Prima lezione
Definizione Istologia: studio dei tessuti.
I tessuti si possono definire come complesse associazioni di cellule o di cellule e sostanza tra di esse. Sostanza che
viene chiamata intercellulare anche detta matrice extracellulare.
I tessuti si organizzano tra di loro e i risultati sono strutture anatomiche rivelabili come tali definite organi, che
hanno una specifica funzione. Per es. prendiamo un epitelio di rivestimento mettiamoci sotto un connettivo poi
una parte muscolare poi ancora del connettivo, integrati possono dare un organo come lo stomaco che ha come
funzione specifica quella digestiva.
Ma l’organo è ben più del complesso dei singoli tessuti così come più organi vanno a costituire un’unità
complessa, integrata da un punto di vista funzionale, che è l’apparato. Per esempio l’apparato digerente costituito
da più organi che tutti insieme collaborano a quella che la digestione e l’assimilazione. L’insieme degli apparati
danno l’organismo. L’organismo è un insieme mai una somma. L’insieme in biologia è sempre più di una somma
tra loro di vari apparati, ciascun apparato di vari organi, gli organi di vari tessuti, i tessuti di cellule.
Allora la cellula è il punto di partenza per capire i vari livelli organizzativi del mostro organismo e come tale la
prenderemo in considerazione, partendo dalla definizione di cellula: la più piccola porzione costituita di materiale
vivente capace di vita autonoma. Questo significa che la materia vivente si organizza a formare delle unità
funzionali e morfologiche che vengono chiamate cellule.
Definiamo la materia vivente partendo dalle sue caratteristiche che sono:
- La variabilità
- La capacità di adattamento
- La riproduzione
(la materia vivente non si presenta sempre uguale, ha insito nelle proprie strutture, nel proprio significato,
appunto la variabilità, che noi tocchiamo e vediamo a livello cellulare nella grande variabilità che esiste tra una
cellula e l’altra sebbene tutte costituite su uno stesso schema. Ma la cellula del fegato è uguale a un neurone,
non è identica. E’ sempre una cellula, è sempre
quell’elemento ma non è uguale. Saliamo, il
tessuto epiteliale dello stomaco riveste lo stomaco come riveste l’epidermide di un corpo ma non sono uguali
identici, sono diversi . Saliamo la scala, per cui un organismo è simile al suo vicino ma non identico a meno che
non esistano due gemelli monovulari. Ecco la grande variabilità che parte dalla materia vivente come substrato
su
cui si costituiscono i vari livelli organizzativi. Ed è importante che esista questa variabilità a
tutti i livelli perché la comparsa periodica di variazioni sotto forma di mutazioni ha reso possibile quella che
Darwin ha descritto come evoluzione. In atto tutt’ora, sempre come la selezione di caratteri più adatti
all’ambiente.
Capacità di adattamento all’ambiente: La materia vivente non è inerte, non è semplice aggregato di materiale
biologico, risponde attraverso l’irritabilità. Se io sollecito in qualche maniera la materia vivente questa risponde
per diventare adatta a quello stimolo. Pensate alla cellula nervosa in cui l’irritabilità è espressa al massimo grado,
tale a trasmettere stimoli che corrono sulla sua membrana, che sono tutte quelle informazioni che ci rendono atti
a vivere insieme agli stimoli del sistema endocrino. Ma anche la cellula è in grado di rispondere nel suo piccolo
in maniera adatta allo stimolo che gli viene applicato, tutte le cellule.
Poi questa capacità di adattamento si esprime attraverso la capacità di assorbimento. Noi non siamo in grado di
fabbricarci materiale organico, non siamo piante verdi, quindi a livello cellulare e di organismo assimiliamo e
assorbiamo sostanze che poi elaboriamo attraverso meccanismi metabolici, e questo ci rende in gradi di
accrescerci. Oltre ad accrescerci perché
sintetizziamo, siamo anche in grado di motilità inteso come
movimento, contrazione, cellulare (ameboide), ma anche come spostamento di organuli all’interno di una cellula.
Quindi a tutti i livelli è sempre valida questa affermazione.
Inoltre la riproduzione: La materia vivente si crea da ciò che rea già la materia vivente, niente nasce dal nulla,
la cellula nasce dalla cellula madre, un individuo nasce dalla fecondazione dei gameti della madre e del padre.
Questa materia vivente si organizza in livelli strutturali diversi, organizzazione in cellule procariotiche ed
eucaristiche (livello più complesso).
Le cellule eucaristiche oltre ad avere un organizzazione strutturale molto più elevata di quelle procariotiche
(tipica dei batteri), è anche in grado di una specializzazione (nella pluricellularità le cellule si indirizzano verso
direzioni diverse per portare avanti funzioni diverse che daranno vita all’organismo si specializzano acquisendo
caratteristiche morfologiche biochimiche particolari. Poi si aggregano formano i tessuti fino ad arrivare
all’organismo pluricellulare. Se noi guardiamo una cellula eucaristica, quando si aggrega ad un’altra, fa colonia,
dove ciascuno continua a portare avanti le proprie funzioni. E’ raro che ci sia un’integrazione del tipo di cui
parliamo noi, anche se è semplice. D’ora in poi noi parleremo delle caratteristiche della cellula eucaristica.
La materia vivente prende il nome di protoplasma che è costituito da componenti inorganici e organici.
I componenti inorganici sono di due tipi: l’acqua e i Sali minerali. L’acqua è essenziale per la vita (si dice che
dove non c’è acqua non c’è vita, ed è vero). L’individuo maschile ne ha il 70%, l’individuo femminile un po’ di
meno perché ha più grasso. Alcuni tessuti innalzano questa percentuale, e quindi la dobbiamo pensare come un
elemento essenziale per la vita. L’acqua è un solvente e le molecole possono avere due tipi di comportamento:
interagire (polari, idrofile) o non avere questo tipo di capacità (non polari, idrofobe). Poi noi in istologia ce ne
dimentichiamo perché studiamo la struttura, e l’acqua non si vede, anzi l’istologo spesso la porta via per potere
osservare i tessuti. Ma voi non ve ne potete scordare anche perché è nell’acqua che avvengono tutte quelle
reazioni che hanno a che fare con il metabolismo cellulare, tessutale e poi dell’organismo.
I Sali minerali sono un’altra componente importante in organica della materia vivente. I componenti organici sono
tutte e quattro le categorie dei composti che voi conoscete:
- carboidrati
- lipidi
- proteine (insieme di amminoacidi caratterizzate da una struttura primaria che identificano la proteina, che può
dare una sequenza amminocidica secondaria, terziaria, quaternari ecc..)
- Acidi nucleici (sapere una loro descrizione generalizzata vedi: proteine)
LA CELLULA EUCARIOTICA
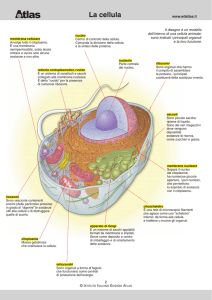
E’ costituita dalla membrana plasmatica che la avvolge, la separa e la connette con l’ambiente, e un altro sistema
membranoso che si chiama involucro nucleare che separa il contenuto della cellula in due distretti. Uno esterno (il
citoplasma) uno interno (il nucleo). Dobbiamo conoscere tutte queste parti a livello strutturale e funzionale.
Apriamo una parentesi: l’occhio umano ha una risoluzione di circa 0.1/0.2 millimetri, questo significa che
un’occhio riesce a distinguere due punti a 0.1 mm per un occhio d’aquila. Sotto questo limite non si focalizza non
si distinguono. Poiché le dimensioni medie delle cellule sono nell’ordine di 20-25 micrometri e poiché l’istologo
studia le cellule e poi i tessuti, va da sé che a occhio nudo non vede nulla. Ecco l’urgenza di introdurre strumenti
d’indagine chiamati microscopi che sono in grado di alzare questo potere e quindi di farci vedere due punti che
sono più vicini di 0.1 mm.
Il microscopio ottico è uno strumento che può stare sul tavolo e che porta il potere di risoluzione a circa 0.2
micrometri (nel caso di un buon microscopio ottico). Permette di osservare cellule, tessuti, strutture cellulari fino
al limite di 0.2 micrometri.
Il microscopio così detto elettronico permette di osservare strutture fino a 2 nanometri. Le strutture subcellulari
sono quasi tutte visibili con il microscopio elettronico a trasmissione. E’ uno strumento grande che entra in una
piccola stanza davanti al quale sta seduto l’operatore. E usa un pennello di elettroni che corrono lungo un tubo
sottovuoto, è una struttura complessa paragonata ad uno strumento ottico valido che utilizza una sorgente di
illuminazione quale la lampadina. Questa è microscopia elettrica a trasmissione detta TME.
Di tutti microscopi elettronici che usano il pennello elettronico, c’è il SEM (microscopio a scansione), che ha
potere di risoluzione minore perché ci fa vedere strutture fino a 20 nm. Ci permette di vedere la superficie delle
strutture dando un’immagine che sembra tridimensionale.
Quindi due metodiche di indagine che danno informazioni complementari.
Parliamo di immagini e di risoluzioni di strutture per cui si può usare un analizzatore senza dover vedere.
Per vedere al microscopio io non posso prendere una biopsia mentre quello viene operato perché ho delle
difficoltà derivanti dal fatto che il tessuto separato dall’organismo muore andando incontro a deterioramento
Quindi l’istologo provoca la morte immediatamente dopo che il pezzetto di tessuto è stato asportato, lo fissa,
rendendo l’immagine il più vicina possibile a quella che aveva nel vivente di solito denaturando le proteine
rendendole insolubili, allontano l’acqua e tutti i processi enzimatici vengono bloccati quindi anche la
rigenerazione. Poi ho bisogno di ridurlo in sottilissime sezioni, ma poiché è materiale biologico estremamente
variabile, si ha poco contrasto tra le strutture, quindi coloro per la microscopia ottica, cioè conferisco dei colori
che sono sempre delle ricette che rendono le colorazioni ripetitive e per il microscopio elettronico a trasmissione
spolvero dei metalli pesanti che interagendo con gli elettroni gli danno immagini nere (quindi dico elettropache o
elettrodense) mentre le porzioni prive di questi metalli lasciano passare gli elettroni e quindi sono bianche
(elettrotrasparente).Trasmissione osservo strutture subcellulari, scansione osservo la superficie.
Entrando nei particolari si perde in ampiezza generale.
Le dimensioni cellulari: medie sono circa 20-25 micrometri. Ce ne sono anche di più piccoli tipo 3, 3.5
micrometri come il piccolo linfocita, lo spermatozoo, il globulo rosso (7.2).Anche qui grande variabilità, per la
maggior parte cellule medie, in parte piccole, poi ci sono cellule grandi come la cellula adiposa o come i neuroni
che arrivano 60-70 micrometri fino a che superato il limite dei 150 micrometri si parla di cellule giganti, esempio
su tutti la cellula uovo (il gamete femminile). Se si superano i 200 micrometri si parla di cellule particolari quali
quelle che ritroviamo nel tessuto muscolare che, in certi casi, come ad esempio le cellule lisce dell’utero durante
la gravidanza, possono arrivare ad alcuni mm o addirittura cm, allora le vedo anche ad occhio nudo.
Anche la forma delle cellule è variabile. I fisici ci dicono che se la cellula è libera di prendere le dimensioni che
più gli aggradano, per fenomeni di tensione superficiale in cui noi non entriamo, sarebbe una sfera. Di fatto in
natura poche cellule sono sferiche, ce ne sono alcune che fanno parte del pool dei globuli bianchi come il
granulocita e il monocita, che però rotolano dentro un sistema fluido. A livello dei tessuti possiamo trovare la
cellula adiposa che è globosa per un grosso accumulo di trigliceridi dentro, poi la cellula uovo che per
antonomasia è una cellula rotonda.
Ma la forma cellulare si modifica per due motivi:
Mutui contatti:se una cellula deve aderire strettamente ad un’altra a formare delle fibre come negli epiteliali e
negli agglomerati, due palle si toccano in un solo punto, mentre i prismi sono adatti quindi selezionati per avere
contatti più stretti. La forma più poliedrica nelle sue più svariate manifestazioni (cubico, cilindrico,
parallelepipedo, tronco di piramide ecc..). Forma poliedrica sfaccettata nella più grande variabilità.
L’altra spinta è la funzione. La cellula si specializza durante la sua vita. Nasce secondo modello base, poi per
stimoli ambientali ed espressione del proprio genoma, acquisisce caratteristiche morfologiche diverse in base alla
funzione.
La prima caratteristica che può assumere è la forma in base alla funzione. La cellula nervosa, il neurone, è una
cellula grande avvolte gigante che si è selezionata per ricevere informazioni dalla periferia e portarle ai centri per
elaborare risposte dai centri e portarle anche in periferia oppure per fare circuiti. Allora si costituisce questa forma
costituita da un corpo con dei prolungamenti, e uno di questi è in grado di raggiungere distretti lontani dal corpo e
avere contatto con tantissime altre cellule uguali. Quindi esprime in forma una funzione acquisita. Lo
spermatozoo si è adattato a risalire apparato femminile e raggiungere la cellula uovo. Ha la forma ridotta ad una
testa che porta il nucleo, dei lisosomi e tanti mitocondri per avere energia, più un lungo flagello. Terzo
esempio:gli elementi muscolari. Che cos’è la contrazione?è un meccanismo complesso in cui l’elemento in
questione si accorcia a parità di forme quindi diventa più grosso. Sono selezionate strutture molto allungate che
accorciandosi possono gonfiare rendendo la contrazione efficace, a differenza se fossero dei palloni.
Membrana plasmatica:
E’ una sottile pellicola che separa e unisce la cellula dall’ambiente. Anche gli storici con microscopi rudimentali
si erano accorti che c’era una pellicola a dare individualità alla cellula perché, se la bucavano con microaghi,
usciva del materiale. Negli anni ’50 con l’avvento della microscopia elettronica riuscimmo a mettere in evidenza
la membrana con tre strati, due eletropachi (uno che guarda all’ambiente extra e l’altro intracellulare) con nel
mezzo uno strato elettrotrasparente. Lo spessore era tra 0.7 e 10 nm. I chimici si misero all’opera scoprendo i
costituenti ma smontando le strutture. I costituenti sono proteine, lipidi e carboidrati. I lipidi fanno parte
integrante della membrana e sono posti in doppio strato. Le proteine possono essere parte integrante della
membrana oppure no e cmq ne fanno parte in due aspetti diversi. I carboidrati sono posizionati nella porzione
esterna della membrana verso l’ambiente extracellulare, a formare il glicocalice.
I fisiologi si erano dati da fare per spiegare il funzionamento della membrana, mancava un modello che mettesse
tutti d’accordo. Tra gli anni ’50 e ‘60 sono venute fuori tante idee, fino al ’71 ad opera di Singer e Nicolson, che
idearono un modello. Modello che mise tutti d’accordo, ed è ancora valido, secondo il quale le membrane cellulari
sono fatte da lipidi fatti in modo che una porzione della loro molecola ha affinità con l’acqua (porzione polare o
testa), poi hanno delle code fatte da acidi grassi che non hanno affinità con l’acqua e quindi sono idrofobe, non
polari. Questi lipidi appartengono alla categoria dei fosfolipidi e colesterolo, quindi le membrane cellulari sono
costituite da fofsolipidi e in qualche parte da colesterolo, va da sé che, visto la grande variabilità tra cellule, i tipi
di fosfolipidi e quantità di colesterolo può variare. Questi lipidi se gettati in ambiente acquoso quale quello
vivente, si organizzano in un doppio strato esponendo le teste idrofile all’ambiente intra ed extra cellulare dove
c’è acqua, affondano le code idrofobe tra loro. Si crea quindi uno strato di lipidi a circondare le cellule, a doppio.
Questo triplo strato è quello che si vede in microscopia.
I chimici ci hanno detto che ci sono anche le proteine. Proteine di membrana integrali intrinseche che hanno la
molecola costituita in maniera tale da affondare nel doppio strato lipidico, alcune un po’ ,altre a tutto spessore
sporgendo sia nell’ambiente intra che extra cellulare.
Poi esistono proteine estrinseche così dette, cioè proteine che contraggono coi lipidi dei legami deboli, e si
adagiano sul versante sia interno che esterno. Il tipo di interazione delle proteine intrinseche, rispetto alle
estrinseche col doppio strato lipidico, è minore, avendo una conformazione molecolare che gli permette di
interagire con i lipidi, ma non tanto da affondarcisi dentro al doppio strato (dipende dall’aggregazione terziaria o
quaternaria e dagli amminoacidi che contengono all’interno).
Anche i carboidrati fanno parte integrante della membrana. Queste formazioni zuccherine a catena si possono
legare a lipidi di membrana formando glicolipidi. Oppure si legano alle proteine, sia intrinseche che estrinseche,
formando glicoproteine o proteoglicani (le prime differiscono dalle seconde per le catene glicidiche che sono
formate da monomeri o no e sono più o meno lunghe).
Si crea quindi una grossa asimmetria nella struttura della membrana perché la porzione carboidratica è solo sul
versante extracellulare. Potremmo dire che intorno alle cellule è presente una nuvola zuccherina data dai
carboidrati legati a lipidi o proteine che prende il nome di glicocalice (si vede al microscopio elettronico). Ma non
tutte le cellule lo esprimono allo stesso modo, può essere più o meno accentuato e costituito da diversi tipi di
zuccheri (grande variabilità).
Questo nell’insieme è il modello di membrana che prende anche il nome di mosaico.
Nel mosaico abbiamo le piastrelle che fanno il disegno (proteine), c’è la malta, una sostanza che tiene le piastrelle
adese tra loro (lipidi). Un mosaico fluido perché il doppio strato deve essere visto come un mantello oleoso che
circonda la cellula, che è più o meno fluido a seconda della quantità di colesterolo e del tipo di fosfolipidi che
avrò in quel tipo cellulare.
Quindi l’affinità della membrana è variabile da cellula a cellula. Ci sono cellule come i macrofagi che hanno tante
interazioni con l’esterno quindi hanno un membrana estremamente fluida. Ci sono invece cellule del nostro
sistema nervoso centrale che chiudono e proteggono il loro ambiente intracellulare per cui la fluidità sarà minore.
In questo fluido più o meno denso ci stanno le proteine che si possono spostare o rotolarci sopra.
Si chiama sistema membranoso citoplasmatico un insieme di vescicole, tubuli, cisterne rivestite da membrana
unitaria. La membrana unitaria non è appannaggio solo della membrana plasmatica ma anche di tutti i sistemi che
stanno dentro la cellula.
FUNZIONI DI MEMBRANA
1.La cellula viene identificata dalla sua membrana che gli dà una sua individualità e la separa
dall’ambiente esterno.
2.Garantisce l’adesione tra cellula e cellula. L’adesione è mediata principalmente dal
glicocalice. Le glicoproteine di membrana hanno spesso un dominio che riconosce l’ambiente intracellulare ma
anche l’ambiente extra dato dalla sostanza intercellulare, la riconoscono e ci si possono legare. Non è una
funzione di poco conto perché qualora la cellula perde la capacità di riconoscere il contatto con cellule simili,
autolimitarsi quindi nella divisione, quando perde la capacità di riconoscere dove si trova è già neoplasia,
colonizzano i tessuti estranei dando le metastasi.
3.Scambi tra cellule e ambiente. La membrana plasmatica, questo involucro, applica una
permeabilità selettiva. Dà un preciso ordine a ciò che passa. Attraverso la membrana può passare acqua, ioni,
piccole molecole. Oppure possono passare strutture vescicolari in un senso o nell’altro, materiale articolato o
macromolecole, va da sé che la membrana deve operare dei meccanismi di importazione ed esportazione più
complessi.
Il trasporto transmembranico:
T. passivo: quando acqua, ioni, piccole molecole o gas respiratori passano la membrana da un versante all’altro
seguendo un gradiente di concentrazione o elettrico. Vanno da dove è più concentrata a dove è meno. Il doppio
strato lipidico non è per loro una barriera, ci si sciolgono e riaffiorano dall’altra parte. Ma l’acqua dovrebbe
trovare un muro e stessa cosa vale per le piccole molecole e ioni. Ci vengono in aiuto le proteine intrinseche di
membrana, le quali si conformano a formare canali idrofili attraverso la membrana. Alle volte ci possono essere
proteine che riconoscono le macromolecole da far passare, ci si legano e sempre secondo gradiente le fanno
passare fuori o dentro (proteine vettrici).Riassumendo, secondo gradiente, ciò che si scioglie nei lipidi ci passa da
solo.Ciò che trova barriera trova aiuto nelle proteine sotto forma di canali idrofili o sottoforma di proteine
vettrici.
T. attivo: Viene trasportato contro gradiente di concentrazione o elettrici. Si va da dove è meno a dove è più
concentrato, quindi ci vuole energia. Qual è la molecola che immagazzina energia?ATP. Quindi c’è consumo di
ATP.Il trasporto attivo è quindi sempre legato ad attività ATP-asica, in cui c’è scissione dell’ATP in ATP+ più
fosfato (??), creando energia. Il trasporto attivo è mediato dalle proteine di membrana che scindono l’ATP e poi
trasportano le sostanze contro gradiente. In particolare voglio portare la vostra attenzione al trasporto contro
gradiente di ioni, fatti da particolari proteine di membrana che si chiamano pompe.
Le pompe altro non sono altro che proteine di membrana che trasportano ioni contro gradiente. Ci sono pompe di
vario tipo sulle membrane cellulari, per il calcio estremamente importanti per il fenomeno della contrazione. Una
moltostudiata è la pompa sodio/potassio che è legata a molti lavori cellulari ma in particolare a quel fenomeno di
membrana che è l’impulso elettrico proprio delle cellule nervose. Questa pompa trasporta ioni diversi in diverse
direzioni. Porta il sodio fuori dalla cellula contro gradiente di concentrazione stessa cosa per il potassio in
direzione opposta. Sono entrambi ioni positivi, ma mentre trasporta fuori più ioni sodio che dentro ioni potassio,
si crea una disuguaglianza di cariche positive sui due versanti della membrana, che sono alla base dei fenomeni
elettrici.
Seconda lezione
Pompe: esempio di trasporto attivo quindi contro gradiente di concentrazione. D’esempio la pompa
sodio/potassio, che si è detto essere mediata da trasporto transmembranico di ioni sodio all’esterno e ioni potassio
all’interno, entrambi contro gradiente. Sono entrambi ioni positivi, ma mentre si esporta sodio in quantità, si
importa potassio in minori quantità, si viene così a creare una differenza di cariche sui due versanti della
membrana, col versante interno meno positivo di quello esterno.
Questa differenza di potenziale elettrico, che sarà alla base di molti fenomeni di metabolismo cellulare, ma
soprattutto, a livello di quel fenomeno che va sotto il nome di corrente elettrica (ci torneremo al momento del
tessuto nervoso).
Ora ci dobbiamo occupare di trasporti transmembrana di molecole più grosse, cioè di macromolecole, o di
materiale particolato, prendendo in considerazione i due meccanismi di trasporto: o dall’esterno verso la cellula o
viceversa.
Dall’esterno verso l’interno prende il nome di endocitosi, (cioè interiorizzare queste sostanze dentro la cellula), è
un meccanismo di introduzione di materiale solido o liquido, mediante fenomeni di membrana che richiedono
energia. E’ quindi un trasporto attivo legato alla scissione dell’ATP.
Si parla di fagocitosi quando si parla di ingestione di materiale solido, di pinocitosi quando si parla di ingestione
di materiale fluido, liquido in generale. Per endocitosi si intende questa serie continua di avvenimenti a livello di
membrana. La membrana riconosce ciò che deve interiorizzare e forma una specie di goccia, una invaginazione
della sua membrana. Questa membrana coglie parte del materiale, o la particella cellulare che deve interiorizzare,
e contemporaneamente approfonda la membrana nel proprio citoplasma formando una sorta di canale , una
goccia, che tende a richiudersi al di sopra, facendo rimanere integra la soluzione della membrana e chiudendosi a
circondare il materiale ingerito.
Si ha così la potenzialità, attraverso movimenti di membrana attivi, di riconoscere, interiorizzare lasciando integra
la membrana e richiudendo, ciò che è interiorizzato, in membrane che separano materiale potenzialmente lesivo.
Si chiama esocitosi, invece, il fenomeno in direzione opposta, quindi eliminazione di materiale in ambiente
extracellulare. Si parla sempre di esocitosi legata a consumo di energia quando per esempio abbiamo il fenomeno
della secrezione cellulare. Materiale che viene prodotto dalla cellula viene espulso, e l’espulsione avviene perché
un materiale circondato da membrane si avvicina per movimenti attivi del citoplasma alla membrana
citoplasmatica, si fonde con essa, e per un fenomeno di evacuazione di ciò che era contenuto dentro la vescicola,
questa fuoriesce senza toccare il citoplasma.
Nell’eso-, come nell’endocitosi, i settori di membrana, così come il trasporto interessati, sono costantemente
riciclati, cioè devono essere sostituiti da segmenti di membrana rinnovati. Vedremo quando la cellula porta avanti
questo meccanismo più avanti.
Abbiamo visto quindi come trasporto quello transmembranico passivo (secondo gradiente), attivo (contro
gradiente). Poi abbiamo preso in considerazione il trasporto per movimento di membrana, l’eso- e l’endocitosi,
come due facce dello stesso fenomeno che coinvolge gli stessi meccanismi molecolari e citologici.
Ma la membrana continua ad avere altre funzioni:
1.
conferisce identità immunologica alla cellula:
sulla membrana cellulare esistono glicoproteine di membrana, che si chiamano antigeni di istocompatibilità.
Queste molecole glicoproteiche di membrana sono geneticamente determinate, e rendono tutte le cellule di un
individuo tutte uguali tra di loro ma diverse da altri individui, a meno che non si parli di gemelli monovulari.
Significa che il mio organismo riconosce le mie cellule grazie agli antigeni di istocompatibilità che si esprimono
su determinate cellule, perché sono geneticamente determinate ed esprimono la mia struttura intrinseca.
Attraverso questo meccanismo riconosco come non mio, quindi
no- self, ciò che è di struttura diversa
proveniente da un altro organismo, e questo scatena una risposta, che tende ad eliminare quello che non è self.
Così come si scatena una risposta quando un agente patogeno esterno si insinua nel nostro organismo. Allora,
grazie alle glicoproteine, le cellule hanno la facoltà di riconoscersi tra di loro,cioè riconoscendo ciò che è self di
un individuo da ciò che non è self. Questa caratteristica che ci rende capaci di difenderci è purtroppo alla base dei
rigetti dei trapianti di organi, per quanto compatibili per gruppo sanguigno, portano ad una differenza anche se
lieve (si cercano prima i familiari che altre persone) stimolando sempre una reazione che deve essere tenuta sotto
controllo con farmaci.
Quindi l’identità immunologica delle cellule viene conferita della loro membrana.
