna’.
Mouritsen et al. 2005; Heyers et al. 2007
Evidenze di magnetorecezione tramite fotorecettori
Ruolo del Cluster N
Pettirosso
(Erithacus rubecula)
Zapka et al. 2009
Evidenze di magnetorecezione tramite fotorecettori
Ruolo dei Criptocromi
Drosophila
melanogaster
Gegear et al. 2008
Evidenze di magnetorecezione tramite fotorecettori
Ruolo dei Criptocromi
Dopo training
Risposta spontanea
Drosophila
melanogaster
Gegear et al. 2008
Evidenze di magnetorecezione tramite fotorecettori
Ruolo dei Criptocromi
Dopo training
Risposta spontanea
Drosophila
melanogaster
Gegear et al. 2008
Evidenze di magnetorecezione tramite fotorecettori
Ruolo dei Criptocromi
Drosophila
melanogaster
Wild-type
Gegear et al. 2008
Omozigoti
(Cry inattivo)
Eterozigoti
(Cry attivo)
Magnetorecezione tramite particelle ferromagnetiche
Magnetorecezione tramite particelle ferromagnetiche
Ruolo della magnetite (Fe3O4) biogenica
• Magnetite a dominio singolo (magneti permanenti)
cristalli di 50-70 nm, spesso in catene
possono ruotare ed allinearsi con i campi esterni
Batterio magnetotattico con catene di
magnetite a dominio singolo.
Catene di magnetite a dominio singolo
della regione etmoidale di un salmone.
Magnetorecezione tramite particelle ferromagnetiche
Ruolo della magnetite (Fe3O4) biogenica
• Magnetite a dominio singolo (magneti permanenti)
cristalli di 50-70 nm, spesso in catene
possono ruotare ed allinearsi con i campi esterni
• Magnetite superparamagnetica (senza momento permanente)
particelle più piccole
si possono allineare con i campi esterni senza che la particella
debba muoversi
Campo assente
Magnetorecezione tramite particelle ferromagnetiche
Ipotetici meccanismi di trasduzione tramite magnetite biogenica
Tramite catene di magnetite a dominio singolo
Canali ionici
Canali ionici
Campo magnetico
Walker et al. 2002
Magnetorecezione tramite particelle ferromagnetiche
Ipotetici meccanismi di trasduzione tramite magnetite biogenica
Tramite catene di magnetite a dominio singolo
Filamenti del citoscheletro
Canali ionici
Campo magnetico
Walker 2008
Magnetorecezione tramite particelle ferromagnetiche
Ipotetici meccanismi di trasduzione tramite magnetite biogenica
Tramite clusters di magnetite superparamagnetica
Canali ionici
Canali ionici
B=0
Clusters di magnetite
Clusters di magnetite
Campo magnetico
Davila et al. 2003
Magnetorecezione tramite particelle ferromagnetiche
Questi modelli (puramente congetturali) hanno
due caratteristiche in comune:
1) essendo formati da elementi subcellulari, questi
trasduttori magnetici possono essere anche minuscoli
e quindi difficili da localizzare ("truly, a magnetic
needle in the haystack“ – Kirschvink et al. 2002)
2) questi sistemi possono percepire variazioni
(potenzialmente anche minute) nella direzione e/o
nell'intensità del CMT. Entrambi i tipi di magnetite
sembrano quindi essere in grado di rilevare non solo la
direzione del CMT (prestazioni di bussola magnetica)
ma anche variazioni spaziali del campo stesso
(prestazioni di mappa magnetica).
Localizzazione della magnetite biogenica
Magnetite a dominio singolo nelle lamelle olfattive della trota iridea
Cellula
con magnetite
(a dominio singolo)
Lamelle olfattive
Trota iridea
Oncorynchus mykiss
ros V
(branca del trigemino)
Epitelio
olfattivo
Capsula
nasale
Lamina
basale
Nervo olfattivo
Walker et al. 1997; Diebel et al. 2000
Localizzazione della magnetite biogenica
Magnetite superparamagnetica nel colombo
Colombo viaggiatore
Columba livia
Membrana
Aggregato
di magnetite
‘Piastrine’
Cristalli di magnetite
superparamagnetica
Fleissner et al. 2003, 2007
Localizzazione della magnetite biogenica
Magnetite superparamagnetica nel colombo
Colombo viaggiatore
Columba livia
Modello dell’aspetto della
terminazione nervosa
(branca oftalmica del
trigemino)
‘Piastrine’
Aggregati
Fleissner et al. 2003, 2007
Localizzazione della magnetite biogenica
Magnetite superparamagnetica nel colombo
Colombo viaggiatore
Columba livia
Orientamento preferenziale dei dendriti
• Simile disposizione rilevata anche nel pettirosso, nel beccafico e persino
nel gallo domestico
Fleissner et al. 2007
Localizzazione della magnetite biogenica
Magnetite superparamagnetica nel colombo
Colombo viaggiatore
Columba livia
Treiber et al. 2012
“Here we show that clusters of iron-rich cells in the rostro-medial
upper beak of the pigeon Columbia livia are macrophages, not
Orientamento preferenzialemagnetosensitive
dei dendriti
neurons.