2.
Funzione di riconoscimento:
Anche questa è parte della membrana. Spesso ho detto che il materiale viene riconosciuto e viene esportato,
fagocitato ecc.. Si chiama sito di legame, la molecola proteica di membrana che è capace di legarsi ad altre
molecole che si trovano in ambiente extracellulare.
Possono essere molecole della sostanza extracellulare, (e l’abbiamo già visto: la membrana è capace di interagire
con ciò che sta fuori, e cellule vicine), ma può anche riconoscere ormoni ad esempio della crescita o citochine, e
queste molecole che vengono riconosciute si chiamano ligandi.Allora il sito posto sulla membrana riconosce il
ligando e si lega. Se questo legame scatena una risposta della cellula, si parla di recettore invece che sito di
legame (molecola proteica o glicoproteica capace di riconoscere una struttura extracellulare o una qualche
molecola).
Per essere più aderenti alla verità, non tutti i recettori sono di membrana, esistono recettori anche nel citoplasma,
per esempio i recettori degli ormoni steroidei, di quegli ormoni cui fanno parte per esempio tutti gli ormoni
sessuali, estrogeni, testosterone come componente della porzione corticale del surrene. In questo caso l’ormone
penetra nelle cellula attraverso la membrana, (sono di natura lipidica), e trovano il loro recettore dentro la cellula.
La reazione può portare al mutamento della forma attraverso movimenti del citoscheletro.
Quindi abbiamo visto la membrana da un punto di vista morfologico e funzionale sotto più
punti di vista..
IL CITOPLASMA
Si rivela in tutta la sua complessità col microscopio elettronico a trasmissione. Gli istologi per tenere a bada
questa grande complessità, variabilità della materia vivente, hanno un linguaggio e quindi noi descriveremo, a
livello della cellula del citoplasma, la matrice ialoplasmatica, gli organuli, il citoscheletro e gli inclusi.
La cellula eucariotica la dobbiamo interpretare, conoscere e capire come una struttura altamente integrata nelle
sue subcomponenti, e vi ho più volte detto che, l’unità, la cellula, è molto più della somma dei suoi componenti,
perché l’integrazione tra le varie parti porta a selezionare e controllare tanti meccanismi. Abbiamo visto il
trasporto per esempio, il riconoscimento, e così via.
Se noi la guardiamo da vicino questa cellula, vediamo che il nucleo è l’intelligenza della cellula, il DNA, il
materiale genetico. Non è in grado solo di selezionare i caratteri che poi verranno anche ereditati, ma è anche in
grado, attraverso la sintesi proteica, di controllare tutte quante le funzioni cellulari. Perché il metabolismo
cellulare è controllato dagli enzimi, gli enzimi sono proteine e le proteine sono codificate a livello di DNA, quindi
il DNA codifica, sceglie e fa, in modo che attraverso gli RNA messaggeri, questo fattorino, che su progetto del
DNA se ne va nel citoplasma, vengono trascritte proteine specifiche. Dove? a livello di strutture che fabbricano
quindi, e sono i ribosomi. Poi queste proteine potranno essere variamente modificate, dicosilate (?), solfatate, lo
vedremo, in altri distretti che poi le imballeranno, impacchetteranno. Poi vedremo che ci saranno distretti in cui il
materiale viene demolito perchè non serve, perché è usurato, perché è nocivo. Poi ci saranno distretti per l’energia
che servirà per tutti questi meccanismi. Poi avremo, per così dire, un confine, un portierato, dove il controllo della
membrana si esplica portando avanti la permeabilità selettiva, il riconoscimento, l’identità e così via.
Quindi un sistema integrato tenuto sotto controllo dal nucleo.
Noi partiamo a vedere quali sono le strutture cellulari dal citoplasma. Eravamo partiti col dire che era costituito da
ialoplasma, detto anche matrice citoplasmatica. La matrice è priva di struttura identificabile al microscopio, per
cui è ialina, è come un gel che è costituito in gran parte da acqua e da una serie di macromolecole, macromolecole
precursori di elementi semplici, sali, ioni, che la cellula utilizzerà per il proprio metabolismo. Nel complesso di
questo ialoplasma avvengono reazioni particolari quali la glicolisi anaerobia, ad opera di enzimi che ci sono
dislocati, e in questo ialoplasma dove avvengono tante reazioni, ci si collocano le strutture visibili come tali al
microscopio.
Quindi la prima cosa che abbiamo visto è la matrice, una specie di gel ad alto contenuto di acqua, sali, ioni,
macromolecole, enzimi particolari che porteranno avanti funzioni altrettanto particolari di cui vi renderete conto
in chimica.
Poi abbiamo un componente che si vede al microscopio e prende il nome di organulo. Gli organuli sono
compartimenti del citoplasma, per lo più membranosi ma ci sono le eccezioni, con specifica funzione. Abbiamo
detto che la cellula è un sistema integrato che porta vanti funzioni complesse, e gli strumenti per portare avanti
queste funzioni sono gli organuli, ciascuno con le proprie competenze, integrato secondo uno schema, un progetto
nucleare con le altre, capace di modificarsi e di essere regolato. Questi organuli sono pertanto strutture
obiquitarie, cioè si ritrovano in tutte le cellule, ovviamente saranno più o meno svilupparti a seconda del
differenziamento, la specializzazione che la cellula avrà fatto nel suo percorso per far parte di quell’organismo.
Facciamo un esempio: l’individuo è un insieme di apparati, tutti gli individui hanno gli stessi apparati, però se io
sono un culturista e faccio tutti i giorni ginnastica avrò molto sviluppato l’apparato scheletrico, i muscoli. Se io
sono una persona che mangia tanto e si muove poco, magari avrò visibilmente espresso l’adipe sottocutaneo.
Così la cellula, in relazione a una funzione specifica, andrà a sviluppare alcuni compartimenti rispetto ad altri, se
una cellula dovrà soltanto demolire, avrà molto sviluppato quel compartimento, se ho una cellula sintetica, avrà
soprattutto sviluppato la parte che porta avanti questo.
Quindi una domanda sugli organuli c’è per forza nel compito, che possa riprodurre tutto ciò che
abbiamo detto. (P.S.: per gli organuli sapere:cosa sono, come sono fatti, quale è la loro funzione).
Quali sono gli organuli?
ribosomi
mitocondri
lisosomi
reticolo ruvido e liscio
centrioli
l’apparato di Golgi o complesso di Golgi
I ribosomi
Sono un organulo non membranoso, cioè non è circondato da membrana, ma è costituito da granuli che appaiono
al microscopio ottico elettronopachi. Hanno dimensioni tali da non poter essere visti al microscopio ottico. Sono
formati da due sub-unità, cioè due porzioni più piccole, che nel loro insieme fanno il granulo, dato da RNA
ribosomiale complessato a proteine. La loro funzione è la sintesi proteica e solo i ribosomi portano avanti la
sintesi proteica.
Nello ialoplasma, che abbiamo detto essere il supporto in cui gli organuli stanno, troviamo due popolazioni di
ribosomi diverse. I così detti ribosomi liberi e i ribosomi adesi alle membrane.
Si parte a descrivere i ribosomi liberi, cioè quelli non fissati ad alcuna membrana. Possono ritrovarsi nel
citoplasma, sembrano i grani di un rosario, e di fatto il ribosoma è la pallina e i filo altro non è che l’RNA
messaggero che li tiene uniti e gli dà l’informazione al ribosoma su quale proteina sintetizzare. La sintesi proteica
come complesso di operazioni non è compito dell’istologia specificarlo, basta sapere che il ribosoma attraverso
informazione dell’RNA messaggero inviato dal DNA, è in grado di sintetizzare proteine specifiche.
Quali proteine? Le proteine strutturali così dette, fatta eccezione per le proteine di membrana, poi ci saranno
sintesi di proteine specifiche quali ad esempio la miosina negli elementi muscolari, dell’emoglobina nel globulo
rosso e così via. Quindi proteine specifiche di alcuni tipi cellulari, e proteine strutturali fatta eccezione per quelle
di membrana.
Ribosomi fissati alle membrane
L’insieme di ribosomi adesi alle membrane e le membrane stesse formano il reticolo endoplasmatico ruvido.
Partiamo da identificare cos’è un reticolo endoplasmatico: è un insieme di vescicole, tubuli, cisterne costituite da
membrane chiuse a struttura della membrana plasmatici. Le vescicole sono delle piccole sferule di dimensioni
variabili, i tubuli sono strutture allungate ampiamente anastomizzate tra loro, le cisterne sono delle formazioni
appiattite, schiacciate rivestite da membrana. Quando vanno a costituire il reticolo endoplasmatico ruvido, anche
detto granulare, cisterne, tubuli e vescicole ampiamente tra loro comunicanti, portano sul versante citoplasmatico
adesi i ribosomi, che danno a queste membrane un aspetto ruvido da cui il nome al reticolo endoplasmatico, cioè
dentro il citoplasma (assolutamente non rugoso).
Questo sistema costituito da membrane e da ribosomi, porta i ribosomi adesi, che su informazione dell’ RNA
messaggero, producono proteine che, invece di rilasciarle nel citoplasma come fanno i ribosomi liberi, le infilano
nelle cavità del sistema membranoso, che accoglie queste proteine sintetizzate.
Quale destino avranno queste proteine sintetizzate e poi immesse nelle vescicole, tubuli, cisterne del reticolo
ruvido? 3 destini:
potranno essere proteine di membrana (le uniche che i ribosomi liberi non fabbricano)
potranno essere proteine secretorie, quindi destinate ad essere esportate dalla cellula
potranno essere enzimi dei lisosomi cioè gli enzimi di un altro organulo.
Nella cellula oltre ad un reticolo ruvido esiste anche un reticolo endoplasmatico liscio, cioè un insieme di
vescicole, tubuli, cisterne a parete liscia, non porta adesi i ribosomi.
Questo significa che il reticolo liscio non sintetizza proteine. Allora quale è la sua funzione? In tutti i tipi di
cellule è legato alla sintesi dei lipidi, o almeno ad alcune tappe della sintesi dei lipidi. Significa che le proteine
che vanno a costituire la parete membranosa del reticolo endoplasmatico liscio, hanno la capacità di poter
sintetizzare i lipidi che vengono veicolati in questa anastomosi, rete di tubuli.
Poi il grande sviluppo del reticolo endoplasmatico liscio è spesso legato a particolari funzioni, per esempio è
legato alla produzione di ormoni steroidei, quei famosi ormoni che hanno i recettori dentro il citoplasma. Gli
ormoni steroidei sono ormoni lipidici per cui, tutte le cellule ghiandolari fatte di ormoni steroidei, dall’interstiziale
del testicolo a quella dell’ovaio che fabbricano ormoni sessuali, a tutte le cellule della corticale del surrene, hanno
un grande sviluppo del reticolo liscio. Se ci spostiamo ad esempio nel fegato la grande esplosione del reticolo
liscio è spesso legato a diverse funzioni ma in particolare ha funzione di detossificazione cellulare (il reticolo
liscio ha la capacità di liberarci attraverso delle proteine di membrana da certe molecole dannose per la nostra
salute, dividendo queste molecole in porzioni più piccole che non hanno un impatto lesivo sull’organismo, ad
esempio detossificarsi dalle droghe, inquinanti, farmaci, e così via).
Poi è legato al metabolismo del glicogeno. Il glicogeno è una forma di accumulo di zucchero intracellulare che
viene tenuto sotto controllo dal reticolo liscio. In alcuni elementi come quelli muscolari il reticolo liscio diventa
una riserva di ioni calcio. Allora il reticolo liscio è un organulo fatto da tubuli anastomizzati fatti da membrana a
parete liscia, cioè non hanno ribosomi. La funzione universale è quella di fare alcune tappe della sintesi dei lipidi
che sono parte integrante della cellula. In cellule che hanno metabolismo di un certo tipo è particolarmente
sviluppato in funzione della produzione degli ormoni steroidei, della detossificazione cellulare, al metabolismo
del glicogeno, all’accumulo di ioni calcio.
Apparato di Golgi
L’apparato complesso di Golgi, che prende nome da Camillo Golgi, ricercatore che per primo lo mise in evidenza
in cellule nervose, è un organulo crocevia di funzioni cellulari. Morfologicamente è un sistema membranoso
costituito da cisterne impilate le une sulle altre, a specifica funzione, macrovescicole che emergono dalla cima di
questa pila e microvescicole che si trovano in vicinanza della base delle cisterne. La funzione dell’apparato di
Golgi è quella di compiere la glicosilazione, cioè attaccare gruppi glicidici alle proteine, la solfatazione,
aggiungere gruppi solfatici alle proteine o alle glicoproteine facendole diventare solforati, la fosforilazione, cioè
aggiungono delle porzioni che fanno si che queste proteine possano essere modificate con l’aggiunta di gruppi
fosfati, operano i legami tra le proteine lipidiche. Sappiamo, per esempio, che le membrane, proteine e lipidi
complessati, operano la sintesi dei glicosamminoglicati (sono grossi polimeri zuccherini che dentro l’apparato di
Golgi vengono assemblati, poi può condensare questo prodotto, e lo impacchetta, cioè fornisce a questo prodotto
una membrana andando a costituire quelle che abbiamo definito macrovescicole, anche detti granuli di
secrezione).
Abbiamo più volte detto che dentro le cisterne del reticolo ruvido vengono immesse delle proteine che i ribosomi
adesi fabbricano, queste proteine, già passando nei vari distretti del reticolo ruvido, possono subire una prima
glicosilazione, cioè il primo passo per diventare glicoproteine, ma ad un certo momento il soggiorno dentro queste
cisterne/tubuli del reticolo ruvido è giunto al termine. Queste glicoproteine circondate da membrana liscia, si
avvicinano all’apparato di Golgi (la cellula è capace di spostare i propri costituenti, quindi per movimenti attivi) e
la vescicola si fonde, per un meccanismo di fusione di membrane, ai vari distretti golgiani. Non è detto che una
proteina o un complesso di proteine circondate da membrana di una vescicola debbano legarsi alle varie cisterne
del complesso golgiano, possono passarne alcune, ci scaricano dentro il loro prodotto, questo subirà uno di queste
modificazioni a seconda del destino che questa proteina dovrà avere, dopo di che emergerà dalla porzione così
detta trans del golgi,quella di maturazione, sotto forma di un prodotto che potrà essere avviato a 3 destini diversi:
o diventerà una riserva di membrana (cioè le proteine complessate ai lipidi dentro l’apparato di golgi, proteine che provenivano dal reticolo, lipidi che provenivano dal reticolo liscio, ne emergono come delle
vescicole che via via si staccano, s’attaccano alla membrana plasmatici e la vanno a rinnovare). Primo destino:
vescicole per il ricambio di membrana.
Secondo destino: Vescicole piene di enzimi per i lisosomi.
Terzo destino: Granuli di secrezione, se la cellula è ghiandolare, ma cmq una polisecrezione molte cellule
ce l’hanno, prendono la via e verranno portate fuori dalla cellula.
Riguardando la struttura vedo il nucleo che ancora non conosco ma che so che fa i progetti, gli RNA messaggeri
che vanno nel citoplasma a portare questi progetti,e incontrano i ribosomi liberi e fanno le proteine strutturali ad
eccezione di quelle di membrana, poi fanno le proteine specifiche a secondo del tipo cellulare, poi incontro i
ribosomi adesi alle cisterne e lì fabbricano determinate proteine, proteine che verranno infilate dentro le cisterne
del reticolo, queste sono o proteine di membrana perché verranno legate ai lipidi (le proteine sono sulle membrane
del reticolo liscio, i lipidi sono stati creati in questo distretto). Poi molte di queste cose che erano nate nel reticolo
ruvido passano nell’apparato di Golgi, che variamente le modifica le impacchetta e le distribuisce secondo tre
uscite: verso i lisosomi, verso tutte le membrane e verso la secrezione.
Lisosomi
Vengono definiti come formazioni vescicolari citoplasmatiche rivestite da membrana e contenente enzimi
idrolitici, che sono in grado di demolire la maggior parte dei costituenti organici. I lisosomi digeriscono in
maniera controllata materiale proveniente dall’esterno e dall’interno della cellula. Sono tutti provvisti di
membrana, ma intensamente diversificati per dimensioni e in base al contenuto. Questo è spiegabile in quanto il
lisosoma è una struttura che digerisce a forma e dimensione diversa a seconda di cosa sta demolendo.
Il lisosoma è un organulo, quindi sta dentro la cellula, digerisce roba che viene dall’esterno che dovrà essere
rimaneggiata o distrutta perché lesiva, quindi avremo il fenomeno dell’endocitosi. Per esempio capta un batterio
proveniente dall’esterno, è una struttura lesiva che viene inglobata (o la cellula viene infestata). Questa struttura
però entra dentro la cellula rivestita dalla membrana plasmatici, perché è la modalità di ingresso per l’endocitosi,
e già viene separata dal citoplasma. Questa struttura fagocitata si chiama fagosoma. Lo stesso, se sono molecole
che vengono riconosciute dalla membrana, vengono interiorizzate , si fa un endosoma. Che sia un fagosoma o un
endosoma, dipende dalla dimensione di struttura, questa circondata da membrana me la ritrovo nel citoplasma
interno, perché man mano il citoplasma per movimenti attivi l’avvicina all’apparato di Golgi. Ma se io dentro la
cellula ho degli organuli che sono vecchi, questi vanno demoliti. Ecco che il reticolo liscio, un’altra delle sue
funzioni, va vicino a questo settore di citoplasma usurato e gli fa un involucro attorno in maniera da avere il
settore di citoplasma da digerire circondato da membrana, un autofagosoma. Quindi che venga dall’esterno e sia
articolato, molecole o venga dall’interno, sempre un endosoma avrò, che sia auto o che sia etero, è lo stesso. Ora è
dentro la cellula e va digerito…come si fa? Arrivano via via come delle navette, vescicole provenienti dal Golgi
che contengono enzimi litici per distruggere le macromolecole organiche, basi, idrolasi in generale. Queste
vescicole provenienti dal Golgi si uniscono al fagosoma, qualunque origine abbia, e ci scarica dentro gli enzimi,
ecco che si forma il lisosoma vero e proprio, che è l’unione di un materiale cmq endocitato a cui viene aggiunto il
prodotto di enzimi dell’apparato di Golgi (più volte può unire vescicole a questo corpo in dipendenza da quanto è
grande e duro da digerire il materiale che è endocitato). Quindi a livello dei lisosomi ho il prodotto golgiano, il
lisosoma e gli enzimi, e il materiale che è il substrato da digerire;questi vengono in contatto, e tutto questo
avviene lasciando integro il citoplasma. Perché se questa digestione non fosse controllata da membrane, attraverso
questo meccanismo gli enzimi lederebbero la cellula con un grave danno per l’organismo. Quindi il lisosoma è
l’incontro tra gli enzimi prodotti dall’apparato di Golgi e materiale che deve essere digerito.
Che fine fa il materiale man mano che viene digerito? Può essere demolito fino a molecole semplici e queste
passano, abbiamo visto che lo sanno fare, attraverso la membrana, tale e quale al passaggio che si è visto per la
membrana plasmatica. Passano la membrana, vanno a finire nel citoplasma e lì verranno riutilizzati o passeranno
la membrana e verranno definitivamente espulsi.
Molecole semplici come ad esempio delle proteine vengono disgregate e gli amminoacidi ritornano nel pool per
rifabbricare altre proteine.
Questo nel caso in cui il lisosoma ce la faccia coi suoi enzimi a operare una digestione completa, cioè sgombra lo
stomaco, e questo lisosoma come tale sparisce. Ma se il materiale endocitato è in parte o del tutto inorganico
allora i lisosomi non lo possono digerire. Esempio: siamo nel contesto dell’apparato respiratorio. Respiro l’aria
inquinata di polveri, pulviscolo vario, scorie, carbone, queste arrivano alla porzione bassa del mio apparato
respiratorio e incontrano delle cellule che per antonomasia sono gli spazzini, i macrofagi, che riconoscono queste
particelle strane e ripuliscono gli alveoli polmonari, ingerendo e cominciano la digestione. Ma gli enzimi
lisosomiali attaccano i componenti organici, il particolato inerte non viene digerito e si accumula come corpo
residuo dentro la cellula sovraccaricandone il sistema, rendendole meno disponibili alle aggressioni ad esempio
batteriche o di altra natura, corpi residui inorganici. Corpo residui particolari si possono avere quando il lisosoma
attacca i lipidi. I lipidi sono duri a digerire, vuoi perché le lipasi sono scarse nel lisosoma, vuoi perché i lipidi man
mano che vengono aggrediti cambiano ossidandosi, dando origine a dei composti difficilmente aggredibili. Allora
rimangono anche questi corpi residui che però prendono il nome di corpo lipofuscinico. Non è infrequente nei
soggetti anziani, qualora si rendano necessarie autopsie, che i neuroni del SNC sono sovraccaricati da corpi
lipofuscinici, come esempio di materiale lipidico non completamente digerito. Si accumula in cellule come il
neurone, ad esempio, e da qui non viene espulso, e questo crea un addensamento della funzionalità della cellula.
Ultimo esempio di corpo residuo (per fare presente quanto è variegato questo mondo) ci sono dei macrofagi della
milza che riconoscono i GR vecchi e li digeriscono, ma il GR è fatto anche di emoglobina. E il gruppo N(?)
contiene ferro e il macrofago non è in grado di digerirlo. Ecco che si accumulano questi corpi. Questa volta, poi, il
ferro l’organismo lo recupera attraverso un meccanismo di trasporto al midollo.
Cosa si deve ricordare? Cos’è il lisosoma, il suo significato biologico, e che ci possono essere esempi di
incompleta digestione varia che provocano il corpo residuo, corpo a cui applico nomi diversi a seconda del
materiale ingerito.
Ci sono deficit genetici di enzimi lisosomiali diagnosticabili anche alla nascita, che possono portare, se non tenuti
sotto controllo, a malattie da accumulo lisosomiale, ecco perché mi sono tanto dilungata su il significato del corpo
residuo, perché ci possono essere esempi di carenze di idrolasi varie per cui certi substrati non vengono digeriti e
questi corpi residui si accumulano in cellule di vari distretti dell’organismo, portando a malattie, qualche volta a
morte. Ma identificandoli in maniera precoce, si possono fare diete adatte, magari non somministrando,
eliminando sostanze non digeribili dai lisosomi, e questo porta ad una riduzione molto marcata del danno (deficit
già a livello genetico).
Ridurre i lisosomi a ciò che ho detto è limitativo, perché non è vero che fanno solo la digestione intracellulare,
ma i lisosomi hanno un ruolo in importanti fenomeni che prevedono il riversarsi all’esterno della cellula che li ha
prodotti, di questi enzimi, e queste operano digestioni controllate attraverso altri meccanismi, all’esterno della
cellula.
Due esempi:
1. l’osteoblasta è una cellula dell’osso che riassorbe tessuto osseo, come fa? Attraverso la digestione lisosomiale
controllata ma esterna alla cellula, quindi è alla base di rimaneggiamenti, rimodellamenti ossei molto importanti.
2.Lo spermatozoo ha nella testa l’acrosoma (?) che è praticamente una vescicola piena di enzimi, che riversa per
ledere le membrane della cellula uovo. Il primo spermatozoo che riesce a far questo poi penetra nella cellula uovo
e la feconda, la cellula uovo che subisce l’ingresso di uno spermatozoo da origine ad una reazione che impedisce
la penetrazione di altri spermatozoi. Per cui il fenomeno della fecondazione è tenuto sotto controllo anche dagli
enzimi lisosomiali.
Questo per dire che anche il mondo lisosomiale è un mondo altamente specializzato e con più facce.
Mitocondri
Sono organuli membranosi allungati, ovalari o più o meno rotondeggianti, e qui la novità, costituiti da doppia
membrana unitaria (involucro). Una esterna liscia senza pieghe (il liscio non deve sempre rievocare l’idea dei
ribosomi o non), quella interna da origine a delle pliche più o meno allungate di forma più variabile, che prendono
il nome di creste mitocondriali, e che si affondano dentro il mitocondrio stesso, la matrice del mitocondrio.
I mitocondri sono la centralina energetica della cellula, essendo i maggiori produttori di ATP (quella molecola
fatta da diplotide (??) dell’adenina + tre gruppi fosfato di cui specialmente l’ultimo legando con il fosfato è
altamente energetico. L’enzima DPasi (?) scinde il legame, avvolte anche più di uno, dando origine, se ne togli
uno solo, ad una molecola scarto, l’ADP + gruppo fosfato.
Se noi usiamo ATP per tutti i processi cellulari (es.: motilità, contrazione, biosintesi di sostanze, trasporto attivo,
trasmissione degli impulsi) riducendolo ad ADP + gruppo fosfato noi depauperiamo la potenzialità della cellula di
portare avanti il lavoro, fino a che questo lavoro non è più capace di essere portato avanti. Ci vuole quindi qualche
organulo che ridia ATP a disposizione della cellula per il suo lavoro. Come fa a ripristinare ATP? Ce n’è una
quota parte che se ne incarica la glicolisi anaerobia a livello del citoplasma ad opera di enzimi che sono disposti
nella matrice ialoplasmatica e sono capaci di risintetizzare ATP, a partire dall’ADP + gruppo fosfato, non in
presenza di ossigeno. I mitocondri invece, e questa è la differenza, oltre che a produrre la maggior parte di ATP,
lo fanno attraverso il consumo di ossigeno, attuando quella che è una vera è propria respirazione cellulare
(consumo di ossigeno e produzione di anidride carbonica). Se vogliamo insistere sul concetto: noi abbiamo ATP
cellulare che è espressione di energia che noi introduciamo mangiando.
Attraverso l’assimilazione di prodotti a livello intestinale, noi abbiamo la capacità di produrre energia attraverso i
mitocondri. Soprattutto quest’energia verrà utilizzata per tutto il lavoro cellulare, quindi compresa la contrazione.
Ma questo lavoro cellulare ci depaupera piano piano dell’ATP dandoci molecole scariche, polarità di gruppo
fosfato, che vanno ricaricate. Nei mitocondri avviane la maggior parte di questa ricarica. Dentro i mitocondri
vanno a finire composti combustibili (circa 2 atomi di carbonio) che sono carboidrati, grassi, qualche volta anche
le proteine. Con l’ossigeno si hanno una serie di reazioni che producono energia, energia che il mitocondrio
acchiappa e riutilizza per risintetizzare ATP da ADP + gruppo fosfato, producendo anidride carbonica ed acqua
Allora i mitocondri sono organuli a doppia membrana con creste, sono responsabili del ripristino della maggior
parte dell’ATP attraverso la respirazione cellulare che avviene utilizzando prodotti di assimilazione intestinale,
ossigeno, producendo energia che verrà veicolata per riattaccare il gruppo fosfato all’ADP e si produrrà come
scarto anidride carbonica ed acqua. Il metabolismo cellulare passa anche attraverso la respirazione, infatti dentro i
mitocondri ci sono varie catene enzimatiche di cui gli enzimi del ciclo di Krebs sono nella matrice, gli enzimi
della catena respiratoria sono sopra le creste. Qui, attraverso questi due sistemi si produce energia, sistemi che ve
verranno esposti a biochimica, di cui noi ora conosciamo l’ubicazione. Questa energia verrà captata da enzimi
(particelle F1) che praticano la fosforilazione, cioè l’attacco del gruppo fosfato all’ADP, in questo processo si è
consumato O e si è prodotto CO2 e H2O.
I mitocondri sono organuli particolari, oggetto di tante teorie su cui sarebbe bello dilungarsi ma non abbiamo
tempo. Essi sono imprigionati nella cellula eucaristica, ma sono particolari perchè hanno un DNA distinto da
quello nucleare e propri ribosomi. Avendo questi 2 ingredienti sono in grado di effettuare una propria sintesi
proteica anche se modesta e di dividersi, perché con il DNA che possiedono possono operare la divisione dei
mitocondri anche al di fuori della divisione nucleare.
Questo non vuol dire che siano organuli deputati alla sintesi proteica (questa viene fatta dai ribosomi che possono
essere liberi o adesi e qualcuno sta anche dentro ai mitocondri).
Cetriolo
E’ un organulo a forma di cilindretto la cui parete è data dall’assemblarsi di piccoli tubuli cavi, microtubuli, che si
dispongono in triplette. Per farvi capire: pensate ai microtubuli come a delle cannucce, (piccoli tubuli, sottili,
cavi), si assemblano le cannucce e si fanno le triplette, ripetete l’operazione 9 volte in un complesso elettronopaco
di materiale che li ingloba e avrete fatto un centriolo. Pensatelo di una certa lunghezza, visibile al microscopio
elettronico.
Il centriolo è costituito da due strutture ravvicinate a costituire un diplosoma, due centrioli si avvicinano tra di
loro si pongono a 90°, così che se li taglio insieme ne vedo uno longitudinalmente e l’altro trasversalmente, che
sono entrambi due cilindretti con la parete costituita da 9 triplette di microtubuli (in tutto: 3x9=27). Perché si parla
dei centrioli e quindi del diplosoma? Perché se noi prendiamo una cellula (citoplasma, lisosoma e centrioli), la
zona attorno a questa struttura, fa vedere chiaramente essere l’organizzatore di questi microtubuli,e che non solo
questi microtubuli vanno a costituire il centriolo del diplosoma ma vanno anche a costituire la struttura delle
ciglia, dei flagelli, e come strutture labili, faranno parte del citoscheletro cellulare e del fuso mitotico quando la
cellula si divide.
Quindi il diplosoma, struttura costituita da due centrioli, costituita a sua volta da microtubuli, è al centro
dell’organizzatore dei microtubuli, strutture che nella cellula sono impiegate per fare i centrioli, l’asse delle ciglia,
dei flagelli, una delle componenti del citoscheletro, il fuso mitotico.
Citoscheletro
Insieme di strutture citoplasmatiche di natura proteica filamentosa, costituita da tre tipi di strutture:
filamenti sottili detti anche actinici.
microtubuli di cui abbiamo appena parlato.
e filamenti intermedi, di dimensioni intermedie.
La funzione del citoscheletro: acquisizione della forma, mantenimento della forma, cambiamento della forma (per
es.: quando si ha una risposta ad uno stimolo oppure al movimento cellulare), movimento degli organuli (più volte
abbiamo detto che per movimenti attivi le vescicole si avvicinano alla membrana ecc.. all’interno della cellula).
Con la stessa valenza di un organulo perché è obiquitario, ma sfugge a quella che è stata la definizione di
organulo, abbiamo una struttura obiquitaria di natura proteica, costituita da tre categorie di elementi, strutturati in
strutture allungate che intersecandosi tra di loro danno origine ad uno scheletro della cellula (il citoscheletro) che
permette alla cellula di assumere una forma, di mantenerla o di cambiarla, di muoversi e di muovere i suoi
costituenti. Come un intelaiatura che occupa tutta quanta la cellula.
Vediamo più da vicino: i filamenti sottili sono costituiti da una proteina, l’actina (questo ci fa comodo perché
quando arriveremo al muscolare abbiamo già fatto una parte) sotto forma globulare, a pallina. Queste palline
vengono assemblate in fili, due di questi fili si arrotolano tra loro a spirale, formando la
F-actina(?), il
filamento di actina, che va a costituire una parte della trama del citoscheletro. Questo filamento di actina
interagisce con una molecola che si trova nelle immediate sue vicinanze, che si chiama miosina, e l’interazione tra
queste due strutture porta a contrazione il citoplasma perché la miosina è capace di far scivolare l’actina rispetto
ad altri filamenti in maniera da creare delle piccole contrazioni. Questo meccanismo mediato dall’actina e dalla
miosina avrà la massima espressione negli elementi muscolari, dove la contrazione è la funzione principe.
Poi abbiamo i microtubuli. Si formano in un pool di tubuline presenti nel citoplasma, proteine globulari sotto
forma diversa, a faldella. Sono strutture formate da due tubuline (dimeri, leggermente diversi tra loro) che sotto la
direzione dell’organizzatore si assemblano tra di loro in file, le file affrontate le une alle altre a formare lamine, la
lamina si riavvolge su se stessa formare una struttura cava, i mcrotubuli.
Il microtubulo, una volta che è stato fabbricato, è stabile a livello di centrioli, ciglia e flagelli, ma è una struttura
labile (cioè da una parte deposita ma dall’altra possono togliere dimeri) nel citoscheletro per esempio, e nel fuso
mitotico. Questo ci fa capire come queste strutture possano determinare l’acquisizione di una forma ma anche il
suo mutamento, lo spostamento, il movimento cellulare, attraverso questo equilibrio tra depositazione e
sottrazione di elementi dei microtubuli stessi.
I microtubuli quindi sono alla base di tutte queste funzioni che sono integrate tra di loro e determinano il
mantenimento della forma, l’ordine intracitoplasmatico, lo spostamento, lo spostamento dei cromosomi quando
avverrà la divisione, il movimento ciliare e flagellare.
A far parte del citoscheletro ci sono anche i così detti filamenti intermedi, con dimensioni intermedie tra i
filamenti sottili e spessi di actina o miosina, per questo vengono chiamati in questo modo. Strutture anch’esse di
natura proteica ma con una caratteristica: ogni cellula o popolazione cellulare ha filamenti intermedi fatti da una
proteina diversa, per esempio le cellule epiteliali hanno le proteine dei filamenti intermedi che fanno parte del
gruppo delle creatine. E’ importante perché, come già detto, popolazioni cellulari diverse hanno filamenti
intermedi di natura diversa Quando ad esempio le cellule impazziscono, ad esempio nelle neoplasie, perdono la
capacità di riconoscere le simili, perdono la capacità di riconoscere i contatti, continuano a proliferare dando
origine ad ammassi, poi nei casi più gravi perdono il controllo di dove stanno e se ne vanno a giro a popolare altri
organi, le così dette metastasi. Pur nella loro completa “pazzia” mantengono i filamenti intermedi dalle cellule di
origine, quindi attraverso delle banali tecniche citologiche si possono sempre identificare il tipo di filamenti
intermedi, questo ci dà un vantaggio nel riconoscimento nelle cellule alterate, neoplastiche, e nella attuazione di
meccanismi terapeutici più mirati.
Incluso
Vi dò il concetto di incluso e quindi il citoplasma l’abbiamo finito. Gli inclusi sono strutture del citoplasma
espressione di una particolare funzione. Alcune cellule ne presentano un tipo altre ne presentano un altro, certe
cellule hanno inclusi altre non ne hanno, per questo non si ritrovano in maniera obiquitaria, sono cioè cose diverse
dagli organuli, sono NON gli strumenti per la funzione cellulare ma l’espressione di una particolare funzione
cellulare. Ce ne sono di diverso tipo. Si diceva prima dell’aggregazione tra miosina e actina , le miofibrille si
ritrovano come tali solo nelle fibre muscolari, tutte le cellule hanno filamenti di actina e molecole di miosina che
interagiscono, ma a formare strutture che si chiamano miofibrille ce l’hanno solo gli elementi muscolari. Così il
glicogeno si ritrova in certe parti, in fibre muscolari come sostanza di accumulo, di natura trofica. Oppure i lipidi,
le cellule adipose ce l’hanno altre cellule no. I pigmenti, anch’essi sono inclusi. Il pigmento melanico, i granuli di
melanina le possiedono le cellule dell’epidermide, altre cellule non le posseggono. I granuli secretori come tali,
fatti per esempio in un certo modo, come secrezioni di zimogeno (?) ce l’ha solo le ghiandole a secrezione
proteica altre no. Se poi arriviamo alla gonade maschile, e guardiamo un ghiandola chiamata l’interstiziale del
testicolo, queste cellule che secernono testosterone, posseggono nel loro citoplasma dei cristalli che sono
espressione di quel particolare tipo cellulare, di quell’organo solamente, per quel tipo di persona, cioè del sesso
maschile.
TERZA LEZIONE DI ISTOLOGIA.
Nucleo: è quella porzione che contiene il DNA, costituita dall’involucro nucleare che delimita il nucleo e divide il
materiale dal citoplasma.
All’interno troviamo un nucleo scheletro rete di strutture proteiche che si ancorano a varie zone della membrana
citoplasmatica.
All’interno del nucleo troviamo il nucleolo un corpiciattolo molto colorato elettronopaco al M.E di forma
rotondeggiante. La restante parte del nucleo è occupata dalla cromatina costituente specifico nucleare.
Un nucleo una cellula, esistono cellule prive di nucleo es. i globuli rossi. Cellule con 2 nuclei, con 4 nuclei,
cellule plurinucleate cioè distretti citoplasmatici in cui si vengono a ritrovare più nuclei. Ciascun nucleo terrà
sotto il proprio controllo metabolico una quota di citoplasma che gli starà attorno. Il rapporto di nucleo e quantità
di citoplasma tende ad essere costante.
Come si possono formare questi elementi plurinucleati?
Questi nascono per unione di singoli elementi che durante lo sviluppo mettono in comunione il citoplasma
segregando i vari nuclei all’interno.
Questa metodica viene chiamata Sincizio = elemento plurinucleato formatosi per unione di elementi singoli.
La struttura plurinucleata si può formare anche per plasmodio, cioè per divisioni successive di una cellula che
divide il proprio nucleo ma non divide il citoplasma. Il nucleo dividendosi più volte da origine a più nuclei tutti
contenuti in quel citoplasma che lentamente si ingrandisce.
Plasmodio = elemento plurinucleato che origina da una cellula primitivamente singola, mononucleata che divide
il proprio nucleo ma non il citoplasma. Avremmo all’inizio una cellula binucleata, poi i nuclei si dividono e danno
origine a 4, il citoplasma si ingrandisce ma non si divide, da 4 ne otterremo 8…..
Il nucleo si occupa:
della trasmissione dei caratteri da una cellula ad un’altra.
tiene sotto il proprio controllo la riproduzione cellulare.
controlla il metabolismo cellulare attraverso la sintesi proteica.
Membrane nucleari
L’involucro nucleare è costituito da due membrane unitarie di cui:
quella più esterna ( che guarda verso il citoplasma ) si presenta ruvida perché porta adesi i ribosomi ed è in intimo
contatto con le cisterne del R.R.
La membrana interna invece è liscia priva di ribosomi e guarda verso l’ambiente intracellulare.
Le membrane si continuano l’una nell’altra a livello di aperture chiamati pori nucleari i quali vanno a costituire
delle strutture rotondeggianti che sono i punti di passaggio facilitato tra nucleo e citoplasma.
Alla membrana interna prende adesione la componente citoscheletrica del nucleo.
Le membrane mediano il passaggio di sostanze secondo le regole della permeabilità, ma si creano delle strutture
che aumentano ed alzano la possibilità di passaggio delle sostanze, anche se questi pori nucleari che si trovano
lungo tutto il percorso dell’involucro nucleare non sono delle porte spalancate, ma bensì dei passaggi altamente
regolati dalla presenza di proteine, proteine del poro. Queste ultime funzionano come un diaframma aprendo e
chiudendo a seconda del controllo questa possibilità di passaggio.
Interno del nucleo
Entriamo dentro il nucleo.
Il nucleo al M.O. appare come un corpiciattolo molto colorato di forma rotondeggiante.
Di solito c’è un nucleolo per ogni nucleo, ma esistono esempi di nuclei con più nucleoli e anche di nuclei in cui
non si vede il nucleolo. Al M.E. appare come una zona che alterna porzioni più elettronopache e meno.
Di fatto il nucleolo altro non è che il sito di formazione dei ribosomi o unità ribosomiali. Infatti queste unità
nascono nel nucleo vanno nel citoplasma e lì si complessano a formare quella sferula che è il ribosoma.
Nucleolo = è una struttura nucleare sito di formazione delle sub-unità ribosomiali.
Nel nucleo vi è il DNA ( controlla la sintesi proteica e la formazione dell’RNA ) un cui tratto controlla la
formazione dell’RNA ribosomiale. A questo RNA si uniscono le proteine provenienti dal citoplasma e si
complessano a formare delle strutture rotondeggianti che altro non sono che le subunità dei ribosomi. Queste
subunità piano piano che si formano prendono la via dell’attraversamento dell’involucro nucleare, vanno nel
citoplasma e formano i ribosomi.
Cromatina = insieme di strutture filamentose o granuli costituite da DNA e PROTEINE. Quest’ultime si
dividono in:
Proteine istoniche, di natura basica.
Proteine non istoniche, di natura acida.
La cromatina è il materiale di cui è costituito maggiormente il nucleo, ed essendo costituita da DNA ne forma la
porzione specifica.
Osservando al M.O. cellule diverse il nucleo a volte appare poco o per niente colorato, per cui la cromatina venne
chiamata la cromatina vera che non si colora. Es. neurone = la cromatina appare come una sferula bianca in cui
spicca il nucleolo. Mentre in altri tipi di cellule ( es. il linfocita ) il nucleo viceversa è talmente colorato in rosso
vivo che sembra essere riempito da una sostanza tale che non fa vedere nemmeno il nucleolo. Questo tipo di
cromatina la chiamiamo l’altra cromatina cioè eterocromatina.
Eucromatina ed eterocromatina sono un linguaggio per descrivere una morfologia al M.O. di un’unica realtà, la
cromatina che è DNA più proteine.
Perché in alcune cellule non si colora la cromatina ed in altere si?
Il neurone e il linfocita sugli estremi di una variabilità di disegno cromatico, alterano zone molto colorate a zone
meno. Per cui il disegno cromatico è stato oggetto di denominazioni diverse, paragonando la cromatina che si
colora a “zolle”.
Eucromatina ed eterocromatina non sono che immagini diverse al M.O. di una stessa realtà, la cromatina DNA più
proteine.
La molecola di DNA è un doppio filo ad elica che si gira attorno ad un rocchetto dato da un’insieme di molecole,
gli ISTONI. Un ottametro, otto molecole di istoni che si uniscono formando una specie di cilindretto. Il DNA si
gira attorno agli istoni quasi due volte per poi percorrere un tratto isolato proseguendo dinuovo successivamente
con altri due avvolgimenti e così via, creando quella che ci ricorda una collana di perle. La lunghezza del DNA si
accorcia in seguito agli arrotolamenti che compie attorno a questi isomeri di istoni. Pensiamo di arrotolare ed
impacchettare questa collana facendola divenire sempre più corta e visibile, ecco che si passa dalla eucromatina
alla eterocromatina quella che si comincia a colorare.
Il massimo di questi impacchettamenti si avrà nel cromosoma che è l’immagine della cromatina quando la cellula
si divide. La cromatina è un insieme di filamenti nucleosomici (dna e proteine).
Quanti filamenti nucleosomici avremo? 1 per ogni cromosoma, la specie umana ha 46 cromosomi.
44autosomi uguali a due a due, quindi 22 coppie di cui una l’abbiamo ereditata dalla madre e una dal padre che
tengono a controllo le caratteristiche generali del nostro organismo e
2cromosomi sessuali, 2X se è un individuo di sesso femminile e 1X e 1Y se è un individuo di sesso maschile.
Cromatina = DNA più proteine, costituite da un insieme morfologico di fibrille nucleosomiche.
La fibrilla nucleosomica si può impacchettare in maniera diversa e dare immagine al M.O. più o meno colorate.
Questa è l’immagine del nucleo quando la cellula non si divide, cioè quando porta avanti tutte le funzioni
connesse con la vita.
Esistono poi dei momenti in cui la cellula si divide, da una cellula se ne originano due.
Nel momento in cui la cellula si divide, questa suddivide il proprio patrimonio in cellule figlie, lo ridistribuisce
avendo duplicato il proprio DNA.
Quando dalla cromatina si vedono i cromosomi significa che il DNA ha duplicato la propria struttura da una
fibrilla nuleosomica ne sono venute due in tutto e per tutto uguali.
Cromosoma = immagine della cromatina quando la cellula si divide. I cromosomi sono strutture con 2 filamenti
ravvicinati nello spazio e tenuti insieme in un punto specifico detto centromero.
Questa morfologia è la spia del fatto che quando la cellula si divide ha duplicato le proprie fibrille cromosomiche
e da 1ne ha fatte 2, ciascuna delle quali prende il nome di cromatidio.
Durante l’interfase quando la cellula porta avanti le proprie funzioni e non si divide, nel nucleo vediamo la
cromatina. Successivamente la cellula in un certo momento sente degli stimoli, il citoplasma si ingrandisce si
altera il rapporto nucleo plasma e la cellula si divide.
Ma per fare ciò la cellula ha bisogno di duplicare il proprio DNA per poi risuddividerlo in cellule figlie.
Il cromosoma è l’espressione della duplicazione avvenuta del DNA.
Quando la cellula entra in divisione viene chiamato processo di mitosi-caricinesi = riproduzione delle cellule
somatiche, tutte le cellule dell’organismo ad eccezione delle cellule sessuali che danno origine ai gameti.
Queste cellule sessuali si dividono per meiosi, processo riduzionale in quanto crea cellule figlie con contenuto
dimezzato di fibrille nucleosomiche, i gameti.
Mitosi = divisione equazionale delle cellule somatiche. Equazionale cioè significa che la cellula madre prima di
dare origine a 2 cellule figlie duplica il proprio patrimonio genetico per poi poterlo suddividere.
La mitosi è una serie di eventi che si suddivide in fasi.
Cariotipo: corredo cromosomico di un individuo.
Sono nuove tecniche che sfruttano la divisione cellulare e riuscendo a bloccare, attraverso particolari sostanze, le
divisioni cellulari in una specifica fase detta metafase (in cui i cromosomi hanno la loro massima condensazione e
quindi appaiono ben visibili come strutture) riuscendo così a fotografare il cariotipo.
Nell’individuo adulto si preleva il sangue, si mette in cultura i linfociti, induciamo loro a dividersi, li blocchiamo
in metafase, li coloriamo e infine li fotografiamo.
Tagliamo e uniamo i cromosomi a seconda della lunghezza, della posizione del centromero etc. ed otteniamo il
cariogramma.
Cariogramma = una rappresentazione del corredo cromosomico.
È possibile così vedere cromosomi sovrannumerali che possono dare origine a trisonomie come la trisonomia 21,
sindrome di Down, oppure possiamo vedere monosemie etc. Anche durante la gestazione è possibile avere un
cariotipo dell’embrione attraverso il liquido amniotico prelevando i villi della placenta.
Ciclo cellulare.
Quando la cellula nella sua vita alterna in maniera continua le sue funzioni e poi si divide, stabilisce quello che si
chiama un ciclo cellulare.
Porta avanti le sue funzioni e ad un certo momento duplica il proprio DNA, doppio DNA significa doversi
dividere.
Il ciclo cellulare è un giro tondo che si può schematizzare con una circonferenza:
un periodo intervallo1(GAP in inglese ) G1 che segue la divisione in cui la cellula porta avanti le sue funzioni,
successivamente in seguito a segnali duplica il proprio DNA, questo viene nominate periodo S di sintesi, in cui la
cellula duplica il proprio patrimoni genetico.
Poiché quest’ultimo è stato duplicato è inevitabile passare all’intervallo2 o G2, in cui la cellula si prepara alla
divisione, alla mitosi.
Questo ciclo cellulare può anche essere descritto come l’interfase, periodo S, periodo G2, mitosi.
Mentre il periodo G1 può essere molto variabile da cellula a cellula, per alcune può durare tutta la vita, il periodo
S di sintesi G2 e mitosi è pressocchè uguale per tutte.
Il ciclo cellulare può essere descritto in base a quanto dura il periodi G1, ci sono cellule che continuano a ciclare
per tutta la vita dando origine a più generazioni cellulari.
Altre escono dal giro tondo dando origine ad un periodo G0 che può durare tutta la vita in maniera irreversibile,
cioè quelle cellule non possono più dividersi.
Oppure periodo di reversibilità, perché se tutto va bene quelle cellule non tornano a ciclare altrimenti posseggono
ancora la proprietà intrinseca di fare la divisione.
“Bizzozero” un ricercatore dette questa classificazione in base al tipo di ciclo cellulare che esse presentavano,
dividendole in :
LABILI, quelle che ciclano continuamente.
STABILI, quelle che escono da questo ciclo in maniera reversibile.
PERENNI, quelle che ormai non ciclano più.
Sviluppo embrionale.
Noi siamo il risultato dello sviluppo di una cellula uovo fecondata.
La cellula uovo che ci proviene da nostra madre viene fecondata dagli spermatozoi provenienti dal padre, uno di
questi spermatozoi attraverso gli enzimi dell’acrosoma scinde la membrana della cellula uovo e vi entra dentro.
Una volta che lo spermatozoo è entrato dentro la cellula, questa da origine ad una reazione corticale che
impedisce ad altri spermatozoi di penetrare, è avvenuta la fecondazione.
Lo spermatozoo perde tutte le porzioni che non gli servono più ed unisce il proprio nucleo a quello della cellula
uovo la quale grazie a RNA e mitocondri porta avanti il proprio metabolismo.
Questa unione fra i due nuclei da origine alla ZIGOTE la prima cellula dell’organismo.
Noi deriviamo in tutta la nostra variabilità cellulare e tissutale dallo zigote.
Poi segue lo sviluppo, la fecondazione avviene nelle tube, lo zigote scende nella cavità uterina dove trova
l’endometrio, sotto stimolo ormonale ben sviluppato vi si annidare portando avanti la gestazione.
Dentro questo annidamento lo zigote porta avanti il suo sviluppo, non si chiamera più zigote ma blastocisti
perché nel frattempo si è diviso e da 1 cellula se ne sono originate 2, questi blastomeri daranno origine a 4 poi a 8
cellule etc. fino al momento in cui questa cellula costituita da tanti blastomeri non avrà raggiunto la morfologia di
una mora dalla quale prenderà il nome di morula.
Dopo aver proliferato e continuato a proliferare queste cellule come seguendo un destino scritto dentro il proprio
DNA, che esprime attraverso situazioni di localizzazione, si vengono a trovare spinte da migrazioni, dovuta alla
pressione cellulare esercitata da cellule accanto,in posizioni diverse. Alcune daranno origine al rivestimento
esterno e daranno origine agli annessi embrionali es.placenta. tutte quelle strutture embrionali e poi fetali che
assicurano lo sviluppo e quello che poi sarà del nuovo individuo. Altre invece si rannicchiano e danno origine al
vero e proprio corpo del futuro individuo.
Queste cellule dell’individuo futuro ad un certo momento daranno origine ai così detti 3 foglietti embrionali:
ectoderma – mesoderma – endoderma.
Ad opera di questi 3 foglietti embrionali poi se ne formerà un altro che non è un foglietto ma un tessuto di cellule
e liquido che si chiama MESENCHIMA.
Da questi 3 primitivi agglomerati di cellule si formano tutte le strutture del nuovo organismo. Siamo prodotti per
il differenziamento dei foglietti embrionali che si aggregano.
Il fenomeno del differenziamento, che è attivo durante lo sviluppo embrionale e fetale, è un processo attivo
prenatale e postnatale che porta una cellula, a partire da quella primitiva cioè lo zigote, alla molteplicità delle
cellule del nostro organismo diverse per morfologia, costruite però tutte su uno stesso schema esprimono
morfologie diverse per funzioni diverse.
Le cellule possono morire attraverso due meccanismi diversi.
Necrosi = detta anche morte accidentale, è un evento non previsto nella vita cellulare. Nel morire da origine ad
una lisi della propria membrana e questo risveglia una reazione da parte dell’organismo che riconosce questa
situazione come non propria, da origine ad una infiammazione localizzata che risveglia le cellule dell’immunità.
La apoptosi è una morte programmata nella vita della cellula che sa quando è venuta l’ora. È una morte
silenziosa la cellula comincia a regredire perché non trascrive e quindi non sintetizza proteine, comincia a perdere
gli organuli perché il suo nucleo diventa sempre più piccolo impallidisce e poi scompare. La cellula senza dare
origine a nessuna reazione si toglie di mezzo per morte programmata.
Popolazione cellulare = un insieme di cellule contraddistinte dalla composizione chimica , dall’aspetto
macroscopico e dalla funzione specifica.
Ecco quindi che si possono raggruppare le cellule in popolazioni in base alla loro derivazione embriologica, alla
loro composizione chimica, all’aspetto macroscopico, alle differenze diverse e quindi alla diversa funzione.
Sostituiamo così popolazione cellulare con il termine cellula che della popolazione ne è componente.
Le cellule ora le vediamo come un insieme integrato di strutture che tutte collaborano alla funzione di quella
popolazione cellulare dove ognuno ha il proprio compito.
Le popolazioni cellulari possono essere divise a seconda della dinamica in:
Popolazioni soggette a rinnovo = fisiologicamente riescono rimpiazzare le perdite
Popolazioni non soggette a rinnovo = quando la cellula è perduta (neuroni) non vi è nulla da fare.
A livello delle popolazioni cellulari soggette a rinnovo potremmo distinguere anche dal punto di vista
morfologico oltre che funzionale una parte di cellule che si mantengono tali da poter ciclare e sostituire le perdite.
Compartimento germinativo, sono le cellule che ciclando e danno origine a cellule che ripristinano le perdite.
Il compartimento maturativo è invece l’insieme delle cellule che porta avanti la funzione.
Le cellule staminali, una componente del tessuto germinativo cioè la porzione più resistente a qualsiasi tipo di
aggressione esterna e agli agenti ionizzanti, di regola si dividono poco ma intervengono nei momenti critici delle
perdite.