Our systematic characterization of the pigeon upper beak identified
• Simile disposizione rilevata anche nel pettirosso, nel beccafico e persino
iron-rich cells in the stratum laxum of the subepidermis, the basal
nel gallo domestico
region of the respiratory epithelium and the apex of feather follicles.”
Fleissner et al. 2007
Localizzazione della magnetite biogenica
Magnetite superparamagnetica nel colombo?
Colombo viaggiatore
Columba livia
Treiber et al. 2012
Esperimenti di “Pulse magnetization”
Magnetic pulse: intensità 0.5 T (500,000,000 nT), durata 3-5 ms
Australian Silvereys
Zosterops lateralis
Esperimenti di “Pulse magnetization”
Magnetic pulse: intensità 0.5 T (500,000,000 nT), durata 3-5 ms
Australian Silvereys
Zosterops lateralis
Controlli
Pulsed
Munro et al. 1997
Esperimenti di “Pulse magnetization”
Magnetic pulse:
intensità 0.04 T (40,000,000 nT),
durata 4 ms
Tartaruga comune
Caretta caretta
Irwin & Lohmann 2005
Controlli
Pulsed
Esperimenti di “Pulse magnetization”
Magnetic pulse: intensità 0.1 T (100,000,000 nT), durata 0.1 ms
Rilevazione orientamento tramite radio-tracking VHF
Controlli
Pulsed (perpendicular)
Pettirosso
(Erithacus rubecula)
Cannaiola
(Acrocephalus scirpaceus)
Holland 2010
Esperimenti di condizionamento magnetico
Colombo viaggiatore Columba livia
(ca. 100 000 nT)
Mora et al. 2004
Ruolo della branca oftalmica del nervo trigemino
Esperimenti di condizionamento magnetico
Sezione della branca
oftalmica del trigemino
Colombo viaggiatore
Columba livia
Mora et al. 2004
Ruolo della branca oftalmica del nervo trigemino
Esperimenti di condizionamento magnetico
Abolizione della capacità
discriminativa degli
stimoli magnetici dopo
sezione della branca
oftalmica
Colombo viaggiatore
Columba livia
Mora et al. 2004
Ruolo della branca oftalmica del nervo trigemino
Esperimenti di navigazione nel colombo
Sezione della branca oftalmica
del trigemino
Ruolo della branca oftalmica del nervo trigemino
Esperimenti di navigazione nel colombo: orientamento iniziale
Colombo viaggiatore
Columba livia
Falsi operati
Gagliardo et al. 2009
Resezione della
branca oftalmica
Ruolo della branca oftalmica del nervo trigemino
Esperimenti di navigazione nel colombo: prestazioni di homing
Kruskal-Wallis P<0.002
Gagliardo et al. 2009
Ruolo della branca oftalmica del nervo trigemino
Esperimenti nel pettirosso
Pettirosso
Erithacus rubecula
Zapka et al. 2009
Ruolo della branca oftalmica del nervo trigemino
Esperimenti nel pettirosso
Pettirosso
Erithacus rubecula
Zapka et al. 2009
Evidenze di magnetorecezione tramite fotorecettori
Ruolo del Cluster N
Pettirosso
(Erithacus rubecula)
Zapka et al. 2009
Magnetorecezione tramite particelle ferromagnetiche
Riassunto delle evidenze sperimentali
1) Effetto della pulse magnetization in alcuni casi (silvereyes,
passeriformi migratori, piccoli di tartaruga; non nel colombo). Ruolo
nella navigazione magnetica?
2) Indicazioni contrastanti sul ruolo della branca oftalmica del
nervo trigemino:
• Evidenze morfologiche/ultrastrutturali non confermate
• Lesioni della branca non sempre producono effetto
• Effetti derivanti dall’anestesia dell’epidermide del
becco di interpretazione non chiara
• Risultati elettrofisiologici contrastanti e/o difficili da
replicare
Meccanismi di magnetorecezione: riassunto
Meccanismo Caratteristiche –
funzionamento
Per
induzione
Informazioni
fornibili
Rilevazione dei
Direzionali,
campi elettrici
a polarità o
derivanti dal
inclinazione
movimento nel
campo magnetico
Evidenze sperimentali
Strutture
coinvolte
Presenza di
elettrorecettori
sensibili negli
Elasmoranchi
Ampolle del
Lorenzini
Chimica
Modifiche nei
tramite
prodotti di
fotorecettori reazioni chimiche
con radicali liberi
indotte da
fotoeccitazione
Direzionali,
a
inclinazione
Effetti sull’orientamento
di variazioni nella
qualità e intensità della
luce; espressione dei
criptocromi nella retina
durante l’orientamento
magnetico
Retina
(cellule
gangliari),
vie visive
(nBOR,
tetto ottico)
Tramite
particelle di
magnetite
Direzionali
(polarità o
inclinazione)
e di intensità
magnetica
Presenza di magnetite a
dominio singolo o
superparamagnetica in
varie strutture, anche
nervose; effetto dei
magnetic pulse
sull’orientamento.
Regione
etmoidale becco;
ramo
oftalmico
del
trigemino
Allineamento
delle particelle
con campi
esterni; influenza
su canali ionici
meccanosensibili