Le cellule staminali a cui si affida il nostro organismo per le riprese massive, sono capaci di dividersi per tutta la
loro vita. Esse possono generare sotto stimolo appropriato tutti i tipi di popolazioni cellulari ( PLURIPOTENTI),
oppure mantengono la possibilità di dare una gamma di tessuti (es. le cell.del midollo emopoietico sono capaci di
dare origine a tante linee cellulari , le cellule del cordone ombelicale sono quasi totipotenti cioè possono dare
origine sotto stimoli particolari a cellule di vario tipo).
Cellule staminali = altamente resistenti poco differenziate che possono dare origine sotto stimoli appropriati a
cellule diverse.
I tessuti dell’organismo sono 4:
Tessuto epiteliale
Tessuto connettivo → sangue
Tessuto muscolare
Tessuto nervoso
Le cellule che si aggregano a formare i tessuti si riconoscono tra di loro e aderiscono tramite il glicocalice, questa
adesione può essere rafforzata da giunzioni, strutture che legano tra loro il plasmalemma delle cellule ravvicinate
dando origine a strutture che prendono nome di differenziazioni laterali delle cellule (?) perché mettono in stretta
comunicazione le porzioni laterali. 3 tipi di giunzioni:
Occludente
Aderente
Comunicante
-La giunzione occludente è una giunzione che si sviluppa su tutto quanto il perimetro della cellula e chiude gli
spazi fra le cellule. Una serie di proteine si agganciano tra loro, queste proteine di membrana si incastrano con le
proteine intrinseche delle membrane vicine chiudendo gli spazi aperti fra le cellule. Hanno quindi il significato di
limitare la permeabilità di lamine cellulari perché tutto quello che passa da un versante viene tenuto a controllo
dalle cellule stesse.
-Le giunzioni aderenti sotto forma di zonula o macula ancorano le cellule fra di loro per mezzo di sistemi proteici
trasmembrana che poi sul versante citoplasmatico legano i componenti del citoscheletro, rendono così le cellule
saldamente legate le une alle altre anche in sinercismo con gli elementi del citoscheletro. La macula o desmosoma
lega i componenti intermedi del citoscheletro, mentre la zonula lega i filamenti sottili del citoscheletro.
-Giunzioni comunicanti, punti in cui le cellule vicine possono comunicare tra di loro grazie a dei canali che
passano da una cellula all’altra.
Cosa passa da una cellula all’altra?
Possono passare piccole molecole ( funzione regolativa ), RNAm, piccole molecole a carattere trofico per il
metabolismo cellulare e ioni.
Abbiamo la loro massima espressione nel tessuto epiteliale che è per antonomasia è il tessuto con cellule a mutuo
contatto.
Tessuto epiteliale.
Insieme di tessuti accumunati dal fatto di essere costituiti da cellule a mutuo contatto. Tra le cellule c’è solo lo
spazio del glicocalice. Aggregato di piastrelle, il contorno sferico e sinuoso.
Cellule poliedriche:
Pavimentose → schiacciate
Cubiche
Cilindriche → alte e strette
Le cellule sviluppano tutti i tipi di giunzioni in sequenza, complesso di giunzioni.
Studiare da soli ciglia e microvilli.
Istologia quarta
TESSUTI EPITELIALI:
Sono tessuti costituiti da cellule a stretto contatto, di forma poliedrica, che presentano specializzazioni della parete
cellulare sotto forma di giunzioni e differenziazioni apicali che sono per esempio i microvilli, espansioni
digitiformi della superficie cellulare in relazione al fenomeno dell’assorbimento (vedi libro per struttura e
funzioni) hanno una morfologia a “dito di guanto” con un asse di microfilamenti di actina (fateveli da soli) e sono
espressione di un aumento della superficie della cellula. Le ciglia, altre estroflessioni della superficie cellulare
apicale, hanno una struttura interna data da doppiette di microtubuli (9 periferiche e 2 centrali) sono strutture
ancorate all’apice cellulare su una struttura centriolare e sono in grado, grazie a lamine interne di microtubuli, di
portare avanti il movimento, che non è un semplice battere nel vuoto, ma che è un fenomeno che permette alla
cellula di far avanzare le secrezioni che vengono prodotte dalla cellula stessa per vari scopi e che si depositano
sull’apice di queste strutture (vedi disegnino) Il movimento del ciglio è simile a quello delle spighe di grano in un
campo, sospinte dal vento “ad onda”: parte dalla posizione dinamicamente più vantaggiosa (leggermente arretrata
rispetto all’asse perpendicolare della cellula), batte cadendo e graffiando lo strato secretivo che giace al proprio
apice e lo sposta. Ovviamente il movimento di ritorno non può essere l’inverso di questo, perché altrimenti
annullerebbe l’effetto del battere, così le lamine del ciglio si srotolano al di sotto dello strato mucoso spostato e
ritornano nella posizione di partenza. Il movimento del ciglio consiste quindi in due momenti specifici: un “colpo
efficace” e uno “di ritorno”. Le ciglia sono disposte in file e si muovono in sincrono (ciascuna si abbatte
immediatamente prima di quella che la segue e immediatamente dopo di quella che la precede) proprio come le
spighe in un campo di grano battuto da vento, questo movimento è detto metaclonato(?). Se io guardo le ciglia in
direzione ortogonale all’osservatore una fila è tutta nella stessa posizione di battito cigliare, spostando ad ogni
battito un fluido che si deposita all’apice nella medesima direzione.
Fluido depositato all’apice
Ciglia in movimento
Cellula epiteliale
Strato basale
emidesmosomi
Si è detto anche che la cellula epiteliale possiede nella sua faccia inferiore delle strutture di ancoraggio alla
membrana basale gli emidesmosomi, perciò possiamo dire che la cellula epiteliale, che è sfaccettata, mostra delle
caratteristiche diverse a seconda che ne guardi le varie facce. Questa diversità è anche sottolineata da dislocazioni
degli organuli all’interno della cellula che segue il percorso funzionale della cellula stessa.
Un esempio per tutti: una cellula ghiandolare (vedi immagine sul libro) è un trapezio che poggia su un connettivo
sottostante che porta i vasi da cui dipende il trofismo di questa cellula, all’interno della cellula vediamo il reticolo
endoplasmatico, quindi il nucleo e sopra l’apparato di Golgi,. Sappiamo che le proteine secrete vengono
sintetizzate prima nel reticolo poi passano al Golgi dal quale emergeranno come granuli secretori, già vicini
all’apice cellulare, da dove con processo di esocitosi fuoriusciranno.
E’ evidente una polarità di questa cellula rispetto alla sua funzione che si rispecchia nella sua morfologia:
parliamo quindi per le cellule epiteliali di POLARITA’ MORFOFUNZIONALE (ineguale distribuzione degli
organuli all’interno della cellula - porzione osservata - in relazione alla sua funzione) I filamenti intermedi delle
cell epiteliali si chiamano POLIFILAMENTI (?), perché formati da proteine, le cheratine – chiedere a lei
conferma se poli = polimero o altro sigificato–
Gli epiteli sono privi di vasi sanguigni e sono un esempio di popolazione cellulare soggetta a rinnovo, sono
innervati, poggiano su connettivo, tessuto che per antonomasia porta i vasi sanguigni, dal quale dipende il loro
trofismo, che avviene per fenomeni di capillarità-diffusione attraverso i capillari dal sangue dell’ossigeno e dei
nutrienti provenienti dai processi digestivi e di assorbimento dell’intestino, questi attraverso la sostanza
intercellulere del connettivo giungeranno al tessuto epiteliale, dal quale poi attraverso un percorso inverso,
porteranno via i prodotti del catabolismo cellulare e l’anidride carbonica. Gli scambi che avvengono tra tessuto e
capillari vengono detti EMATOTISSUTALI.
Tra TESSUTO EPITELIALE e CONNETTIVO esiste una struttura ubiquitaria che fa da interfaccia tra connettivo
e ciò che non lo è (in questo caso il tessuto epiteliale), conferendo la massima aderenza e possibilità di scambi
ematotissutali trai due tipi di tessuto. Questa struttura è detta MEMBRANA BASALE, e verrà trattata con il
tessuto connettivo pur se in parte è di pertinenza anche degli altri tipi di tessuto, per ora è sufficiente sapere che è
costituita di due porzioni: una detta LAMINA BASALE, che è prodotta dal tessuto sovrastante (in questo caso
quello epiteliale) e una porzione prodotta dal tessuto connettivo stesso detta LAMINA RETICOLARE. La sua
funzione è assicurare l’ancoraggio delle cell al sottostante connettivo, permettere gli scambi ematotissutali e
permettere gli spostamenti delle cellule lungo il margine della membrana basale stessa.
Le mnb basali sono strutture che mutano, cambiando di conformazione e spessore in base alla loro dislocazione e
anche in conseguenza di numerose malattie metaboliche.
I tess. epiteliali sono INNERVATI, ossia le terminazioni nervose dal tess. connettivo valicano la memb basale e
vanno a raggiungere, insinuandosi tra le cell epiteliali a mutuo contatto, cellule o aggregati cellulari addetti alla
secrezione (ghiandole) regolandone l’ECCITOSECREZIONE o aree sensibili, mediandone la sensibilità
I tess. epiteliali sono esempi di popolazioni cellulari soggette a rinnovo, che è un processo fisiologico con ritmi
diversi in funzione del tipo di epitelio che consideriamo, legato alla presenza di un compartimento germinativo
Il turn over cellulare è più breve per gli ep. di rivestimento, a sua volta differenziato come rapidità a seconda della
funzione specifica dell’ep. di rivestimento – epitelio intestinale: 2-3 gg, epidermide: 20 gg - e più lento per gli
epiteli ghiandolari, che a volte non si rinnovano per tutta la vita, ma avendo in sé questa capacità, possono
all’occorrenza ripristinare la loro funzionalità perduta in conseguenza di danno meccanico o dovuto a patologia.
I tessuti sono classificati in base a specifici criteri:
Definizione: enunciando le caratteristiche generali, poi si introducono i criteri classificativi e si dividono in
gruppi.
Su base funzionale i tessuti epiteliali si dividono in:
DI RIVESTIMENTO
GHIANDOLARI O SECERNENTI
SENSORIALI (li definiremo soltanto)
PARTICOLARMENTE DIFFERENZIATI (non li tratteremo)
I tessuti epiteliali derivano da tutti e tre i foglietti embrionali (ectoderma, mesoderma e endoderma)
Tratteremo anche un tessuto di derivazione mesenchimata (tessuto padre del connettivo), poiché per le sue
caratteristiche richiama quello dei tessuti epiteliali.
EPITELI DI RIVESTIMENTO
RIVESTONO CON UNA LAMINA, INTERFACCIATA DA UNA MEMBRANA BASALE, UN TESSUTO
CONNETTIVO SOTTOSTANTE, come localizzazione possono RIVESTIRE LA SUPERFICE DEL CORPO
(epidermide) o CAVITA’ IN ESSO CONTENUTE che possono essere comunicanti con l’esterno(mucose), o no
(endotelio, è questo il tessuto con caratteristiche epiteliali, ma di origine mesenchimale che va a rivestire le
camere cardiache e tutte le pareti del sistema circolatori). Vi sono inoltre altri spazi che sono rivestiti da tessuto
epiteliale e sono gli spazi perilinfatici dell’orecchio e la camera anteriore dell’occhio, che non tratteremo e le
cavità correlate col sistema nervoso centrale che verranno riprese al momento della trattazione di quel sistema.
Anche nel tes. Epiteliale di rivestimento troviamo tutte le caratteristiche peculiari degli epiteli, la poliedricità delle
cell, la loro polarità morfofunzionale, la stretta connessione tra loro e l’ancoraggio al tessuto connettivo trofico
tramite la memb basale. Questa caratteristica rende tutti gli epiteli uguali tra loro se osservati in sezione
trasversale, ma se io farà una sezione sagittale allora potrò osservare le diversità citoplasmatiche presenti, un po’
come se osservassi una casa dal tetto alla cantina anziché stando sul pavimento
Per classificare gli epiteli di rivestimento si invocano due criteri classificativi:
1)
stratificazione lamina epiteliale SEMPLICI o MONOSTRATIFICATI -1 solo strato di cell,tutte
poggianti sulla memb basale
PSEDUDOSTRATIFICATO a più file di nuclei , con cell tutte poggianti sulla memb basale ma aventi altezze
diverse e nuclei posti ad altezze diverse
COMPOSTI o PLURISTRATIFICATI più strati di cell di cui solo il primo strato poggia sulla membrana
basale
2)\forma delle cellule EPITELI PAVIMENTOSI POLIEDRICI appiattita (simile ad una mattonella)
CUBICI con lati di uguali dimensioni
CILINDRICI, in realtà sono parallelepipedi la cui dimensione maggiore è perpendicolare alla membrana
basale.
Dalla combinazione dei due criteri di classificazione avrò la possibilità di definire tutti gli epiteli di rivestimento
presenti nel corpo umano.
Ep. semplici pavimentasi, cubici, cilindrici e cilindrici pseudostratificatI
Ep. composti (il nome è dato dalla forma delle cellule dello strato che aggetta slla sup libera, in quanto le cellule
dello strato basale sono sempre cubiche , quelle degli strati intermedi possono assumere varia forma) pavimentosi
e cilindrici, manca il cubico e poi abbiamo un epitelio di transizione bistratificato, con caratteristiche peculiari
LE FUNZIONI DELL’EPITELIO DI RIVESTIMENTO.
La funzione principale è quella di protezione, massimamente espressa dagli epiteli composti, mentre negli epiteli
semplici possono prevalere altre funzioni, come quella di assorbimento (ad es.: a livello di mucosa intestinale) di
trasporto di fluidi frutto di secrezione cellulare attraverso il movimento ciliare (mucosa delle prime vie aeree), la
secrezione (epitelio della mucosa gastrica) o gli scambi gassosi
IL COMPARTIMENTO GERMINATIVO
Le cellule del compartimento germinativo sono dislocate tra la lamina semplice, via via che questa perde elementi
lo strato germinativo li ricostituisce rinnovando l’epitelio. Negli epiteli composti lo strato germinativo è quello
cubico che poggia sulla memb.basale e…..???
EPITELIO PAVIMENTOSO SEMPLICE
(prototipo di risposta d’esame)
E’un tessuto epiteliale, ossia di rivestimento di una cavità interna, poiché l’epidermide che riveste esternamente il
corpo è costituita da un epitelio pluristratificato, come tutti gli epiteli è costituito da cell a mtuo contatto, di forma
poliedrica e caratterizzate da polarità morfofunzionale, ancorate ad un connettivo tramite una memb basale. Tre
esempi:
APP.RESPIRATORIO.: epitelio di rivestimento dell’interno dell’alveolo polmonare, che è l’unità funzionale
del polmone, dove avvengono gli scambi gassosi tra anidride carbonica e ossigeno nel sangue. L’alveolo ha la
forma di un acino d’uva, una sfera cava il cui lume interno, pieno di aria inspirata, è appunto rivestito di un
epitelio pavimentoso semplice, che poggia su una memb.basale a sua volta ancorata ad un connettivo densamente
vascolarizzato che trasporta il sangue refluo dall’aver circolato nell’organismo, carico di anidride carbonica, e qui
per differenza di pressione, libera la libera arricchendosi di ossigeno presente nell’aria inspirata.
La membrana basale dell’alveolo polmonare ha la caratteristica di non presentare il reticolo basale, proprio perché
trai capillari presenti nel connettivo e l’aria presente nell’alveolo ci deve essere meno barriera possibile.
Le cellule che costituiscono l’alveolo polmonare vengono chiamate pneumociti di primo tipo.
Dal vaso sanguifero l’anidride carbonica attraversa
Il percorso che fa l’anidride carbonica dai capillari all’alveolo è il seguente:
la parete del vaso stesso, penetra la lamina basale
alveoli
dell’alveolo polmonare e attraverso il citoplasma del
Pneumocita di secondo
tipo,
cell cubica,
costituente
il
compartimento
germinativo
e
secernente il fattore
sulfattante
Vasi sanguigni
Membrana basale,
costituita
dalla
sola lamina, di
pertinenza
dell’epitelio
Epitelio pav sempl,
cell con porzione
slargata dove c’è il
nucleo costituito da
PNEUMOCITI
DI
PRIMO TIPO dal
citoplasma sottile, che
favorisce gli scambi
Interstizio connettivale
Riccamente vascolarizzato
pneumpocita dell’epitelio pavimentoso semplice di
rivestimento interno dell’alveolo si libera in cavità.
Percorso inverso farà l’ossigeno per raggiungere i
globuli rossi nel vaso sanguifero e legarsi
all’emoglobina, con rinnovamento del pool di
ossigeno Una lamina di rivestimento sottile, ma
estremamente estesa garantisce
tappezzare queste cavità con maggior efficienza
degli scambi respiratori, ma nell’epitelio pav sempl
di rivestimento dell’alveolo sono presenti anche cell
cubiche (PNEUMOCITI DI SECONDO TIPO),
poco adatte agli scambi, ma che hanno il compito
ben preciso di secernere una sostanza chiamata
SULFATTANTE, di natura lipoproteica, che viene
riversata nel lume dell’alveolo, stratificandosi sulla
sua superficie impedendo all’alveolo stesso di
collassare durante l’atto espiratorio,
infatti l’ambiente alveolare è umido e una volta svuotato dall’aria, per fenomeni di tensione superficiale, le cariche presenti sulle pareti
potrebbero farle collabire, annullando la pervietà delle camere alveolari.
E’ importante conoscere questo fenomeno, perché gran parte dei problemi respiratori dei bambini nati immaturi, deriva dalla mancata
efficienza degli pneumociti di secondo tipo nella produzione di fattore sulfattante.
Gli pneumoniti di tipo due sono importanti anche perché costituiscono lo strato germinativo delle cell del tessuto epiteliale dell’alveolo
polmonare. Dalla loro divisione si hanno due linee cellulari distinte, una che andrà a ricostituire gli pneumoniti di primo tipo e l’altra
quelli di secondo tipo
APP. CARDIOCIRCOLATORIO:L’epitelio pavimentoso semplice che va a rivestire le cavità interne di tutto
l’apparato cardiocircolatorio, comprese le camere cardiache, viene chiamato ENDOTELIO. E’ molto importante
perché è a livello dell’endotelio dei capillari che avvengono tutti gli scambi di gas e nutrienti trasportati dal
sangue con i tessuti (scambi EMATOTISSUTALI)
Nei confronti dell’endotelio esistono due scuole di pensiero, quella che lo classifica tra i connettivi, per la sua
origine embrionale dal tessuto mesenchimale e quella, a cui ci rifaremo noi, che lo classifica tra gli epiteli per la
sua struttura (pavimentosa semplice) e funzione (di rivestimento, protezione e scambio).
L’endotelio è dunque un tessuto di derivazione mesenchimale, con aggregazione e funzioni di tipo epiteliale
E’ la parte più interna dei vasi e poggia su un connettivo ancorandovisi con la menb basale, tutto ciò costituisce il
vaso che se è di piccolo calibro, come i capillari è il vaso in toto, se invece si tratta di vasi di calibro maggiore la
parete si arricchisce di altre tonache di altra natura.
Sono cell sottili connesse tra loro per mezzo di interdigitazioni della parete cellulare stessa. Inoltre l’endotelio ha
anche la caratteristica di poter creare dei varchi sulla parete, varchi tali da far uscire dal circolo sanguigno i
globuli bianchi che con movimenti ameboidi strisciano sulle pareti dei vasi e così possono penetrare nel
connettivo e svolgere la loro azione di difesa. Questo fenomeno si chiama DIAEDESI.
Un altro tipo di giunzione che possono attuare le cell dell’endotelio è quello delle g. aderenti e infine possono
anche creare delle g. OCCLUDENTI. Queste limitano molto gli scambi e sono importanti per impedire l’accesso
di patogeni e permettere l’ingresso selettivo di nutrienti in taluni distretti del corpo. Stiamo parlando della
BARRIERA EMATOENCEFALICA, che protegge, giustappunto l’encefalo da aggressioni esterne e fornisce
un medium nutritizio adatto.
In altri casi, p.e. le ghiandole endocrine, dove l’ormone prende come via di escrezione il circolo sanguigno, le
membrane delle cellule collabiscono, annullando il citoplasma, fino a creare dei varchi (FENESTRATURE).
Pertanto posiamo dire che in base al distretto in cui ci troviamo e alla funzione che il vaso ha possiamo riscontrare
diverse tipologie di giunzioni delle cell del tessuto di rivestimento dei vasi stessi, ossia dell’endotelio.
G. occludenti = passaggio selettivo; g. aderenti = ancoraggio meccanico; fenestrature = passaggio materiali trans
cellula endoteliale, proveniente dal sangue ai tessuti connettivi avviene per MICROPINOCITOSI (vedi precedenti
lezioni).
Spesso le vescicole derivanti dalla micropinocitosi si creano sul versante luminale del capillare, poi attraversano il
citoplasma dell’endotelio del capillare e vengono scaricate dall’altra parte, quindi servono a mediare il trasporto
di materiali, in modo veloce, ma molto controllato.
Oltre al rivestimento, l’endotelio ha altre funzioni regola, ad esempio, la permeabilità vascolare diminuendo o
aumentando il grado di sostanze che possono passare, poi regola l’aggregazione piastrinica (le piastrine sono
elementi del sangue circolanti che presiedono alla funzione della coagulazione), permettono la diapedesi
leucocitaria, sintetizzano le macromolecole della matrice, si fabbricano la lamina basale, entrano nel rinnovo della
sostanza intercellulare del connettivo e sintetizzano CITOCHINE (sostanze che presiedono al processo di
riparazione di danni cellulari di varia origine, richiamando cell della difesa o attivando cell che nel tess connettivo
si incaricano dei processi di riparazione)
Altro tessuto di origine mesenchimale è il MESOTELIO, questo tessuto detto anche SIEROSA è quello che va a
costituire i foglietti che rivestono le cavità toracica (PLEURE), cardiaca (PERICARDIO) e addominale
(PERITONEO). Questo tessuto ha la caratteristica di presentare una lamina basale estremamente ridotta e
presenta un foglietto parietale (addossato alla parete della cavità) e uno viscerale addossato all’organo, tra i due
foglietti vi è un velo di liquido, sieroso, appunto, che serve ad ammortizzare gli attriti dell’organo durante i
movimenti. Questo liquido è prodotto e riassorbito in continuo grazie ad una serie di microvilli presenti sulla
parete interna dei foglietti, che ne mantengono la quantità costante. Causa di accumulo di liquidi sono i danni a
questo epitelio che non è più n grado di espletare le sue funzioni di regolatore.
Un altro esempio è quando l’endotelio dei vasi capillari non regola più bene la permeabilità delle proprie
membrane, per una qualche ragione, possiamo avere un accumulo di liquidi (EDEMA)
L’EPITELIO CUBICO SEMPLICE va a costituire il rivestimento dell’ovaio, la gonade femminile che ne è
rivestita da un unico strato di cell cubiche
GLI EPITELI CILINDRICI sono un mondo assai variegato in quanto possono presentarsi nella forma semplice
e nella forma pseudostratificata ed avere differenziazioni apicali sottoforma di microvilli e ciglia
Carrellata per il test:
Lamina semplice costituita da cell cilindriche tutte poggianti sulla memb basale, che entrano nel gioco del
passaggio di sost dal lume al connettivo e viceversa.
LUME DELL’INTESTINO TENUE: siamo in una porzione del ns app digerente dove si attua l’assorbimento
della maggior parte dei prodotti della digestione, che inizia nella bocca (BOLO) con la masticazione e la scissione
degli amidi per mezzo degli enzimi salivari, continua nello stomaco, con la digestione delle proteine grazie
all’acido cloridrico prodotto dalla mucosa gastrica (CHIMO) e si completa a livello di intestino tenue nella sua
prima parte, il duodeno, con la digestione dei grassi grazie alle secrezioni emulsionanti biliari e ai succhi
pancreatici riversati all’interno del lume intestinale (CHILO). Qui dobbiamo sicuramente avere un tessuto in cui
sia sviluppata al massimo la funzione di assorbimento, la parete del tenue non sarà dunque liscia ma rialzata in
pliche ad aumentare la superficie stessa del lume.
Se io taglio secondo una sezione sagittale ciascuna plica, che è chiamata VILLO INTESTINALE, vedrò che c’è
un asse di tessuto connettivo ricco di vasi, rivestito da una memb basale e da una epitelio cilindrico.
Le cell costituenti questo epitelio, mostrano al loro apice una linea o CUTICOLA COLORATA, che non è altro
che una serie di microvilli, che essendo come dimensioni al disotto della visione del microscopio ottico, appaiono
nelle preparazione istologica come una crestina colorata.
Siamo di fronte dunque ad una amplificazione dell’amplificazione: l’epitelio del tenue presenta villi che ne
amplificano la superficie assorbente i quali villi a loro volta sono tappezzati di cellule al cui apice sono presenti
In altri distretti, il rene per esempio, organo escretore per
microvilli che ne amplificano la capacità assorbente.
Microvilli
di uguale
altezza
Membrana basale
Cell cilindrica dotata
di microvilli
assorbenti
Villo
intestinale
eccellenza, nella parte distale del tubulo prossimale a funzione
di riassorbimento dell’acqua filtrata insieme al sangue, qui le
cell del tessuto epiteliale di assorbimento che ne rivestono il
lume non sono di uguale altezza, ma di dim diverse, tanto che
all’immagine ottica, sembra di vedere uno smerlo, da qui il
nome di ORLETTO
L’orletto dei tubuli renali, la cuticola striata dell’intestino
tenue, altro non sono che l’espressione al microscopio ottico di
microvilli deputati all’assorbmento o di nutrienti o di acqua
connettivo
Se ci spostiamo nell’app. genitale maschile, sezionando l’epididimo, che è il tubulo attraverso cui passa il liquido seminale
contenente gli spermatozoi che si sono formati a livello della gonade, vediamo che che la mucosa che lo riveste è costituita da cell
cilindriche e cubiche tutte impiantate sulla lamina basale: ci troviamo dunque di fronte ad un epitelio PSEUDOSTRATIFICATO
All’apice di ciascuna cellula vediamo delle strutture simili a pennacchi che i vecchi istologi, non disponendo di microscopi
elettronici, chiamarono STEREOCIGLIA, perché assomigliavano alle ciglia, ma non si muovevano. In realtà non sono ciglia, perché
non solo non si muovono, ma sono proprio prive della struttura ciliare, sono prive dell’ASSONEMA, sono quindi in definitiva
GROSSI MICROVILLI anche qui legati a fenomeni di riassorbimento del liquido seminale che transita nei tubuli
Quindi: STEREOCIGLIA,ORLETTI, CUTICOLA STRIATA, altro non sono che microvilli di grandezza diversa assestati
diversamente, destinati ad aumentare la superficie attiva del lume in cui si trovano e deputati all’assorbimento di un qualcosa che non
deve andare disperso, o eliminato.
GLI EPITELI SEMPLICI possono presentare anche ciglia. Prendiamo in considerazione la mucosa della tuba uterina.
In questa porzione dell’APP: GENITALE FEMMINILE transita la cell uovo prodotta dall’ovaio, prima di arrivare all’utero,
fecondata o meno ed essere accolta dall’ENDOMETRIO per una gravidanza o espulsa a seconda. In questo tratto troviamo
un’amplificazione in pieghe della superficie della mucosa che è costituita da un epitelio di cellule cilindriche che portano all’apice le
ciglia, strutture dotate di movimento, che muovono un secreto neutro di natura mucosa in cui si viene a trovare la cellula uovo che
transita verso la cavità uterina ed eventualmente gli spermatozoi che hanno risalito le vie genitali femminili, per andare a fecondare
l’ovocellula. Spostando questo secreto le ciglia aiutano nel loro movimento la cellula uovo e gli spermatozoi
EPITELIO PSEUDOSTRATIFICATO CON CIGLIA siamo nella trachea quel tratto dell’apparato
respiratorio costituito da un tubo di anelli cartilaginei, sempre beante per permettere il transito dell’aria, qui,
abbiamo un rivestimento del lume beante con un epitelio pseudostratificato formato da cell cilindriche e cubiche,
tutte impiantate sulla lamina basale, quelle cubiche incastrate e più basse, quelle cilindriche più alte con l’apice
dotato di ciglia che si affaccia nel lume tracheale. Queste ciglia toccano con il loro apice secreti di natura mucosa
(liquido appiccicoso filante), che imbriglia meccanicamente tutte le sostanze estranee che sono contenute nell’aria
che inaliamo (pulviscolo, polveri, pollini, batteri, antigeni etc.) questo allo scopo di ripulire al massimo l’aria che
dovrebbe arrivare agli alveoli polmonari (si è visto strutture fragili, dotate di pareti sottili, proprio per le loro
caratteristiche funzionali) il più pulita possibile. Questo passaggio dalle coane nasali attraverso la trachea verso i
bronchi per poi giungere agli alveoli è un percorso che l’aria fa per subire oltre ad un riscaldamento anche una
PULIZIA MUCOCILIARE. Questa si espleta con l’inglobamento delle particelle estranee per semplice adesione
fisica al muco o per adesione immunitaria, dovuta alla presenza di particolari molecole, chiamate anticorpi, che
vengono prodotte dall’organismo in difesa contro gli antigeni (sost. Estranee) e sono secrete a questo livello
dentro lo strato mucoso. Quando qualcosa sfugge a questa puizia iniziano i guai e ci ammaliamo.
Questo muco inglobante tutte queste schifezze deve essere allontanato, cosicché il battito ciliare sarà nelle parti
basse della trachea un movimento verso l’alto che spinge il muco nel retrobocca dal quale andrà direttamente
nell’esofago e sarà espulso attraverso l’apparato digerente oppure espettorato; nel tratto alto della trachea verso il
basso per subire il medesimo destino. Qualunque stimolo negativo (il freddo blocca il movimento ciliare con
ristagno di muco e schifezze e quindi ipersecrezione e pericolo di infezioni) in questo sistema si traduce in un
sovraccarico e quindi in una non completa pulizia dell’aria e in una sovraespressione di un sistema fisiologico
normale che si manifesta con TOSSE e RAFFREDDORE.
N.B. Il fumo non solo inibisce il movimento ciliare, creando noxe (corpi estranei) di muco e particelle
normalmente presenti nell’aria, ma sovraccarica di particelle inorganiche estranee (carta e carbone) l’aria
inspirata, con stimolo dei macrofagi (cell deputate alla difesa dagli agenti estranei) alla captazione e riduzione
della capacità di questi ultimi ad espletare ulteriore azione fagocitarla verso altri agenti infettivi. Si tratta di
particelle inorganiche che i lisososmi dei macrofagi non riescono a distruggere, così ne rimangono “intossicati”
riducendo le loro potenzialità
Il fumo fa ispessire la memb basale con riduzione degli scambi, inoltre in soggetti particolarmente predisposti i
processi infiammatori provocati dal fumo possono portare a fenomeni di degenerazione cellulare con l’insorgenza
di neoplasie dell’apparato respiratorio.
GLI EPITELI COMPOSTI Epitelio pavimentoso composto che si può presentare in due forme diverse con e
senza strato corneo a seconda della localizzazione i ambiente secco (l’EPIDERMIDE) o in ambiente umido. Sono
epitei formati da più starti di cell sovrapposti di cui solo il primo prende contatto con la memb basale assicurando
l’ancoraggio dell’epitelio al connettivo, questo strato è fatto di cell cubiche che costituiscono anche il
compartimento germinativo di tutto quanto l’epitelio. Al di sopra troviamo più strati di cell regolarmente
poliedriche che costituiscono lo strato intermedio che può essere da zona a zona più o meno spesso, come ultimo
strato troviamo cellule pavimentose che danno il nome all’epitelio. Eccezione fa l’epidermide nella quale
troviamo al di sopra dello strato pavimentoso un ulteriore strato di cellule morte dette squame cornee.
L’ESOFAGO e la CAVITA’VAGINALE sono organi in cui la funzione di rivestimento e protezione dalle lesioni
meccaniche viene espletata da questo tipo di epitelio, estremamente spesso e resistente. Qui essendo l’epitelio
situato in ambiente umido, non presenta squame cornee al di sopra dello strato pavimentoso e le cell di
qest’ultimo sono perfettamente funzionanti. In cavità vaginale le cellule dello strato pavimentoso presentano la
caratteristica di distaccarsi e vengono espulse con i fluidi vaginali, secondo un turn over fisiologico. Questa
caratteristica è sfruttata per l’effettuazione del pap test che serve per la diagnosi precoce del tumore della cervice.
ISTOLOGIA LEZIONE 5
GLI EPITELI PAVIMENTOSI COMPOSTI Epitelio pavimentoso composto che si può presentare in due
forme diverse con e senza strato corneo a seconda della localizzazione in ambiente secco (l’EPIDERMIDE) o in
ambiente umido. Sono epiteli formati da più starti di cell sovrapposti di cui solo il primo prende contatto con la
memb basale assicurando l’ancoraggio dell’epitelio al connettivo, questo strato è fatto di cell cubiche che
costituiscono anche il compartimento germinativo di tutto quanto l’epitelio. Al di sopra troviamo più strati di cell
regolarmente poliedriche che costituiscono lo strato intermedio che può essere da zona a zona più o meno spesso,
come ultimo strato troviamo cellule pavimentose che danno il nome all’epitelio. Eccezione fa l’epidermide nella
quale troviamo al di sopra dello strato pavimentoso un ulteriore strato di cellule morte dette squame cornee.
L’ESOFAGO e la CAVITA’VAGINALE sono organi in cui la funzione di rivestimento e protezione dalle lesioni
meccaniche viene espletata da questo tipo di epitelio, estremamente spesso e resistente. Qui essendo l’epitelio
situato in ambiente umido, non presenta squame cornee al di sopra dello strato pavimentoso e le cell di
qest’ultimo sono perfettamente funzionanti. In cavità vaginale le cellule dello strato pavimentoso presentano la
caratteristica di distaccarsi e vengono espulse con i fluidi vaginali, secondo un turn over fisiologico. Questa
caratteristica è sfruttata per l’effettuazione del pap test che serve per la diagnosi precoce del tumore della cervice.
La cute (strato corneo?), l’epidermide e il derma possono essere più spessi in alcune zone del corpo, in quelle
zone esposte a maggior stimolo meccanico e la porzione ispessita oltre che allo strato epidermico è riferita anche
allo strato corneo.
Considerando le varie funzioni dell’epidermide, ripetiamo come tra derma e tessuto connettivo sottostante, ove gli
stimoli meccanici sono più forti, le giunzioni si presentino come papille(?)
La composizione cellulare dell’epidermide è varia :
1.
CHERATOCITI
2.
MELANOCITI
3.
CELLULE DI LANGHERANS
4.
CELLULE DI MERKEL
Sono 4 popolazioni cellulari che collaborano a portare a termine le funzioni dell’epidermide.
I cheratociti sono le cellule vere e proprie dell’epidermide, che dalla formazione nel compartimento germinativo
risalgono i vari strati, spostandosi dallo strato basale al pavimentoso portando avanti la cosiddetta
CITOMORFOSI CORNEA, differenziandosi da cellule vitali in squame cornee cadendo poi sotto la spinta delle
nuve cellule che si formano e risalgono gli strati.
CITOMORFOSI CORNA:
STRATO BASALE CUBICO SPINTE DALLE CELLULE CHE VIA VIA SI DIFFERENZIANO MUTANO
DI FORNMA SI RIEMPIONO DI CHERATINA, PERDONO LE CARATTERISTICHE TIPICHE
CELLULARI, SI STACCANO DALLE GIUNZIONI (CHERATOCINICHE?) E COME SQUAME CORNEE
RIMANGONO ADESE UN PO’ ALL’EPIDERMIDE E POI CADONO.
Il tempo di spostamento dallo strato a quello corneo è di circa 20/30 gg, questo è il rinnovo fisiologico
dell’epidermide.
Lo strato basale, cellule tute poggianti sulla memb basale, cellule tipiche da cui partono le divisioni in cui una
cellula resta in loco e l’altra perde il contatto con la memb basale rompendo le giunzioni (?) e comincia ad essere
spinta negli strati sovrastanti cominciando il suo percorso per diventare squama cornea..
Quali sono le manifestazioni di questo differenziamento?
Già nello strato basale le cellule destinate a spostarsi e cominciare la differenziazione perdono i connotati di
cellule indifferenziate e accumulano nel proprio citoplasma i TONOFILAMENTI (sono filamenti intermedi delle
cellule epiteliali costruiti con una proteina che si può presentare sotto forme diverse, che è la cheratina.
Qui troviamo i cheratociti cubici basali che cominciano ad elaborare questa proteina sottoforma di tonofilamenti e
comincianp ad accumularla nel citoplasma. Perdono l’attacco alla memb basale e questo vuol dire salire perché
altre cellule le spingono da sotto, passono così nello
STRATO SPINOSO, un insieme di strati sovrapposti dove l’epidermide è più spessa, gli strati sono più
numerosi.
Nello strato spinoso i cheratociti diventano poliedrici e non hanno più la superficie liscia, ma con tante
protuberanze che vengono a contatto con lembi simili di citoplasma tramite desmosomi.
I cheratociti, cambiano forma assumendo queste spine quindi sono le cell ad essere spinose, si crea così tra queste
cellule che sono connesse con desmosomi, che sono le giunzioni tipiche meccaniche di ancoraggio verso cui i
tonofilamenti si dirigono e quindi è un sistema sinergico ed elastico che ammortizza i vari urti meccanici che
vengono prodotti sulla superficie dell’epidermide.
Si crea un labirinto tra le cell che permette una migliore permeabilità dello strato, visto che la sorgente di ossigeno
e il metabolismo stanno nel connettivo. Questo è uno strato a volte molto spesso che facilitano il fluire dei fluidi
biologici che portano ossigeno e metabolici per un migliore trofismo epidermico.
Mentre risalgono lo strato spinoso, aumenta il tono dei tonofilamenti che si appiccicano tra di loro tramite una
proteina a formare strutture più grosse di diametro dette TONOFIBRILLE. La cheratina, continua ad accumularsi
in queste cellule e le proteine di membrana vengono prodotte e appiccicate sul versante citoplasmatico della
membrana a riforzarlo e a limitarle la capacità di scambio, inoltre dei granuli, contenenti lipidi, vengono fabbricati
dall’apparato di Golgi e hanno grande ruolo nell’impermeabilazzazione dell’epidermide.
Così si sale di strato in strato e continua il differenziamento finché la cellula tende a diventare piatta assumendo,
la caratteristica forma apavimentsa, siamo arrivati al cosiddetto
STRATO GRANULOSO, i cheratociti, ricchi ormai di tonofibrille, verniciano queste strutture a mo’ di filamenti
grossi e spessi con un'altra proteina che hanno fabbricato (testino cina?), che è un ulteriore appoggio alla cheratina
e rende la squama molto resistente, ma morbida. E’ tipico questo strato all’epidermide, mentre nelle unghie sarà
cheratina dura perché gli manca l’apporto di questi granuli.
I granuli lamellari, ricchi di lipidi, vengono esogitati nello spazio tra le cellule che tende a chiudersi, non ci sono
piu spine, le cell sono mattonelline espanse in superficie sottili e robusti che chiudono gli spazi tra di loro con
giunzioni occludenti, quindi si limita qualsiasi tipo di passaggio di sostanza e si depista tutto con i lipidi.
L’epidermide a questo punto è impermeabile. Scambia ma non fa perdere i liquidi, ormai si catena una serie di
eventi che fanno decidere alla cellula di andare in apoptosi, perdono il nnucleo gli organuli non si rinnova piu
niente quale cornea che perdura nello strato corneo con elemento sinergico con altri,verniciato dai lipidi negli
striti piu bassi ma poi libero da ogni legame(anche i desmosomi o le giunzioni si sono perse perche non piu
rinnovate).Tende a cadere con strofinamenti e o varie forze che si hanno in superficie.Dallo strato basale le
cellule cubiche si passa adese alla membrana, sono le cellule del compartimento germinativo che garantiscono
l’adesione al resto dell’organismo.Alcune cellule cominciano a differenziarsi e vi accumulano i
tonofilamenti,salgono,diventano spinose,i tonofilamenti si ispessiscono in tonofibrille,i granuli lamellari.
Si sale ancora e si arriva allo strato granuloso in cui le tonofibrille vengono verniciate con la cherototina? che
rende la squama robusta ma morbida per le caratteristiche fisiche di questo organo.I granuli lamellari vengono
esocitati negli spazi ormai chiusi,giunzioni occludenti e si va in apoptosi;i nuclei spariscono,gli organuli se ne
vanno, ormai questa da cellula è diventata squama cornea, robusta anche per il fatto che come una corazza ad
opera di proteine specifiche si sono depositate lungo tutte le superfici delle membrane dalla parte citoplasmatica,
irrobustendola.La squama cornea fa tutto uno con l’altro costituente del proprio strato, poi mano a mano si isola e
tende a cadere ormai priva dei connotati cellulari.( desquamazione).
La visione istologica mostra come lo strato basale ha una caratteristica in più rispetto a quelle fin ora
descritte:cioè cellule dello strato basale contengono granuli di melanina, un pigmento, quindi ha le caratteristiche
di incluso che non viene introdotto dai cheratinociti ma da cellule diverse, i melanociti che concedono questi
granuli alle cellule vicine, i cheratinociti, conferendo all’epidermide la capacità di impedire il passaggio dei raggi
U.V. che sono mutageni e quindi potenzialmente capaci di dar vita a mutazioni non opportune all’organismo
come:neoplasie.
I granuli di melanina sono alla base di un meccanismo di difesa dei raggi U.V.Difesa naturale che si enfatizza con
l’esposizione al sole.
MELANOCITI:
Sono cellule localizzate nello strato basale che si inseriscono tra un granulocita e l’altro, vicini.Non sono uniti da
giunzioni con granulociti vicini, hanno solo rapporto di contatto.
Unità melanica è l’insieme di un melanocita e di un cheratinocita a cui cede granuli di melanina che via via
sintetizza.E’ il melanocita che sintetizza i granuli.Man mano che il granulocita viene ad avere i granauli dal
melanocita vicino, esso se ne serve per difesa come schermo ai raggi solari.Maggiore esposizione dà maggior
produzione.Man mano che il cheratocita sale,i lisosomi demoliscono la melanina perché è importante che questa
difesa sia negli strati più bassi che difendono il connettivo sottostante e le cellule del compartimento germinativo,
che è quello che deve rimanere integro.
Man mano che si sale, queste cellule demoliscono granuli di melanina,la squama cornea non ne ha quasi più, ma
caratteristiche razziali diverse, fa si che ci siano individui che possono mantenere negli strati più superficiali,
granuli di melanina, quindi la pelle è più pigmentata.
Ricordarsi quindi che questi granuli vengono prodotti dai melanociti, cellule diverse dai cheratinociti,dai
melanociti vengono passate ai cheratinociti che man mano che si differenziano demoliscono i granuli di melanina.
LANGERANS:
La nostra epidermide possiede queste cellule che sono localizzate un po’ più in alto rispetto al basale,nello strato
spinoso.Sono cellule che si insinuano tra i cheratinociti vicini e hanno il significato di cellule di difesa.Attraverso
la superficie corporea possono entrare sostanze strane quindi queste cellule fanno una prima difesa.Fanno parte
del sistema dei macrofagi.
Qual è la loro funzione? Qualsiasi sostanza entra nell’epidermide, loro la riconoscono come estranea, ma se la
lasciassero passare molto probabilmente il nostro sistema di difesa non la riconoscerebbe,allora la captano,la
portano dentro nel proprio citoplasma,le complessano ad altre cose come antigeni di istocompatibilità,la
dispongono come recettore sulla propria membrana e così la presentano,migrando verso le zone giuste, i linfociti,
facendole riconoscere e quindi per questo fanno parte, all’interno dei macrofagi, delle cosiddette “cellule
presentanti all’antigene”,intendendosi per antigeni una sostanza estranea e per cellula presentante la cellula che li
riconosce,la rielabora e la respingono sulla membrana poiché il linfocita se ne accorga.Quindi è un meccanismo di
difesa.Le cellule di langerans come prima barriera difensiva
MERKEL:
Sono cellule immerse nello stato basale ai cheratinociti, ai melanociti, etc….
Hanno la caratteristica di essere uniti con desmosomi alle cellule vicine,cioè ai cheratinociti, poi vengono in
stretto contatto con una terminazione nervosa che a mò di coppa supera la membrana basale e va, così,sotto la
cellula di Merkel.La cellula di Merkel,quindi,che è legata ai cheratinociti,subisce delle deformazioni uguali a
quelle dei cheratinociti che vengono sollecitati in superficie.Questa sollecitazione meccanica porta questa cellula a
produrre delle sostanze che riescono ad eccitare la fibra nervosa producendo una corrente elettrica, ma
depolarizzando la membrana, che corre fino ai centri e ci informa del tutto.Ecco perché queste cellule sono più
numerose in zone come i polpastrelli,contorno delle labbra.
Cellule Merkel come recettori tattili:sta nello strato basale e stà unita alle giunzioni desmosomiche dei
cheratinociti vicini, è in stretto contatto per vicinanza di membrana, con una terminazione nervosa che proviene
dai centri, supera la mamb basale e a mo’ di coppa si slarga e sostiene da vicino le cell di merkel. Recepisce la
deformazione meccanica, la fibra nervosa porta una corrente elettrica che è una conoscenza ai centri.
La cell di merkel percepisce lo stimolo /deformazione dell’epidermide e produce una sostanza che stimola la fiba
nervosa che porta l’impulso.
GUARIGIONE DI UNA FERITA
Per ferita si intende un danno che viene portato solo all’epidermide a memb basale intatta. Quando si ha questo
tpo di ferita, le cell si portano ai margini dell’epidermide laddove si è rotta la continuità e si dispongono a
ripristinare lo strato basale che è quello importante; da lì le divisioni ricostruiscono tutto lo spessore
dell’epidermide.
EPITELI DI TRANSIZIONE
Composto da due strati sovrapposi uno di cell basali dove oltre alle cell cubiche del compart. Germinat. Ce ne
sono altre più alte a clava, al di sopra ci si localizzano le cell ad ombrello, perché viste isolate mostrano un
margine a cupola sono bi o tetranucleate per plasmodio, sono cell grandi che possono ricoprire parecchie cellule
sottostanti, e queste sottostanti a clava hanno una parte più slargata, che prende relazione con le stecchettature
delle cellule ad ombrello, quindi immaginiamo un epitelio di superficie che ha cell cubiche con compartimento
germinativo, cell a calava pggianti slla memb basale e cell ad ombrello
Questo epitelio è detto di transizione perché riesce a passare da questa forma rilassata della parete a cui fa da
rivestimento ad una forma stretta e ciò è dovuto alla necessità dell’epidermide che riveste internamente l’org cavo
(p.e. la vescica) a seguire la distensione e queste cell a clava che erano alte si abbassano e diventao cubiche le cell
ad ombrello si appiattiscono e ricoprono molte cell di sotto e l’epitelio si è abbassato progressivamente seguendo
quest transizione della <parete dell’organo.
Le cell ad ombrello oltre ad appiattirsi hanno un meccanismo di membrana che gli permette un ulteriore
possibilità di distensione e quindi bisogna pensare all’ombrello quando la parete è rilassata con tante riserve di
membrana come delle pieghettature che aprendosi permettano delle grandi distensioni di superficie.
Queste membrane sono poco fluide perché queste cell devono garantire un certo grado di impermealizzazione.
Ecco quindi che si chiudono gli spazi tra le cell con giunzioni occludenti, rinforzando la propria membrana e
impedendo che l’acqua venga richiamata nel lume dlel’organo
E’ un epitelio impermeabile.
EPITELI GHIANDOLARI
Vengono definiti come epiteli specializzati nella elaborazione e secrezione di sostanze secreti a vantaggio
dell’organismo.
Si distinguono su base istologica, anatomica e nunzio nale due tipi di epiteli ghiandolari:
ESOCRINO ed ENDOCRINO
EPITELIO GHIANDOLARE ESOCRINO strutture epiteliali specializzate nella sintesi e nel rilascio di sostanzr
che vengono riversate o nella superficie de corpo o in cavità in esso contenute e questi secreti agiscono laddove
vengono riversati.
Dal punto di vista strutturale si distinguono in:
GHIANDOLE ESOCRINE UNICELLULARI E PLURICELLULARI.
Quelle unicellulari sono immaginabili come cell distribuita dove più dove meno, nel contesto dell’epitelio di
rivestimento o delle mucose, quindi nelle mucose ci possono essere queste ghiandole che possono produrre
secreto, muco, che è appiccicoso filante, viscososo che una volta riversato, si stratifica sulla superf. Dell’epitelio
con vario significato biologico, andando ad accumularsi col secreto di ghiandole pluricell, che verseranno anche
loro i loro prodotti in quelle sedi.
Il muco è costituito da vari elementi, ma la componente specifica è una famiglia di glicoproteine, quindi ci sarà un
percorso dal reticolo endoplasmatico all’app. di Golgi ai granuli di secrezione e tramite processo di esocitosi al
riversamento del prodotto al di fuori della memb cell.
Ecco quindi come si spiega il nome derivante dalla forma queste cellule disseminate nel contesto dell’epitelio di
rivestimento delle mucosae cilindriche sono, quando scariche , cilindriche, ma accumulando prodotto di
secrezione nella parte più superficiale (al di sopra del nucleo e del Golgi) tendono a far dilatare questa porzione,
facendo assomigliare la cell ad un calice, dove nella teca la parete più larga viene immagazzinato il secreto prima
dell’esocitosi, lo stelo, parte più stretta, dove c’è nucleo e maggior parte degli organuli ed infine il piede del calice
che poggia sulla membrana basale.
Nell’intestino possiamo vedere la lamina di rivestimento cilindrico semplice, la cellula mucosa che riversa il suo
prodotto all’apice partecipando al fenomeno di assorbimento protegge l’apice cellulare, “inposta” le scorie di
questo processo.
Nella trachea, c’è strato pseudostratificato e il muco forma un mantello sul quale vengono inglobate tutte le
particelle estranee respirate che però non devono arrivare agli alveoli polmonari.In trachea si vengono a
localizzare anche anticorpi specifici che si tuffano in questo muco per neutralizzare eventuali noxe infettive.
Sull’epitelio della trachea sono presenti ciglia che spostano questo muco favorendone la deglutizione, nel
digerente o l’espettorazione.Il freddo e il fumo immobilizzano le ciglia con conseguente accumulo del muco in
loco.
Nella tuba ovarica c’è epitelio cilindrico semplice con ciglia e sono presenti anche cellule che sembrano calici e
secernano secreto adatto a creare l’ambiente per la cellula uovo.
Quando ci imbattiamo in epiteli di rivestimento di tipo cilindrico in cui tutte le cellule della lamina epiteliale di
rivestimento vanno a secernere il muco parliamo di superficie secernente(per esempio il rivestimento interno dello
stomaco)
GHIANDOLE ESOCRINE PLURICELLULARI: Vanno a costituire delle strutture anatomicamente
riconoscibili.
Durante lo sviluppo delle lamine di rivestimento l’epitelio di superficie va a proliferare creando una specie di
fittone o rdice che si affonda nel connettivo sottostante.Il fittone ed il connettivo sono sempre separati dalla
membrana basale.Questa calata di cellule si approfondisce sempre di più e poi si cavita? Formando una specie di
introflessione che si mantiene in connessione con l’epitelio che l’ha originata.Nasce così la ghiandola esocrina.
Il ghiandolare esocrino rivesa sulla superficie degli epiteli di rivestimento il proprio secreto attraverso il dotto
escretore attraversando l’epitelio che l’ha generata.
Nella ghiandola esocrina posso distinguere 2 porzioni:
ADENOMERO:dato da cellule che elaborano il secreto e lo riversano nel proprio lume.Da questo il
secreto viene spinto attraverso
UN CONDOTTO (DOTTO ESCRETORE): per poi essere riversate sulla superficie dell’epitelio da dove
la ghiandola si è originata
Le ghiandole esocrine dell’organismo sono molte,annesse a quasi tutti gli apparati.
Le ghiandole possono essere classificate anche secondo un criterio topografico che è a cavallo tra istologia e
anatomia microscopica, definendole in:
Intraparietali o intramurali
Extraparietali o extramurali
Le cellule per prolificazione dell’epitelio di rivestimento si approfondiscono nel connettivo sottostante e si
possono fermare o altrimenti andare oltre. Alcune cellule possono addirittura fuoriuscire dall’epitelio da dove si
sono formate, ecco quindi le ghiandole intra- ed extraparietali.
Tutte però restano in contatto con l’epitelio da cui si sono formate (un esempio è il pancreas).
Altro criterio è quello morfologico: a seconda della forma dell’adenomero:
Tubulare : caratterizzata da un estroflessione a dito di guanto giù nel connettivo di cui è molto difficile
distinguere l’adenomero dal dotto escretore.
Alveolare: presenta un adenomero molto slargato in cui c’è un lume altrettanto slargato che può servire da
serbatoio per il secreto
Acinosa: assomiglia ad un chicco d’uva, ha un lume talmente piccolo che qualche volta non è visibile. In
alcuni casi come le ghiandole salivari maggiori si parla di tuba acinosa, perché sotto varie forme l’ultima
porzionedei dotti e l’adenomero che di sotto ha forma acinosa, hanno entrambe capacità secretiva.
Tubuloacinosa: l’ultima parte dei dotti che sono tubuli, può avere funzione secernente e unire il proprio
prodotto a quello dell’adenomero che ha forma acinosa
Altro criterio di classificazione in base alla forma del dotto escretore
Ghiandola semplice: il dotto escretore è uno e porta il secreto all’esterno da un singolo
adenomero
Ghiandola ramificata: il dotto escretore raccoglie il secreto di più adenomeri
Ghiandola composta: il dotto principale( detto COLLETTORE) si ramifica più voltefinché gli
ultimi dotti i più piccoli, detti PRETERMINALI prendono rapporto con gli adeomeri.
Altra classificazione secondo criteri morfofunzionali , per la modalità di emissione del secreto.
MEROCRINA: il secreto elaborato viene riversato nel lume per poi prendere la via del dotto escretore.
Questo secreto può fuoriuscire dalla cell per esocitosi oppure se è un secreto idrosalino attraverso la membrana
APOCRINA: il secreto si immagazzina all’apice della cell produttrice, così essa diventa sempre più alta
da cubica , diventa cilindrica, poi strozza il proprio citoplasma al di sotto della zona in cui si trova il secreto. Si
strozza fino a rompersi, il pezzo di cell cade nel lume, quindi il secreto è fatto dal prodotto di secrezione e parte
del citoplasma della cell. La cell secernente ripristina il suo apice tra un ciclo di secrezione e l’altro.
OLOCRINA: dal greco “olos” = tutto, la cellul si differenzia tutta in prodotto di secrezione e viene
gettatafuoi attraverso il dotto escretore. Le cell si dividono, una resta in loco, l’altra si differenzia, iniziando ad
immagazzinare il secreto all’interno del suo citoplasma fino a saturarsene ad entrare in apoptosi, perde così gli
organuli, il nucleo e finisce nel dotto escretore.
Nello stomaco ci sono delle ghiandole tubulari semplici, ch non sono altro che introflessioni digitiformi della
parete gastrica, dell’epitelio di superficie nel connettivo.
Il secreto risale e si stratifica in superficie.
Quando la ghiandola è composta arriva a diventare un vero e proprio organo, con un parenchima (tessuto
epiteliale stratificato e organizzato per certe funzioni) specializzato nella secrezione e in uno stroma che è il
connettivo con funzione trofica e di conduzione degli stimoli nervosi, attraverso il quale passano i dotti escretori e
che divide la ghiandola in lobi e lobuli.
Ancora un altro criterio e in base alla natura chimica del secreto:
Sierosa: e costitutita da proteine è limpida e trasparente( es digestivi del pancreas)
Muco : secrezione glicoproteica (s.: cell mucosa) misto siero mucoso (salivari ,sottolinguali) idrosalino
(sudore) lipidico (sebacea)
Secreti complesi (latte) glicoproteica lipidica, proteica, anticorpi, acqua, vitamine
Le cell dell’adenomero producono secreto, lo riersano nel lume e lo spingono nel dotto, che può essere anche
lungo, verso la superficie per mezzo della contrazione di una cell mioepiteliale che spreme l’adenomero e i primi
tratti del dotto.
Queste cell mioepiteliali di natura epiteliale, localizzate tra le cell dell’adenomero e la memb basale, hanno
prolungamenti, hanno sviluppato contrattile. Sono una via di mezzo tra cell epiteliale e cell muscolare.
Circondano ladenomero, ogni tanto si contraggono e spingono il secreto.
GHIANDOLE ANNESSE ALLA CUTE
SUDORIPARE: sono presenti in tutto il corpo e sono glomerulari (tubulari) semplici, quindi durante lo sviluppo
si formano per introflessione dell’epidermide nel connettivo. La ghiandola rimane nel connettivo, cresce ei l
tubulo si attorciglia su se stesso. E’ ecrina, perché il sudore è idrosalino; i Sali portati fuori dalla superficie cell
dalle pompe, il lume chiama acqua che fuoriesce dalla cell, entra nel lume e forma il secreto (sudore). Il sudore è
su tutto il corpo, nelle ascelle, però oltre a queste cell ci sono altre più grosse, più ramificate, che hanno un secreto
più denso con alcune componenti glicoproteiche che va a mescolarsi al secreto idrosalino dando quella secrezione
particolare delle ascelle. Le ghiandole delle ascelle sono apocrife.
SEBACEE: sono annesse ai peli. Il sebo è come una crema lipidica naturale che si stratifica sulla pelle a
protezione. Sono ghiandole ramificate acinose, a secrezione olocrina lipidica. Hanno breve dotto escretore che
immette nel camnale del pelo e questo secreto viene spinto ed emerge dalla porzione basale del pelo e si stratifica
sulla pelle. Queste ghiandole sono sotto l’influsso ormonale.
MAMMARIE DI ALLATTAMENTO solo nell’allattamento, questa ghiandola è l’unica ad avere struttura
composta a secrezione complessa dove la parte lipidica viene immessa per apocrinia. Durante la gravidanza e
subito dopo il parto, gi ormoni stimolano la crescita di questa ghiandola che altrimenti è un insieme di piccoli
dotti a fondo cieco, tutti emergenti nella zona del capezzolo. Con la gravidanza diventa alveolare e l’alveolo con i
suoi lumi dilatati diventa serbatoio della secrezione.
CERUMINOSE(apocrina):simili alle sudoripare presenti nel canale dell’orecchio.
GHIANDOLE ENDOCRINE:
Riversano il loro prodotto nel circolo del sangue e qualche volta nel circolo linfatico. Gli ormoni prendono la via
del circolo e vanno a colpire i cosiddetti organi/cellule bersaglio, che hanno i recettori specifici per quel
dterminato ormone e poi daranno risposta. L’ormone puo anche agire sulla cellula che lo produce se ne ha i
rispettivi recettori oppure su cellule vicine.
Quali sono le strutture capaci di elaborare ormoni:
L’epitelio puo formare ghiandole endocrine
Il connettivo puo formare ghiandole endocrine dette interstiziali.
In vari tessuti e organi che portano avanti altre funzioni
Le ghiandole endocrine epiteliali possono essere unicellulari e costituire il cosiddetto sistemo endocrino diffuso,
cioè cell capaci di produrre ormoni nel contesto di lamina epiteliale di rivestimento o ghiandolari.
L’apparato respiratorio e digerente hanno un rivestimento che oltre a protare ghiandole unicellulari esocrine
hanno ghiandole endocrine.
Ghiandole epiteliali sono:
TIROIDE
SURRENALE
PAROTIDI
IPOFISI
IPIFISI
MEPOFISI(?)
PORZIONE ENDOCRINA DEL PANCREAS
L’epitelio che da origina alle ghiandole si puo organizzare o sotto forma del cordone cellulare(fine di cellule che
fiancheggiano i capillari sinusoidi in cui l’ormone viene riversato oppure dare origine a follicoli, strutture chiuse
rotonde nel ci lume puo essere messo il secreto, che ha significato di preormone(tiroide), sacche laterali alla
trachea in cui s’immagazzina il preormone tiroideo chiamato colloide.
La celll del follicolo riassorbe parte del colloide e lo rielabora, forma gli ormoni tiroidei e li rimette in circolo per
mezzo dei capillari sinusoidi che stanno in mezzo ai follicoli. La colloide è una glicoproteina lipidica.
Gli ormoni possono essere di varia natura: proteici, glicoproteici, lipidici(ormone steroideo).
Le ghiandole interstiziali sono localizzate nell’interstizio delle gonadi(ovaio e testicolo)e sono cell del connettivo
tra strutture proprie delle gonadi, producono ormoni estrogeni nella donna e testosterone nell’uomo.
Attività endocrina in organi che portano avanti altre funzioni:
rene, cuore, placenta, cell adipose e neurone
NEUROEPITELIO(Detti anche epitelio sensoriali)
Cell epiteliali che recepiscono stimolo e sono in gradi di stimolare terminazioni nervose per portare un
informazione. Sono epiteli in grado di evocare sensazioni. Il gusto è fatto da cell neuroepiteliali.
ISTOLOGIA:VI LEZIONE
TESSUTO CONNETTIVO
DEFINIZIONE: insieme di tessuti accomunati dall'organizzazione morfologica e dalla origine embriologica.
STRUTTURA: tutti i tessuti connettivi sono costituiti da cellule e dalla sostanza intercellulare. Quest'ultima
chiamata anche matrice ed e' suddivisibile in due strutture diverse nominate sostanza fondamentale anista o
amorfa e fibre.
Dal punto di vista della derivazione embriologica, i connettivi derivano tutti dal mesenchima.
Andiamo a trattare i connettivi dalla sostanza fondamentale anista detta anche amorfa perché non ha una
morfologia descrivibile al microscopio. Di solito, essendo molto ricca di acqua, viene solubilizzata e nei preparati
scompare. La potremmo descrivere come una gelatina, un gel, a vario grado di viscosità, più o meno densa, e
quindi più o meno resistente elasticamente alla compressione. La sostanza fondamentale amorfa, come si e'
detto,è costituita da acqua in cui sono disciolti elettroliti, proteine e precursori delle macromolecole e poi e'
caratterizzata da tre categorie di sostanze macromolecolari che sono i glicosamminoglicani(GAG), i proteoglicani
e glicoproteine.
I GAG sono dei carboidrati, sono delle lunghe catene polisaccaridiche composte dal ripetersi di disaccaridi. Il più
conosciuto tra i gag e' l'ACIDO IALURONICO, un glicosaminoglicano quasi ubiquitario perché si trova in quasi
in tutti i tessuti connettivi nella sostanza fondamentale anista. Di solito i gag, ad eccezione dell'acido ialuronico
che sta da solo, tendono a legarsi su assi di natura polipeptidica, quindi proteici, come fanno le setole in un
spazzolino. Quindi se già i gag sono delle grosse molecole, se si uniscono sopra l'asse proteico a formare i
proteoglicani, sono edifici molecolari estremamente grandi. Edifici molecolari, sia i gag che i proteoglicani,
capaci di legare acqua e di tenerla in equilibrio con l'acqua non legata creando così una specie di movimento dell’
acqua attraverso la sostanza fondamentale anista. questo sta alla base della funzione trofica di questa parte della
sostanza intercellulare.
Abbiamo detto più volte che i connettivi portano il trofismo di tutti quanti gli altri tessuti poiché nel proprio
contesto portano i vasi, i vasi portano i metaboliti e l'ossigeno, questi varicano la parete endoteliale, passano il
connettivo e, attraverso la sostanza fondamentale anista, vengono avvicinati alle cellule o agli altri tessuti e poi
allontanati i prodotti di scarto e l'anidride carbonica. E’ attraverso quindi questo gioco che i gag e i proteoglicani
compiono, legando l'acqua e lasciandola non legata e quindi con tutto ciò che nell'acqua si e' disciolto, si ha
questa diffusione delle sostanze trofiche del connettivo.
Hanno anche un ruolo di difesa perché controllano questo meccanismo e quindi sono in grado di evitare il
passaggio di alcune sostanze o di facilitarlo. Hanno inoltre un ruolo meccanico perché rendono la sostanza
fondamentale anista plastica, soprattutto resistente alla compressione. Se si tocca un connettivo giovane con la
sostanza fondamentale anista (ha una conformazione molto precisa) sentiamo che offre una resistenza, mentre ma
man mano che il connettivo sottostante l’epidermide (il derma) invecchia diventa meno resistente a questi
fenomeni di compressione.
Glicoproteine sono lunghe molecole polipeptidiche a cui si legano degli oligosaccaridi (sono dei carboidrati),
anche qui come le setole di uno spazzolino ma e' da notare che in questo caso la componente proteica e'
preponderante su quella carboidratica. Le glicoproteine hanno soprattutto la capacità di raccordare le molecole
della matrice alle varie popolazioni cellulari del connettivo, cioè ancorano riconoscendo proteine di membrana
delle cellule (ancorano queste cellule alla sostanza intercellulare) di cui il connettivo e' costituito oppure possono
avere il ruolo di ancorare a livello delle membrane basali le porzioni non connettivali al connettivo. Quindi
sempre funzione di raccordo, di legame. Importante da questo punto di vista saranno la laminina e la fibronectina.
Le fibre del connettivo sono di 3 tipi collagene,elastiche e reticolare.
Cosa si vuole sapere delle fibre? e' importante sapere le caratteristiche fisiche e chimiche.
Le fibre collagene sono dei cavi inestensibili e resistenti a forze che tendono a deformarle fino a che non si superi
il 2% o i 500 Kg per cm cubo. Questo darà caratteristiche ai tessuti ricche di fibre collagene. Quindi queste fibre
al più si rompono ma non si deformano. Ma sono flessibili. Sono dette bianche perché conferiscono questo colore
al tessuto a fresco, con la bollitura diventano gelatina da cui il nome collagene. Hanno enzimi comuni e possono
avere spessore variabile e sono costituite da una molecola che si chiama tropocollagene o collagene, possono
avere diametro diverso perché sono il risultato dell'assemblaggio di più microfibrille (più ce ne metti lateralmente,
più diventa grossa la fibra collagene).
Noi abbiamo cellule del connettivo che fabbricano questa molecola che si chiama tropocollagene o collagene
fatta da 3 catene con assone a tre capi che la rende più flessibile e meno estensibile quindi più rsistente. Questa da
origine, assemblandosi lateralmente con altre, a delle microfibrille. Più microfibrille si mettono insieme a mazzo e
formano la fibra collagene (quindi può essere più o meno spessa di diametro).
Di fatto il tropocollagene e' una famiglia di molecole, ne hanno decritti fino a 18-20 tipi numerati con numeri
romani.
Il risultato finale di molecole di tropocollagene diverse è diverso:
Per le fibre collagene di solito partecipa il collagene di tipo I che da strutture molte robuste,molte grosse.
Le fibre reticolari (che sono il II tipo di fibre del connettivo) sono costituite invece da tropocollagene di tipo III.
Il risultato e' di fibrille sottili in estensibili, perché e' tropocollagene, ma pochissimo resistenti (se si tirano si
rompono più facilmente delle fibre collagene) e danno origine delle reti, da qui il nome, reticolati che possono
svilupparsi in un piano e danno origine come esempio a una rete di recinzione che si spande in un piano.
Altrimenti come una giungla cioè un reticolo tridimensionale.
Le fibre elastiche sono estensibili, se applico una forza le deformo, ma elastiche,finita la forza ritornano alla
primitiva lunghezza. Sono gialle perché conferiscono questo colore ai tessuti che ne sono ricchi, resistono alla
bollitura e sono digerite da enzimi specifici quali la elastasi. Queste fibre elastiche hanno una doppia costituzione
sono costituite da microfibrille di fibrillina. Queste fibrille di fibrillina, una glicoproteina, si mettono una accanto
alle altre. Successivamente, durante la loro formazione, viene dimezzato, tra le fibrille di fibrillina, l'elastina che
non ha una conformazione propria e' più amorfa ma che conferisce l'elasticità' alle fibre quindi si deformano e poi
ritornano come prima.
Quindi tre tipi di fibre di cui è impostante ricordare le caratteristiche fisiche e da che cosa sono costituite.
Le cellule del tessuto connettivo derivano dunque dalle cellule mesenchimali, ma la cellula mesenchimale può
avere un comportamento diverso:
• può dare origine direttamente a cellule del connettivo attraverso il differenziamento e
rimanere nel contesto come compartimento germinativo
• o precocemente durante lo sviluppo andare a popolare il midollo emopoietico, quello
rosso, quello che da origine alle linee del sangue, e lì dare origine a un precursore di
queste cellule, poi da questo precursore midollare avranno origine i globuli bianchi (i quali
in parte attraverseranno la parete endoteliale e andranno a differenziarsi in cellule del
connettivo).
Quindi cellule che derivano direttamente dal mesenchima e cellule che derivano da precursori midollari o
precursori ematici,indicano che derivano dalla cellula mesenchimale attraverso un percorso diverso, o
direttamente nel tessuto o attraverso prima una tappa nel midollo e li danno origine a popolazioni cellulari
midollari, e poi da queste le varie categorie dei leucociti che in parte attraverseranno l'endotelio e andranno nel
connettivo a differenziarsi.
Questa è la cellula mesenchimale che presenta molte protuberanze, prolungamenti, è una cellula poco
differenziata che tende ad unirsi con altre a formare il CITORETICOLO per mezzo di giunzioni comunicanti.
Nel mesenchima tra le cellule c'e un liquido tissutale che diventerà sostanza intercellulare solo nei tessuti
connettivi adulti.
CLASSIFICAZIONE DEI TESSUTI CONNETTIVI:
Su base funzionale:
٭Tessuti connettivi propriamente detti
٭Tessuti connettivi di sostegno :
Cartilagine e Tessuto Osseo
٭Tessuti connettivi a funzione trofica:
Sangue e la Linfa (noi faremo solo il sangue)
TESSUTI CONNETTIVI PROPRIAMENTE DETTI
Sono dei tessuti connettivi costituiti da cellule e sostanza intercellulare con le caratteristiche dette fino adesso che
porteranno funzioni meccaniche,trofiche, cicatriziali e di difesa.
Spendiamo qualche parola per spiegare: il tessuto connettivo deve questo nome al fatto che i primitivi anatomici
trovarono che era un tessuto che connetteva varie parti di un organo,e i vari organi tra di loro (viene da qui il
nome). Ha una funzione meccanica perché i connettivi avvolgono gli organi formando delle capsule che
ammortizzano i vari urti. Pensiamo al tessuto connettivo sottocutaneo che ammortizza gli stress portati in
superficie. I tendini che legano la muscolatura alle ossa. sono tutti tessuti connettivi propriamente detti che
portano avanti a vario titolo funzione meccanica.
Trofica perché portano nel proprio contesto i vasi, dai vasi attraverso la diffusione mediata dalla sostanza
fondamentale anista, e' portato avanti il trofismo del connettivo e di tutti i tessuti privi di vasi sanguigni.
Cicatriziale: là dove il tessuto venga leso e le membrana basale interessate nella lesione, il tessuto connettivo si
mette ha fare sostanza intercellulare che chiude e ripristina la soluzione di continuità e quindi ridà integrità al
tessuto formando quello che si chiama cicatrice. Se il tessuto e' soggetto a rinnovo si ripristinerà nella sua forma,
altrimenti resta una cicatrice connettivale che la funzione non la ripristina più, quindi chiude soltanto.
Difesa: al livello dei tessuti connettivi avvengono tutti i fenomeni della difesa sia quella mediata dagli anticorpi
(difesa umorale(?)) sia quella mediata dalle cellule.
Da ora in poi ci occupiamo dei tessuti propriamente detti.
Questi tessuti connettivi possono avere fibre di tutti i tipi che abbiamo visto e la sostanza fondamentale anista.
Quali cellule hanno questi tessuti connettivi? cellule che possono essere divise in 2 categorie:
►derivati da precursori locali
Quali sono le cellule derivate da precursori locali?I fibroblasti e fibrociti,
miofibroblasti e gli adipociti. La cellula staminale che rimane come compartimento
germinativo e' la cellula mesenchimale che resta anche nell'adulto attorno ai vasi
sanguigni.
►derivati da precursori ematogeni.
Le cellule derivate da precursori ematogeni sono i macrofagi, i mastociti, le
plasmacellule. Inoltre in questi connettivi troviamo anche leucociti riconoscibili come tali
cioè cellule provenienti dai vasi sanguigni che mantengono la struttura che avevano in
circolo e non subiscono modificazioni,differenziamenti a livello del connettivo.
FIBROBLASTI-FIBROCITI questo termine indica lo stesso tipo cellulare derivante dal
mesenchima in momenti funzionali diversi. Il fibroblasto produce attivamente tutte le componenti
della sostanza intercellulare, produce le fibre, produce le componenti (tropoglicani, gag e le
glicoproteine della sostanza fondamentale anista) e ne regola il loro turn-over anche se è molto
lungo nel tempo perché sono elementi molto stabili, tutt’al più vengono rimaneggiati. Il fibroblasto
ricorda molto la cellula mesenchimale formando un citoreticolo anch’essa, ma nelle maglie c'e' una
sostanza intercellulare da esse fabbricate e regolate. E’ ricco di organuli proprio perché c’è un intesa sintesi di
proteine e glicoproteine come componenti della matrice. Quando l'attività del fibroblasto termina si riduce a
fibrocita cioè una cellula che riduce molto il suo corredo di organuli e si limita al lento turn-over delle
componenti già fabbricate e come già detto con una vita molto lunga.
Quando i tessuti connettivi propriamente detti sono ricchi di fibroblasti?durante lo sviluppo (perché va
fabbricato),durante l'accrescimento (perché il tessuto va cresciuto),poi nell’ adulto la maggior parte sono fibrociti
pronti a ridiventare fibroblasti, cioè cellule attive, qualora il tessuto venga leso. Per esempio, durante il trauma o
una malattia, il sistema fa le cicatrici connettivali o a sostituire altri tessuti, ecco che ci sono richiami a
ridifferenziarsi in fibroblasti, e quando questo non basta dal mesenchima, dalle cellule mesenchimali vicino ai
vasi capillari, si differenziano di nuovi. Ecco qui, da parte dei fibroblasti la produzione di tutte le componenti
della sostanza intercellulare. Durante la guarigione di una ferita c'e' un momento in cui dal fibroblasti si
differenziano in, così detti, miofibroblasti cioè cellule con requisiti del fibroblasto, ma pongono un apparato
contrattile sviluppato per cui queste cellule funzionano un po' come cellule muscolari lisce.
ADIPOCITI: sono il terzo tipo di cellule differenziate dalle cellule mesenchimali.
si possono ritrovare in 2 morfologie diverse a seconda che l'immagazzinamento dei
grassi, trigliceridi, sia tutto in un accumulo o in più accumuli. Poiché questi
accumuli di grasso l'istologo spesso li solubilizza, si vede l'immagine negativa, da
cui il nome adipocita multivaculato o multivaculare e adipocita monovaculato o
mononucleare. Perché questi nomi che indicano spazio vuoti? perché gli accumuli
di grasso nell'immagine al microscopio sono spesso solubilizzate quindi appaiano
immagini bianche,vuote, da qui i nomi.
Cellule derivate da precursori ematogeni, significa che la cellula mesenchimale ha formato il midollo, il midollo
ha dato origine agli elementi del sangue, questi attraversano l'endotelio, vanno nel connettivo e si trasformano in
macrofagi, mastociti o in plasmacellule.
MACROFAGI:cosa si intende per macrofago? una cellula del connettivo che origina
dai monociti del sangue. Quindi ci dobbiamo immaginare che in circolo, nel sangue, tra i
GB, ci siano i monoliti, quelli che si innalzano quando uno ha la mononucleosi. Questi
monociti attraversano la parete endoteliale vanno nel connettivo e si differenziano in
macrofagi, cellule della difesa, ed esplicano la funzione della difesa rilasciando
citochine cioè sostanze che richiamano altre cellule della difesa,fattori di crescita per la
sostanza intercellulare e azione antibatterica, ma soprattutto attuano la fagocitosi e la
digestione con gli enzimi lisosomiali. Il macrofago può essere fisso, ancorato alle fibre del connettivo, ma può
essere richiamato, come Pollicino che lasciava le bricioline, da dei mediatori chimici nelle zone dove si deve
esplicare la funzione di difesa attraverso la fagocitosi. Ecco quindi che si stacca e segue, con movimento
ameboide, questo percorso (si chiama CHEMIOTASSI) e arriva laddove esplicherà la propria funzione. I
macrofagi sono cellule molto eterogenee perché, a seconda dell'ambiente in cui il macrofago e' localizzato,
faranno la digestione di sostanze diverse. Il macrofago è una cellula con tante protuberanze (lamellopodi(?)) che
gli servono per aderire a substrati molto diversi e ricca di lisosomi che attaccheranno le sostanze fagocitate e
cercheranno di distruggerle. Nel caso di corpi lipidici ci può essere una problematica, quella di rimanere
all’interno come corpi lipofuscinici. Altro esempio: nell’alveolo polmonare: il macrofago scorre, mutando forma
attraverso il movimento, nello strato disulfactante, tenendo a controllo e ripulendo la superficie alveolare. Porta
avanti anche la propria funzione più su nelle vie aeree, di conduzione, nel connettivo sottostante e inglobando il
materiale di scarto. Esempio: nella milza il macrofago digerisce globuli rossi vecchi ma la digestione porta il
corpo residuo con il ferro perché i lisosomi non lo digeriscono, e si accumula nel citoplasma. In questo caso ci
sarà un meccanismo di recupero di questa sostanza.
Detto così abbiamo visto il macrofago come cellula adibita alla difesa, Di fatto i
macrofagi sono più eterogenei e di fatti si parla di SISTEMA DEI MACROFAGI che
intende una superfamiglia di cellule accomunate all'origine dal monocita del sangue (→
)
e dalla difesa attraverso il rilascio di queste sostanze,la fagocitosi,la digestione
lisosomiale interna o esterna o la capacità di presentare l'antigene. Ecco quindi che
questo sistema si arricchisce oltre dei macrofagi di tutti i tessuti di tutti gli organi, portati
avanti fagocitosi verso sostanze diverse, delle cellule presentanti l'antigene di cui
abbiamo visto l’esempio delle cellule di Langerhans dell'epidermide, ma anche di altre
mucose come quelle boccale, degli osteoblasti cellule del tessuto osseo,della microglia
essere cellule del tessuto nervoso. Tutte cellule accomunate da quella funzione e da quell'origine anche se si
diversificano nella modalità con cui esplicano la funzione difensiva.
MASTOCITI:derivano dalla popolazione di cellule monocitarie (un po’ diverse di quelle che
danno origine ai macrofagi) e che sono alla base di processi di difesa soprattutto nelle
sindrome allergiche, sono infatti le cellule che rilasciano granuli (MEDIATORI) contenenti
eparina ed istamina che sono sostanze anticoagulanti, rispettivamente, e vasodilatatrici che
portano il nostro organismo, insieme a molecole di tipo citochine generate anche esse durante
la risposta,a riconoscere come estranee determinate sostanze, gli allergeni, che possono essere
diversi da un individuo all'altro,uno può essere allergico a niente, chi al polline, alle graminacee, profumi perché
l'organismo di queste persone riconosce queste sostanze come estranee, avverte il sistema difensivo che una
sostanza estranea e' entrata nell'organismo, i mastociti liberano istamina ed eparina i quali danno origine a una
serie di eventi come l'iperproduzione di muco per esempio,o l'edema per passaggio di acqua nei tessuti, fino alla
risposta più acuta che e' lo shock, che se non si attivano rapidamente delle metodiche drastiche l'individuo muore.
Le reazione allergiche si blocca prendendo un antistaminico.
PLASMACELLULE:derivano per differenziamento dalle cellule del sangue che
sono i linfociti B,i linfociti B riconoscono antigeni specifici, si dividono tante volte,
vanno nel connettivo dove si differenziano in cellule effettrici (le plasmacellule) che
su preciso disegno di recettori di membrana che hanno riconosciuto l'antigene,
fabbricano anticorpi specifici contro quell'antigene. Sono quelle cellule dotate di
tanto reticolo e di grosso apparato di Golgi perché devono fabbricare tante
glicoproteine che sono gli anticorpi, immunoglobuline gamma, le riversano,
l'anticorpo trova l'antigene,lo riconosce,ci si lega e lo disattiva, altre cellule
toglieranno di torno il complesso antigene-anticorpo. Nel tessuto connettivo propriamente detto ritroviamo anche i
leucociti riconoscibili come tali e per tanto trattati nel sangue.
Quindi il tessuto connettivo propriamente detto è quel gruppo di tessuti connettivi, cellule e sostanza
intercellulare, costituiti da sostanza fondamentale anista così come l’abbiamo descritta, più o meno abbondante,
fibre più o meno abbondanti di un tipo o di un altro e cellule, cellule che possono essere derivate da precursori
locali, la cellula mesenchimale (fibroblasti e fibrociti ubiquitari perché fatti da sostanza intercellulare, qualche
volta miofibroblasti), adipociti se sarà in un tessuto che li prevede. Poi cellule derivate da precursori ematogeni,
macrofagi in tutte le loro forme, mastociti e plasmacellule, inoltre, in alcuni momenti ci potranno essere anche
localizzati globuli bianchi così come sono (monociti, linfociti, e granulociti)
TESSUTO CONNETTIVO PROPRIAMENTE DETTO: LA SISTEMATICA
Il tessuto connettivo propriamente detto si divide in forme:
♦ lasse
♦ dense
sulla base, fondamentalmente, della quantità e delle aggregazione delle fibre.
Forme lasse caratterizzate da abbondanza di sostanza fondamentale anista, abbondanza di cellule. Le fibre se ci
sono non sono mai organizzate in grosse fasci.
Possono essere di tutti i 3 tipi: collage, reticolare ed elastiche.
Forme dense caratterizzate dall'abbondanza della componente fibrosa sempre avvolta o in grosse fasci,le fibre
collagene,o in lamine,le fibre elastiche. Le fibre reticolari non danno origine a forme dense.
Nelle forme dense prevalgono le funzione meccaniche, nelle forme lasse prevalgono la funzione trofica,di
difesa e cicatriziale.
Forme lasse
tessuto mucoso. E' un delicato reticolo di cellule di natura fibrocitaria nelle
cui maglie c'e' lassamente localizzato fibre collagene e sostanza
fondamentale anista. e' un esempio di tessuto connettivo privo di vasi
poiché e' fatta in ballo ai vasi del funicolo ombelicale. E' un tessuto
estremamente turgido, elasticamente resiste alla compressione.
Cosa e' il funicolo? e' il cordone ombelicale, e' quella struttura che lega,porta i vasi dalla placenta che media gli
scambi tra madre e feto a quello che sarà la futura zona ombelicale dell'individuo,dentro ci stanno i vasi che
portano il sangue che si e' ossigenato dalla placenta e arricchito e riportano indietro quelli che sono gli scarti. Si
pensa che il feto si rigira perché sta dentro un sacco, il sacco amniotico, in cui c'e' del secreto, il liquido
amniotico. Questo cordone, funicolo, se il feto lo schiaccia, perché magari si viene a trovare a contrasto con le
ossa del bacino materne, schiaccerebbe i vasi con grande danno per lo sviluppo. Invece è un tessuto che sguscia
che non dà origine a pieghe secche, proprio per la sua struttura ricca di fibre collagene sottili e di sostanza
fondamentale anista particolarmente ricca di proteoglicani.
Il tessuto fibrillare lasso forma le tonache connettivali, le lamine proprie quindi, di tutte le tonache mucose.
quando si parla di mucosa, si parla di una tonaca mucosa alla cui superficie c'e l'epitelio bagnato dal muco, sotto
c'e un connettivo che e' il fibrillare lasso, poi forma lo stroma di tutti gli organi, portando vasi, nervi e quant’altro
e il derma papillare,il derma superficiale. e' un derma stromale riccamente vascolarizzato, ricco di fibrille di tutti
tipi, diventa più ricco di fibre reticolari verso la membrana basale, ed e' l'unico tessuto connettivale in cui si
possono trovare tutte le cellule descritte finora. E’ cioè caratterizzato da polimorfismo cellulare,vuol dire che nel
proprio contesto può avere grande varietà di cellule,cellule polimorfe,diverse.
Tessuto reticolare e' un tessuto del connettivo propriamente detto costituito in prevalenza da fibre reticolari,
possono essere su un piano per cui reticolo bidimensionale o reticolare tridimensionale, con giunzioni. Il tessuto
reticolare e' costituito da fibre reticolari, naturalmente da fibroblasti e fibrociti che sono le cellule che le fibre
vanno a costituire.
La sostanza fondamentale anista e' abbondante nel reticolare bidimensionale (quello in cui le fibre reticolare si
dispongono su un piano a formare il reticolo come fosse un foglio). Questo tessuto reticolare forma la lamina
reticolare della membrana basale, quella struttura che sappiamo essere costituita da 2 porzioni:
■la lamina basale che viene prodotta dagli elementi non connettivali, (l'epitelio, l'endotelio,
gli elementi del tessuto muscolare ed alcuni elementi del tessuto nervoso);
■la lamina reticolare e' connettivo ed e' fatta da fibroblasti e fibrociti.
Come si armonizzano queste due strutture?per esempio nell'epitelio di rivestimento, in cui le cellule dello strato
basale elaborano la porzione, la lamina basale, che lega le cellule epiteliali al connettivo sottostante in particolare
alla lamina reticolare costituita da un intreccio,una specie di impalcatura, una rete, che ci sta sotto e le cui maglie
sono chiuse dalla sostanza fondamentale anista. La lamina basale che e' secreta da un tessuto epiteliale,
dall'endotelio, dagli elementi del muscolo e dal tessuto nervoso, si vede solo al microscopio elettronico di solito
,ed e' costituita da 2 porzioni:
▪ una lamina rara costituita da glicoproteine di legame quale la laminina, è una
glicoproteina che si attacca alla membrana delle cellule epiteliali e poi si attacca al
connettivo sottostante legando l'epitelio in maniera molto stretta.
▪ La lamina densa che e' fatta da un particolare tipo di collagene, di tipo IV che forma un
reticolato, questa lamina basale si lega attraverso altre glicoproteine di legame alla
lamina reticolare, il tutto ancora gli epiteli e altri tipi di strutture al tessuto sottostante che
e' il connettivo che portando i vasi, apporta trofismo, difesa e potenzialità cicatriziale.
In alcuni settori quali la parete degli alveoli polmonari, per fare un esempio, al di sotto degli pneumociti di tipo I
e II e al di sotto dell'endotelio, la membrana basale e' priva di lamina reticolare, sono cioè due lamine basali fuse
per rendere questa barriera meno spessa tra aria e sangue.
Le caratteristiche funzionali della membrana basale sono: meccaniche, ancorano le porzioni non connettivali al
connettivo tramite il sinergismo di glicoproteine e di fibre reticolari,ma controllano anche la permeabilità, le
membrane basali operano un filtro tra ciò che passa e che non deve passare,tra la quantità che passa e che deve
essere diminuita. Ci sono malattie metaboliche tra cui il diabete che può portare ispessimento della membrana
basale con difficoltà di scambi tra i tessuti. Sono quindi una barriera selettiva anche contro la migrazione di
sostanze potenzialmente dannose ed hanno un ruolo nel differenziamento, nell’organizzazione dei tessuti.
Abbiamo più volte detto quando si parlava dell’epidermide, l’avvio al differenziamento che porta a squame
cornee per perdita di contatto con la membrana basale.
Il tessuto reticolare tridimensionale va' a fare l'impalcatura, impalcatura degli organi linfoidi ed ematopoietici cioè
il midollo osseo,oltre ad essere fatta di un reticolo tridimensionale di fibre reticolari, porta nel proprio contesto
cellule che fabbricano le fibre (fibroblasti e fibrociti) e cellule della difesa (linfociti, plasmacellule, macrofagi) e
cellule dendritiche cioè cellule presentanti l'antigene perché i linfonodi sono stazione di difesa dell'organismo.
Tessuto adiposo → Si presenta in 2 varietà di cui una, il tessuto bianco, fatto in
prevalenza da adipociti monovaculato, e' un tessuto che accumula trigliceridi e forma
un tessuto riccamente vascolarizzato che ha molteplici funzioni:
-La prima e' una riserva nutritizia, se noi digiuniamo attingiamo dal nostro tessuto
adiposo
-Poi isola termicamente soprattutto l'adipe sottocutaneo
-Poi imbottisce, specialmente nel palmo della mano e nella pianta del piede dov'e' meno metabolizzabile.
-Ha anche funzione fisiognomonica cioè è diverso come localizzazione da individuo a individuo e da uomo a
donna. E poi è diverso a seconda della dieta, della genetica e a seconda del tipo di vita che si fa.
Fa anche da imbottitura a determinati organi come intorno ai reni e nella zona del bulbo oculare, ma mentre
l’adipe sottocutaneo è facilmente mobilizzabile tramite la dieta, quello dei reni e dell’occhio e delle guance è
meno mobilizzabile.
Il tessuto adiposo bianco ha funzione endocrina, secerne ormoni che riversa nel circolo sanguigno e che sono alla
base regolativi, insieme ad altri ormoni, dell'assunzione di cibo,del metabolismo glicidico e producono dei
metaboliti attivi per la sintesi degli ormoni steroidei.
Il tessuto adiposo bruno e' costituito da adipociti plurivaculati, è riccamente vascolarizzato,
e' abbondante nel feto, nel neonato si esaurisce e nell'adulto è pochissimo e confuso con il
tessuto adiposo bianco; perché questo? perché il tessuto adiposo bruno e' un tessuto che,
invece che attraverso il mitocondrio produrre ATP, utilizza quell'energia per produrre
calore che riscalda il sangue che passa nelle immediate vicinanze dei capillari e quindi ha
questa regolazione sulla temperatura corporea. E' particolarmente importante per gli animali ibernanti perché nel
momento del risveglio utilizzano il grasso bruno per riportare la temperatura corporea a livelli ottimali. Nell'uomo
ha un ruolo essenzialmente al momento della nascita: il feto viene sbalzato da una temperatura interna del corpo
materno di oltre 37° a temperatura ambientale che sarà 21° o 22°, ecco che utilizza il suo grasso bruno per
riscaldare il sangue e superare questo shock termico (viene prontamente metabolizzato).
Le forme dense
Sono caratterizzate da grosse fasci di fibre collagene,che hanno un andamento,queste
grosse fasci, diverso secondo linee dello spazio diverse, a seconda degli stimoli che
vengono portati in superficie al tessuto che fa parte di un certo organo. Il tessuto
elastico e' fatto da lamine o da gruppi di fibre elastiche.
E’ un tessuto a fasci intrecciati disposti su tutte le dimensioni dello spazio,
grossi fasci di fibre collagene che portano fibroblasti che tengono il controllo e vasi
sanguigni nel contesto e sostanza fondamentale anista; queste fibre che decorrono in tutte le direzioni dello spazio
ammortizzano tutte le forze provenienti dalla superficie secondo varie direzioni, tant’è vero che questo tessuto va
a costituire il derma profondo, che contiene nel proprio contesto, oltre grosse fasci di fibre collagene, anche delle
fibre elastiche. Si hanno le smagliature quando si supera il potere elastico delle fibre elastiche e si sono rotte,
dando origine prima a delle strie rosse poi bianche. Ma anche per chi prende troppo sole, le fibre elastiche
vengono lese dai raggi ultravioletti (quando siamo giovani vengono riparati dai fibroblasti).
Il tendine e' un tessuto connettivo denso i cui i fasci sono paralleli, infatti le forze sono
applicate ai capi,da un capo si ancorano all'osso da quell'altro al muscolo,il muscolo si
contrae, il tendine deve essere fatto da fibre che impediscano la distensione sennò la
contrazione non avrebbe nessun effetto,resistenti e disposte a fasci paralleli in cui il
connettivo fibrillare lasso li va a fare degli involucri.
Il tessuto elastico e' una serie di fibre elastiche che formano lamine che vanno a costituire anche parte della parete
dei vasi. Esempio: la parete delle arterie.
IL PROCESSO DI CICATRIZZAZIONE
Quando il tessuto connettivo viene leso, intanto si ha perdita ematica che viene tamponata per mezzo di un
coagulo che forma in superficie una crosta e c’è richiamo di cellule della difesa. Il tessuto connettivo forma vasi
nuovi e poi richiama, per migrazione e proliferazione, fibroblati (si ha così tessuto di granulazione) che
cominciano a deporre il collagene che chiude e ripristina l’integrità formando un tessuto più chiaro.
Successivamente questo rammendo viene ristretto, reso più sottile, perché si sviluppano i fibroblasti che
contraendosi chiudono la ferita. Se il tessuto sottostante era soggetto a rinnovo si reintegra per ragioni sue.
Le parole che ho scritto in arancione sono quelle che non ho ben capito.
7° LEZIONE ISTOLOGIA
TESSUTO CONNETTIVO
Il tessuto connettivo è formato da cellule e sostanza intercellulare. Quest’ultima fatta da sostanza fondamentale
anista con caratteristiche di funzione trofiche, di difesa e selezione. Le costituenti caratteristiche della sostanza
fondamentale anista oltre ad una grossa percentuale d’acqua sono le glicoproteine, proteoglicane e
glicosamminoglicane che sono alla base delle funzioni che dicevamo prima.
Le fibre di questi tessuti che sono di tre tipi: fibre collagene, reticolari ed elastiche, devono essere conosciute nelle
loro caratteristiche fisiche e nella loro costituzione meccanica.
Le cellule che sono tutte di origine mesenchimale possono essere divise in cellule che derivano da precursori
locali: la cellula Mesenchimale nel contesto di uno specifico tessuto connettivo oppure provenire da un precursore
ematogeno che ritrova lontanamente il precursore mesenchimale ma che colonizzando nelle zone omopoietiche
forma li una cellula progenitrice che fa da cellula staminale. Nei tessuti connettivi ci sono poi cellule del sangue: i
leucociti, riconoscibili come tali. Abbiamo preso in considerazione vari tipi cellulari a livello dei tessuti connettivi
propriamente detti. Dopo di che abbiamo fatto la classificazione in forme lasse e forme dense sulla base del
contenuto soprattutto lipide che nelle forme lasse non si aggregano mai in grosse fasce o lamine.
Specificatamente abbiamo fatto tra le forma lasse del tessuto mucoso il tessuto fibrillare lasso e il tessuto
reticolare bidimensionale come porzione della membrana basale di cui dovete conoscere la struttura in generale,
il reticolare bidimensionale come impalcatura degli organi linfoidi o ematopoietici.
Abbiamo poi visto il tessuto adiposo in particolare nella forma cosiddetta bianca e un accenno sul tessuto adiposo
bruno. Nelle forme dense abbiamo considerato i fasci intrecciati, i fasci paralleli e membrane elastiche. Questo è
lo schema dei tessuti propriamente detti.
Stamani prenderemo in considerazione le forme specializzate nel sostegno intendendosi per tali il tessuto
cartilagineo e il t. osseo.
Il T. CARTILAGINEO è un t. connettivo quindi costituito da cellule e sost. intercellulare ma un po’
su generis in quanto è privo di vasi sanguinei e di terminazioni nervose. Poiché il t. cartilagineo è
privo di vasi sanguigni deve il suo trofismo alla possibilità di avere nelle vicinanze un tessuto
connettivo propriamente detto che essendo ricco di vasi assicura per diffusione attraverso la sostanza
fondamentale anista, dei gas, l‘ossigeno e i metaboliti verso la cartilagine, di prodotti di scarto invece in direzione
opposta e quindi porta avanti questa funzione trofica nei confronti di questo particolare tipo di tessuto connettivo.
Il pericondrio, questa lamina di tessuto connettivo propriamente detto che avvolge la cartilagine, manca soltanto
sulle superfici articolari e in altre zone specializzate.
Come vi ho detto il tessuto cartilagineo non è vascolarizzato ne innervato si nutre per diffusione per cui deve la
sua sussistenza a una lamina di tessuto connettivo denso a fasci intrecciati che si chiama pericondrio, il quale è
sempre nelle cartilagini articolari mentre si fa molto vascolarizzato nella parte interna.
Questa è la cartilagine con le sue cellule, la sostanza intercellulare e niente vasi. Nel contesto della porzione più
interna del pericondrio ci sono molti capillari, dai capillari per diffusione le sostanze passano la cartilagine e
vengono riportati indietro i prodotti di scarto sia dal punto di vista gassoso che nutrizionale.
Sulle cartilagini articolari questa mancano di pericondrio, quindi in queste zone si è selezionato un meccanismo
trofico diverso. Esiste cioè una membrana detta m. sinoviale particolarmente turcica, ricca di proteoglicani e
glicosaminoglicani e il liquido sinoviale funge da liquido di scambio per le cartilagini che qui si trovano. E’ anche
capibile come questo meccanismo si sia affermato in quanto che una membrana ricca di vasi a questi livelli
sottoposta a forze meccaniche notevoli potrebbe dare fenomeni ischemici e quindi andare a danneggiare le
strutture stesse. Essendo un T. connettivo la cartilagine è fatta da cellule da sostanza fondamentale anista e da
fibre.
Le cellule sono di due tipi: condroblasti e condrociti. I condroblasti per assonanza ricordano i fibroblasti cioè le
cellule della cartilagine che sono attivamente ingaggiate nella formazione di tutte le componenti della sostanza
intercellulare. Quindi queste C. fabbricano la cartilagine, la ingrandiscono e rimangono al disotto del pericondrio
come compartimento germinativo. Le porzioni più interne della cartilagine sono popolate da condrociti, la forma
matura del condroblasto che oltre ad assicurare un lento tourn-over della sostanza intercellulare che ha emipite
molto lunghe, ha però due caratteristiche proprie. Da un lato è una cellula che tende ad accumulare prodotti sotto
forma di glicogeno e lipidi, dall’altra parte c’è da capire che è una cellula lontana man mano nelle porzioni più
interne della cartilagine dalla fonte di sostentamento che è il pericondrio. Quindi sono cellule che tendono ad
accumulare prodotti di riserva.
Domanda: anche il condrocita è sotto il pericondrio? No. La lamina avvolge tutto poi c’è un t che è la cartilagine,
le porzioni periferiche condroblasti che sono attivi mentre la cartilagine si forma e si ingrandisce poi
compartimento germinativo sono pochi elementi, il resto sono condrociti i quali possono trovarsi molto
internamente nella cartilagine accresciuta e quindi lontana dal pericondrio da cui si devo nutrire per diffusione,
tendono per tanto ad accumulare prodotti di riserva. Questa c al minimo nelle prime fasi dello sviluppo e
dell’accrescimento tende a mantenere una caratteristica di c giovane cioè si divide topicamente dando origine a c
che rappresentano il cosiddetto gruppo isogeno. Cioè un gruppo di c tutte riunite nella stessa zona che
riconoscono la stessa c progenitrice. Quindi un condrocita da origine a un progene che è un clone cellulare
chiamato gruppo esogeno, ogni condrocita può dare origine a questi gruppi. Osservando un’immagine vediamo
quindi: una cartilagine, il pericondrio, qualche condroblasto, alla periferia condrociti isolati o raccolti in gruppi
esogeni. Si notano alcune c ricche di glicogeno come uno dei prodotti di riserva di cui parlavamo prima.
La sostanza fondamentale anista ha le stesse caratteristiche delle sostanze fondamentali aniste dei connettivi
propriamente detti ma è molto ricca di proteoglicani che tendono, ciascuno di questi è schematizzato in un
proteoglicano, ad associarsi come setole di uno spazzolino su assi che sono costituiti da acido ialuronico. Quindi
gli edifici molecolari già grandi dei proteoglicani assemblandosi sopra l’asse dato da acido ialuronico tendono a
diventare ancora più grandi legando grandi quantità di acqua e lasciandola in equilibrio con quella non legate
quindi mediando questo modo di diffusine così importante. Non solo ma determinandone anche la caratteristica
fisica della incomprendibilità. Le cartilagini sono strutture di sostegno che devono in parte le loro proprietà
meccaniche a questa costituzione.
Le fibre possono essere di tre tipi :
-sottili fatte da un tropocollagene, in particolare quelle di secondo tipo. -elastiche
-collagene di primo tipo
Sono alla base della costituzione di cartilagini diverse con proprietà meccaniche diversificate. Abbiamo già posto
le basi per fare una classificazione. La cartilagine ialina, elastica e fibrosa sono le tre categorie in cui viene divisa
la cartilagine, il t cartilagineo.
Partiamo dal dare brevi definizioni, dare qualche caratteristica dei tre tipi di cartilagine con le rispettive
localizzazioni:
Cartilagine ialina deve questo nome all’aspetto translucido trasparente vetroso che
questa cartilagine ha a fresco. E’ bianca ed è caratterizzata da avere gruppi esogeni costituiti da
molte cellule, presenta sottili fibrille di collagene di secondo tipo che essendo molto sottili
mantengono la trasparenza di questa cartilagine. Costituisce la maggior parte dello scheletro fetale che vedremo
poi verrà sostituito da tessuto osseo. Nell’adulto la ritroviamo sulle superfici articolari dove è priva di pericondrio,
cartilagini nasali, tracheali e bronchiali non dove formano anelli che tengono pervie queste vie e la parte
cartilaginea delle coste che si articolano sulla porzione sternale e non.
Cartilagine elastica è una cartilagine che a fresco è giallastra, ha condrociti
disposti, gruppi isogeni ma di poche unità e presenta oltre a fibre collagene tipiche,
una componente di fibre elastiche che ne da una caratteristica fisica alla cartilagine.
Due esempi di cartilagine elastica in cui sono messi in evidenza la grande quantità di
fibre elastiche. Le porzioni in bianco sono dove esistono le cellule non colorate dove le fibre elastiche sono
colorate in blu violetto. La cartilagine E. ha oltre la possibilità di essere un tessuto di sostegno anche quella di
potersi deformare e riformare se non si rompe. Allo stato primitivo si può fare l’esperienza toccandosi il
padiglione auricolare costituito da cartilagine elastica. Esiste questo tipo di cartilagine anche nel condotto uditivo
esterno, in parte nelle cartilagini della laringe e bronchiolare, le sezioni più sottili dell’albero bronchiale.
Cartilagine fibrosa chiamata così dagli istologi, fibrocartilagine dagli anatomici. E’
una C. che ha caratteristiche che vedremo molto vicino al tessuto connettivo propriamente
detto fibroso in quanto è estremamente ricca di fasci di fibre collagene che si dispongono
tra le file di cellule condrociti che spesso hanno questo andamento rispondendo in tal modo
alle pressioni meccaniche di superficie. Sembra che sia assente un tipico pericondrio e che ne facciano le veci i
tessuti connettivi propriamente detti che sono localizzati nelle vicinanze. La C. fibrosa è dislocata come la
porzione periferica nel disco intervertebrale, partecipa alla sinfisi cubica e costituisce i menischi articolari.
Facciamo nei dettagli l’esempio della C. fibrosa nel disco intervertebrale: tra i corpi vertebrali esiste diciamo così
un cuscinetto ammortizzatore che è costituito nella porzione interna di ciò che rimane del tessuto embrionale che
nell’adulto forma il cosiddetto nucleo polposo. La porzione periferica in tale disco intervertebrale è di C. Fibrosa
dove le cellule rispondono alle pressioni tra le vertebre disponendosi in file, risultato anche di proliferazione dei
condrocito, quindi non si vede il tipico assemblaggio del gruppo esogeno ma niente ci dice, nega che ci possa
essere questo fenomeno ma le cellule si dispongono in file. Tra le fibre, grossi fasci di fibre collagene che ne
conferiscono una grossa resistenza meccanica. Sapete tutti che qualunque perdita di contenzione di questo lamello
nei confronti del nucleo polposo da origine alle ernie: le ernie discali molto dolorose e invalidanti. Quindi un
breve accenno che mostra, vuoi come si forma la cartilagine, vuoi come si accresce fino alle dimensioni
dell’adulto. In seno ad un mesenchima le cellule mesenchimali all’inizio cominciano a differenziarsi dando
origine ai condroblasti che secernono sostanza intercellulari della C. e da un nucleo centrale man mano fino alla
periferia questo abbozzo si espande. Man mano che si espande, i condroblasti sono condrociti, i quali continuano
a dividersi dando origine ai gruppi esogeni e quindi espandendo la cartilagine anche dall’interno. Intanto
esternamente dal mesenchima si differenzia laddove esiste un tessuto connettivo ad opera dei fibroblasti
propriamente detto che formerà il pericondrio. Così la C si accresce per apposizione dall’esterno ad opera dei
condroblasti, per divisione dall’interno dei condrociti e quindi interstizialmente. Se ci fosse una domanda come si
accresce la cartilagine? Con queste due modalità. Tanto è vero che se si rompe sembra che sia più facile che si
ripristini l’integrità, anche se con tempi molto più lunghi, se è coinvolta la porzione più esterna che coinvolge il
compartimento germinativo. La C. poi esterna è di numerose notificazioni e quindi delle condizioni non buone
durante il periodo di senescenza e quindi dell’invecchiamento.
TESSUTO OSSEO
Passiamo ora al t osseo che c’ interessa più particolarmente e cerchiamo
di tratteggiarne le linee insieme. Anch’esso è un t. connettivo
specializzato per il sostegno. È costituito da cellule e sostanza
intercellulare come tutti i connettivi ma la sostanza intercellulare si
presenta mineralizzata. Costituisce l’impalcatura del corpo e protegge
alcuni organi come ad es. l’encefalo da parte della teca cranica. Inoltre,
oltre a questa funzione meccanica diversificata, vuoi impalcatura, vuoi
protezione partecipa alla regolazione della concentrazione ematica del
calcio. Cioè quella che prende il nome di omeostasi calcica. Dobbiamo
considerare lo scheletro come una banca del calcio in grado di
mobilizzarlo molto velocemente, immetterlo in circolo quando la
quantità di calcio ematica si abbassa. D’altro canto viene invece segregata dentro le ossa quando questa s’innalza.
Non è l’unico meccanismo regolativi in quanto che, quando s’innalza viene anche inibita la arterica intestinale,
cioè l’infezione di calcio intestinale e l’escrezione renale viene aumentata. Viceversa quando il calcio ematico
abbassa, si aumenta quello che viene assorbito e si diminuisce l’escrezione renale. Quindi, cosa voglio intendere
con questo discorso? Cosa si deve trattenere per l’istologia? Che esiste un equilibrio ematico della concentrazione
del calcio. Questa concentrazione che subisce variazioni viene regolata anche attraverso la mobilizazione del
calcio delle ossa.
I t. connettivi, come il t osseo, continuano ad avere le caratteristiche dei t connettivi, quindi ad avere cellule e
sostanza intercellulare sottoforma di fibre che nel t osseo sono di tipo collagene di primo tipo e sost fondamentale
anista ricca di acido ialuronico, proteoglicani e glicoproteine che oltre ad avere le funzioni che abbiamo più volte
indicato sono anche guida nella deposizione dei sali di calcio. Sali di cacio che vanno a costituire la componente
minerale sottoforma di cristalli che hanno questo nome idrossiapatite sono come degli aghi cristalli che tendono a
riunirsi insieme e a formare cosi la componente minerale del tessuto osseo. Abbiamo 4 fenotipi cellulari
riconducibili a due linee diverse:
-una è quella che prende avvio per differenziamento in sito cioè nel tessuto dalla cellula mesenchimale e
riconosciamo le cellule osteoprogenitrici che sono cellule indifferenziate ma già con un itter cioè commissionate,
indirizzate a diventare c. del t. osseo. Loro non hanno più, sebbene differenziate, la possibilità di differenziarsi in
tante altre cose, hanno ristretto il campo. Gli osteoclasti esprimono le caratteristiche di c del t. osseo e come dice
il nome producono attivamente le componenti della sostanza intercellulare o matrice e inducono la
mineralizzazione. Gli osteociti sono la forma crescente, matura dell’osteoblasto e provvedono alla fineregolazione
anche della mineralizzazione. Quindi, sia il livello dei t propriamente detti della cartilagine come del t osseo esiste
una popolazione c. che si chiama con doppio nome per indicarne la forma attiva e la forma crescente che si
incarica di regolare il fine quindi della sostanza intercellulare già deposta e accresciuta.
-La seconda linea cellulare è data dagli osteoclasti che come dice il nome stesso demoliscono il tessuto osseo.
Sono cellule infatti che derivano dai monociti del sangue circolante e appartengono come già preannunciato alla
famiglia o sistema dei macrofagi in quanto che rilasciano sostanze, cioè enzimi lisosomiali che sciolgono il
tessuto osseo sia la componente minerale che nella componente organica.
Guardando un’immagine:
Questo è un osso in neoformazione cioè si sta formando. Questi alla periferia sono gli osteoblasti, cellule piuttosto
voluminose che tendono a disporsi in file, hanno giunzioni comunicanti tra di loro e attraverso un apparato
proteosintetico molto sviluppato elaborano le componenti della matrice e la mineralizzano perchè fabbricano i
globuli della matrice o granuli calcificanti. Io non entro nel fenomeno della mineralizzazione, voi però siete tenuti
a sapere che gli osteoblasti oltre a fabbricare sostanza fondamentale anista e fibre collagene partecipano al
fenomeno della mineralizzazione rilasciando nella matrice dei globuli, dei granuli che provocano il depositarsi di
cristalli di idrossiapatite. Ricapitolando l’osteoblasto rilascia questa sostanza cellulare nella sua componente
organica, in più elabora e quindi esocita nella sostanza, nella matrice dei granuli, dei globuli che hanno nome
diversi a secondo se stanno dentro o fuori che provocano il depositarsi locale dei sali di calcio sotto forma di
cristalli di idrossiapatite, quindi mineralizzano tessuto osseo. Una volta che questo è avvenuto la sostanza
intercellulare mineralizzata circonda la cellula che tende a diventare la forma matura: l’osteocita.
Apriamo una parentesi. La cartilagine si nutriva per diffusione e quindi poteva contare su membrane con i vasi
sanguinei da cui, attraverso la diffusione mediata dalla sostanza fondamentale anista, ricevevano ciò di cui
avevano bisogno e verso cui mandavano i prodotti di scarto. Ma il tessuto osseo non può far conto sulla diffusione
attraverso la sua sostanza intercellulare, in quanto che calcificata, un muro. Ecco quindi che nel T.osseo
assisteremo alla presenza di tutta una serie di canali in cui si infilano i prolungamenti delle cellule, serti di
connettivi propriamente detti che assicureranno in questo modo il trofismo della struttura. Cominciando
dall’osteocita che non si trova più nella periferia dell’osso ma dentro e quindi sviluppa prolungamenti con cui
rimane in contatto con i suoi vicini tramite giunzioni comunicanti. Il corpo dell’osteocita sta in una lacuna scavata
nella sostanza intercellulare mineralizzata e i suoi prolungamenti in canalicoli. Questi prolungamenti sono in
contatto con gli osteociti vicini e i canali sono foderati da membranelle ricche di proteoglicani che mediano loro la
diffusione. Quindi, vuoi attraverso il contatto, vuoi attraverso queste membranelle che li circondano riceveranno il
trofismo. Ecco come avviene, questa è la sostanza intercellulare, questi sono gli osteociti ampiamente
comunicanti tra di loro e circondati da queste membranelle che permettono il diffondersi a partire da connettivi
vascolarizzati di queste possibilità trofiche. Ecco di nuovo preso il prolungamento di un osteocita, quello di un
altro che nei canalicoli si vengono a toccare e formano una giunzione. Tutti questi canalicoli ampiamente
comunicanti tra di loro sono scavati nella sostanza intercellulare mineralizzata. Dentro questi canalicoli ci stanno i
prolungamenti delle cellule tipiche del T. osseo adulto che sono gli osteociti essendo nei vari osteoblasti la forma
attiva nella deposizione e quindi rimangono tali nei momenti in cui producono poi diventano osteociti. L’
osteoclasto è plurinucleato, si forma per sincizio. L’osteclasto è inattivo e rotola lungo canali del tessuto osseo.
Quando gli arrivano delle informazioni attraverso citochine o ormoni, i propri recettori scatenano una risposta
cellulare che fanno si che l’osteoclasta si attacchi come una ventosa in una zona precisa dell’osso, si giri la zona
che deve riassorbire proprio come se ci attaccassimo una ventosa e amplificando in quel tratto la superficie,
facendo delle pieghe scarica qui gli enzimi. Prima demolisce la componente organica solubilizzando i cristalli di
idrossiapatite e rende così nuda la componente organica. Con gli enzimi lisosomiali attacca la componente
organica e riassorbe il tessuto osseo. Dopo di che, quando ha finito il suo lavoro si stacca e rotola di nuovo. Il
tessuto osseo non è mai un tessuto statico: si è formato, si è sviluppato con le dimensioni dell’adulto, non si
rinnova ma sarà sempre sottoposto a deposizione e riassorbimento.
L’infermiere è tenuto a conoscere questo dinamismo perché sta alla base di un corretto sviluppo anche negli
individui più anziani dell’apparato scheletrico. Quindi, gli osteoblasti depongono, poi diventano osteociti che sono
le cellule tipiche del tessuto adulto. Poi gli osteoclasti riassorbono con un meccanismo mediato da citochine e
ormoni.
TESSUTO OSSEO
Esistono due tipi di tessuto osseo: T. osseo fibroso eT. Osseo lamellato.
Questa è la risposta da dare alla domanda mi parli del tessuto osseo.
Il T. osseo fibroso è un T. osseo molto giovane di prima deposizione. Quando gli osteoblasti fabbricano T. osseo
lo fanno con i connotati del T. fibroso che poi gli osteoclasti demoliscono e al suo posto verrà deposto il T.
Lamellare più adatto a sopportare forze meccaniche di superficie. È un tessuto più maturo dal punto di vista della
sua organizzazione.
Per cui il t fibroso lo ritroveremo durante lo sviluppo embrionale fetale, durante l’accrescimento delle ossa e tutte
le volte che nell’equilibrio distruggere e ricostruire si sta costruendo il tessuto. Poi viene sostituito da t lamellare,
ne rimangono piccole tracce per essere onesti laddove i tendini s’inseriscono sulle ossa e nelle porzioni
dell’aggancio dei denti sulle arcate. Ma m’interessa sapere che è un osso di neodeposizione in cui c’è una
sostanza intercellulare mineralizzata con fibre collagene a gruppi disposte secondo linee di forza più svariate,
quindi s’intrecciano, è anche detto a fibre intrecciate in qualche libro e con una deposizione degli osteociti che
hanno un corpo balare i prolungamenti secondo varie regioni dello spazio. Questo è il t osseo fibroso di
neodeposizione. Questi sono gli osteociti che vedete sono disposte in tutte le direzioni e fasci di fibre collagene in
rosa-aranciato interposte in zone a maggior contenuto di sostanza fondamentale anista questo rosa più violetto,
quindi un t immaturo che verrà sostituito da t osseo lamellare, caratterizzato da una matrice intercellulare
organizzato in lamelle, all’interno di ogni lamella le fibre collagene sono parallele tra di loro. Allora, cosa vuol
dire questo discorso? Voi immaginatevi il t lamellare come una serie di fogli un po’ spessi che si appaiano vicini
gli uni con gli altri nel cotesto di una lamella fatta da sostanza intercellulare anista e fibre mineralizzata. Nel
contesto di un foglio di una lamina le fibre collagene sono parallele ma in quella vicina sono sfalsate. Ecco quindi
che è un t facilmente smontabile, via una lamella le altre restano integre in cui le forze meccaniche di superficie
vengono ammortizzate con un gioco dell’orientamento diverso delle fibre collagene da una lamella all’altra. È
quindi un t che si è affermato come leggero ma resistente ed estremamente dinamico. Ecco perché il t lamellare è
tipico dell’adulto, del t osseo adulto maturo. Tra una lamella e l’altra ci stanno gli osteociti che sono organizzate
secondo la lamella e mandano i loro prolungamenti nei canalicoli a invadere le lamelle vicine dove prendono
contatto con gli osteociti vicini. Il t osseo lamellare è di due tipi: semplice o osteonico.
Semplice- in ossa piccole. Per es alcune piccole ossa della faccia ci sono gruppi di lamelle a costituire il t.
lamellare semplice. Una lamella, due lamelle, tre, quattro, cinque…Sono lamelle di sostanza, ripeto, fondamentale
anista e di fibre parallele mineralizzato, in una lamella hanno un andamento nell’altra sfalsato così si può
ammortizzare le forze in superficie secondo vari angoli. Gli osteociti sono in contatto tra di loro quelli delle
lamelle più periferiche con i loro prolungamenti sono in contatto con lamine di t connettivo propriamente detto da
cui traggono con il meccanismo che abbiamo detto prima il nutrimento. Il lamellare semplice va a costituire
alcune piccole ossa della faccia o si ritroverà a formare le trabecole sottili dell’osso spugnoso come vedremo piu
avanti. Possiamo trovare come un forma più evoluta di t lamellare l’osteone. Cosa è l’osteone? È una struttura
cilindrica cava nel centro la cui parete è data da lamelle di sostanza intercellulare concentriche rispetto al canale
centrale. Quindi non c’è niente di nuovo è come se il lamellare semplice si chiudesse a formare una struttura
cilindrica: l’osteone.Questo ha un canale chiamato canale di Havers in cui si insinua un t fibrillare lasso che porta
vasi e nervi. partono dal canale di Havers più o meno trasversalmente o obliquamente dei canali laterali detti
canali di Wolm che metteranno in contatto vari osteomi tra di loro e alla fine troveranno un connettivo da cui
prendere tralci che si infilano in questo sistema di canali rendendo possibile agli osteociti dell’osteone pescare in
un connettivo propriamente detto da cui trarre il sostentamento. L’osteone quindi è un’unità trofica e morfologica
indipendente. è un cilindro la cui parete è data da lamelle concentriche rispetto al canale centrale. La lamella più
vecchia è quella esterna, la lamella più giovane è quella interna, vicino al canale di Havers. Ecco l’osteone in cui
sono state schematizzate due lamelle ma possono essere anche dei sistemi grandi di una ventina, di una
cinquantina di lamelle, man mano che invecchiano diventano più grossi. In cui nella porzione centrale ci si è
infilato un connettivo che porta i vasi con i globuli rossi e tutto ciò che è il sangue, i nervi e che mantiene nelle
sue vicinanze le cellule ossee progenitrici che sono il compartimento germinativo. Nella porzione più interna della
prima lamella possiamo trovare gli osteoblasti che sono quelle che le formano le lamelle, poi ci restano ingabbiate
e diventano osteociti e gli osteoclasti che vanno su e giù e riassorbono. Il canale di Havers può avere dei
collaterali laterali che sono i canali di Wolman. Il canale di Havers, le lamelle e i vari osteiciti con i loro
prolungamenti stanno ampiamente in contatto fra di loro, tutti rivolto verso il canale centrale da cui arriva il
sostentamento. L’ultima lamella è la lamella di fessura.
Abbiamo visto come è fatto il t osseo, appena deposto è fibroso poi diventa lamellato dove semplice dove
osteonico. È lamellare semplice nelle piccole ossa piatte della testa, nei sistemi circonferenziali interni ed esterni e
nelle trabecole del t osseo spugnoso. Esistono tre tipi di ossa (brevi, lunghe, piatte); esiste al loro interno due tipi
di aggregazione: l’osso spugnoso che sembra un trabecolato con tutte le cavità midollari e l’osso compatto invece
con una cavità midollare unica che ha visto microscopicamente una struttura compatta. L’osso spugnoso e
compatto visti da un punto di vista istologico:
Osso spugnoso: le fini trabecole sono fatte da t lamellare semplice. Laddove le trabecole dell’osso spugnoso
confluiscono tra di loro si può formare un osteone o piu osteoni. A foderare queste cavità delimitata dall’ intreccio
delle trabecole c’è una membranella chiamata endostio che oltre a portare la fonte trofica di sostentamento per gli
osteociti, tiene nel suo contesto anche le cellule osteoprogenitrici. Esiste differenza tra l’osso spugnoso e il tessuto
osseo spugnoso. Nei libri si confonde l’aggregazione macroscopica dell’osso interno con la struttura tessutale che
invece è lamellare. L’aggregazione può essere spugnoso compatta ma il tessuto è lamellare. Questa è una
trabecola tessutale lamellare semplice, questi sono gli osteociti che si incontrano delimitando degli spazi che sono
le cavità midollari. Se ci spostiamo nell’osso compatto prendiamo come esempio il più complesso che è la sezione
di una dialisi. Dal punto di vista microscopico: a circondare il segmento osseo c’è una membrana che prende il
nome di periostio, membrana attorno all’osso. Il periostio è un t connettivo propriamente detti a fasci intrecciati
che quindi protegge l’osso dall’esterno ma nella parte più a contatto con l’osso diventa più vascolarizzato. Poi
troviamo una serie di lamelle parallele tra di loro che è il sistema circonferenziale esterno. A circoscrivere il
canale midollare esiste una membrana che è l’endostio che provvede anche lui al trofismo del t osseo e porta
anch’esso cellule osteoprogenitrici. Nel canale c’è il midollo osseo dove giallo dove rosso. A circoscrivere
all’interno dell’endostio il canale midollare il sistema circonferenziale interno: una serie di lamelle semplici. Poi
abbiamo tutta una serie di osteoni variamente orientati più o meno con assi del canale di Havers parallelo,
leggermente obliquo al canale midollare, tenuti in contatto tra di loro dai canali di Havers che iniziano il loro
percorso a partire dalla porzione periferica o centrale dove quindi si può insinuare il t connettivo portando i vasi e
innervazioni. Se vi rompete un osso sanguinate e avete dolore. Questi osteoni sono accementati tra di loro da
gruppi di lamelle, la breccia ossea o sistema interstiziale, che altro non sono che vecchi osteoni in via di
demolizione in quel gioco di deposito e demolizione che corrisponde al pienamismo del t osseo. Il periostio:
questa membrana di t connettivo è tenacemente attaccato alla superficie dell’osso perchè se si staccasse con lievi
pressioni meccaniche il trofismo del t osseo se ne andrebbe a pallino allora ciuffi di fibre collagene dette
perforateli sciarpei entrano dentro il sistema circonferenziale esterno alle lamelle più esterne ed ancorano il t
osseo al periostio. Il periostio non c’è sulle superficie articolari dove c’è cartilagine ialina priva di pericondrio.
Capiamo ora la dinamica: quando si forma t osseo durante lo sviluppo embrionale fetale e anche dopo, si può
formare attraverso due meccanismi: l’ ossificazione cosiddetta diretta o membranosa con cui si formano durante
lo sviluppo embrionale la teca cranica e parte dello scheletro facciale e l’ossificazione indiretta o condrale con cui
si formano tutte le altre ossa. Per curiosità: il nostro embrione ha la copertura della testa all’inizio formata da una
membrana mesenchimale che man mano che le settimane di gestazione vanno avanti è soggetta ad ossidificazione
diretta o membranosa. Cioè direttamente dentro al mesenchima vi sorgono dei centri di ossificazione uno per
ognuna delle future ossa del cranio. Da li gli osteoblasti che si differenziano dalle cellule osteoprogenitrici a
ridosso dei vasi per formarsi il t osseo bisogna che ci siano vicino i vasi perchè la diffusione è negata e gli
osteociti devono arrivare nelle immediate vicinanze. Allora la vascolarizzazione, le cellule progenitrici, gli
osteoblasti e si forma t osseo che al centro di ognuna di quelle che saranno le future ossa si formano queste
trabecole di t osseo che crescono dentro la membrana che man mano non è più mesenchima ma è connettivo
fibroso denso più denso. Alla nascita quindi le ossa del cranio del neonato non sono saldate, il loro accrescimento
non si è ancora concluso e resta il connettivo a formare questi tralci e soprattutto queste zone dette fontanelle.
Quindi nel contesto di un t mesenchimale si sono create centri di ossidificazione diretta in cui direttamente da
cellule osteoprogenitrici si è creato t osseo. Dal centro si espande in periferia, si formano le varie ossa della testa
che al momento del parto non sono ancora saldate. Abbiamo capito cosa vuol dire ossificazione diretta o
membranosa.
L’altro evento è l’ossificazione indiretta o pondrale in cui da un abbozzo scheletrico cartilagineo che muore
seguendo quel modello si forma t osseo dentro una cartilagine che muore. Come fanno a morire? Le cellule della
cartilagine si dividono e quindi aumentano di numero facendo ingrandire la cartilagine poi diventano grosse
perché ipertrofizzano, cioè accumulano prodotti di riserva che noi sappiamo che il condrocita sa fare, dopo di che
cambiano e riversano i globuli calcificanti in una matrice cartilaginea che si mineralizza e quindi provocano la
morte delle cellule perché la diffusione non viene più portata avanti. Si creano dei buchi nella sostanza
intercellulare che vengono colonizzati da vasi che provengono da membrane cheriostali prima pericondriali poi
cheriostali vicine. I vasi portano le cellule osteoprogenitrici che a ridosso di un abbozzo di cartilagine morente
fanno t osseo. Il nostro scheletro fetale a parte la testa e poche ossa facciali e un pezzo della clavicola è fatto da t
cartilagineo che attraverso il meccanismo della morte delle sue cellule permette l’ingresso dei vasi e la formazione
di t osseo che cresce nelle varie direzioni aiutato da un’ossificazione diretta che avviene nel periostio. Questa
ossificazione è così detta periostatica. Cosi le ossa si ossificano, si accrescono con due meccanismi diversi, uno
dall’interno dentro la cartilagine che muore, uno dall’esterno in seno a una membrana connettivale che segue le
leggi dell’ossificazione diretta o membranosa. Ma noi sappiamo che le nostre ossa non solo durante lo sviluppo si
formano, si accrescono in larghezza, si forma un canale midollare ma si allungano fino a 18/20 anni. Sapete
perché ciò accade? Perchè in una zona chiamata metafisi tra l’epifisi prossimale e distale e la dialisi permane
cartilagine che prolifera in direzione verso la porzione più centrale della diafisi e man mano diventa ipertrofica,
calcifica, muore e viene invasa dall’osso. Questo meccanismo permette l’allungamento delle ossa fino a che con
la maggiore età le cartilagini non proliferano più, si chiudono e uno ha l’altezza dovuta alla sua genetica,
all’attività fisica che ha fatto, a come si è nutrito e via dicendo. Quindi noi abbiamo due tipi di ossificazione: la
diretta che ci fa nascere t osseo in membrana, mesenchimale durante lo sviluppo intrauterino, connettivo
propriamente detto dalla nascita in poi o encondrale cioè dentro una cartilagine che muore e fa da stampo perchè i
vasi che ci si infilano e che porta le cellule osteoprogenitrici fabbricano t osseo secondo il modello gia
prestabilito. (Le cellule della cartilagine muoiono e vengono tolte di mezzo e mineralizzano sostanza attorno alle
cellule cartilaginee e ne provoca la morte delle cartilagine. Guardando un’immagine: in rosa il t osseo, in azzurro
la cartilagine della sostanza intercellulare che è morta; le cellule non ci sono più, hanno lasciata gli spazi vuoti
come un groviera, i buchi sono ampiamente comunicanti, ci si sono infilati i vasi e hanno portato cellule
osteoprogenitrici che a ridosso della cartilagine che muore e che gli fa da stampo producono t osseo, prima t
fibroso e poi anulare. L cellule sono già morte della cartilagine la sostanza intercellulare man mano che l’osso
avanza, muore e se ne va e i macrofagi ripuliscono e tolgono di mezzo. Cresce qui t osseo in tutte le direzioni si
forma l’ipifisi si forma la diafisi resta cartilagine alla metafisi in grado di allungare con le divisioni per
atrofizzare, morire, essere invaso da t osseo, la possibilità dell’allungamento. Ora, noi sappiamo che l’osso
risponde a stimoli meccanici quali la compressione generalizzata, vuol dire che con l’attività fisica, la contrazione
muscolare opera su tutto lo scheletro, dove addome e varie, una trazione che induce il deposito della sostanza
mineralizzata dell’osso. Laddove c’è una gravità, già di per se c’è un meccanismo di questo tipo se noi siamo
inattivi o sdraiati per esempio, questo meccanismo invece è portato al meccanismo inverso cioè al riassorbimento,
ecco quindi che uno scheletro poco sollecitato dall’attività fisica e dalla stazione eretta è piu sottoposto a
riassorbimento che non a deposizione. Se poi noi abbiamo compressioni localizzate, quali apparecchi ortodontici,
apparecchi vari in quel punto riassorbe e dall’altro deposita e quindi lo scheletro si modifica come si tenta di fare
sulle mandibole o sulle mascelle dei ragazzini. L’osso risponde inoltre a stimoli di natura sistemica quali gli
ormoni, ci sono ormoni della crescita, gli ormoni tiroidei, paratormone, la calcitonina che invece porta al
riassorbimento dell’osso, a stimoli metabolici dovuti al fatto che esistono dei meccanismi nutrizionali che portano
all’interno maggiore del calcio o minore per esempio, a stimoli nutrizionali bisogna nutrirci apportando vitamine
nella nostra alimentazione e in quella del malato anche perche oltre a una dinamica positiva generale lo scheletro
ne viene a subire cose negative, esistono dei farmaci quali ad esempio alcuni antibiotici che interagiscono
negativamente con il metabolismo del t osseo. Allora io devo vedere che il metabolismo del t osseo è un continuo
dinamismo prevalendo nell’età dell’accrescimento e della formazione la deposizione del t osseo appunto per un
assetto ormonale, fisico, di movimento che porta a questo tipo di dinamica. Poi c’è un equilibrio vero tra deposito
e riassorbimento durante la vita adulta e nella fase invece della vecchiaia si ha un prevalere del riassorbimento.
Questo dinamismo può portare o a un vero e proprio rimodellamento dell’osso con vistosi cambiamenti di forma
per esempio quello che dicevamo prima la teca cranica,abbiamo il feto si formano le varie ossa non sono saldate
al momento del parto perché questo da un vantaggio nel passaggio del canale del parto e soprattutto permette con
nuova deposizione di sostanza un ampliamento della scatola cranica man mano che il bambino cresce e si fa
adulto. Ma non basta perché a un certo momento le suture avvengono, si chiudono, si suturano questi spazi e
allora ecco il meccanismo di ingrandimento della teca cranica erode dall’interno e applica sull’esterno, quindi
modifica la curvatura e aumenta il volume, permettendo all’encefalo di svilupparsi. Quindi attraverso il fenomeno
di riassorbimento dall’interno e deposito all’esterno, questa teca cranica muta la sua conformazione, la curvatura
aumentando il volume interno. Questo gioco di dinamicità porta ad un vero e proprio rimodellamento come nelle
ossa lunghe che man mano che il bambino procede dalla nascita poi la stazione eretta comincia a camminare, le
sue ossa si allungano e ingrandiscono in tutte le direzioni e il canale midollare si scava via via che lo ossa
crescono. Rimodellamenti li rivedo che sono diversi ma durante la vita adulta abbiamo rimaneggiamenti, io non
mi vedo cambiata ma le mie ossa cambiano in relazione agli stimoli ormonali, alla richiesta di calcio e alle forze,
quanto cammino, quanto mi muovo. Se sto sempre a letto il mio scheletro si riduce perché all’interno cosa
succede? Noi abbiamo detto che il tessuto è fatto da cellule gli osteociti che sono la forma adulta quindi si crede
che abbiano una nascita dopo il differenziamento dall’osteoblasto, una vita e una morte. L’osteocita è una cellula
programmata per morire prima o poi sotto stimoli fisico, nutrizionale, ormonale va in apoptosi e muore. Morendo
li crea una situazione perché l’osteoclasta arrivi e riassorba. Allora negli osteoclasti man mano nel giro di una
decina di anni si modificano sotto lo stimolo delle forze che vengono ad essere espresse, nutrizionali, di attività
fisica e per esempio possono passare da una struttura ad un’altra senza che il segmento scheletrico di per se si
modifichi macroscopicamente. Quindi attraverso il rimaneggiamento le ossa si rendono durante la vita disponibili
per l’omeostasi calcica, disponibile a un mutamento anche se non apprezzabile macroscopicamente alla variazione
di forze, allo stimolo ormonale che dura per tutto l’arco della vita.
Riprendo ora le nomenclature: Il t osseo puo essere rotto. abbiamo capito che viene rinnovato con questo
meccanismo della dinamicità ma possono accadere fatti traumatici o malattie che portano alla rottura del t osseo.
Allora innanzi tutto le aree danneggiate sulla linea di frattura vengono riassorbite e li l’apporto sanguineo viene a
mancare. Dalle tonache connettivali del cui terno si trova potenzialità di rifabbricare vasi e di ridare origine a
cellule osteoprogenitrici e quindi ad osteoblasti si ha la riparazione della frattura attraverso un processo diretto se
la frattura è composta o con le viti o per chi ha legno verde, dal periostio o dalle membrane interne si vanno a
ricostituire le parti rotte e quindi si ricrea la soluzione di continuità. Se invece la frattura è scomposta allora ci
possono essere due possibilità.
Guardando l’immagine: vedete muore il circolo sanguineo nella zona rappresentato in nero, quello attivo invece è
rosso. Qui abbiamo l’accorrere prima di una formazione nuova di vasi dopo di che dai margini di questa struttura
che porta il periostio esterno e l’endostio interno in tutte le varie cavità, si differenziano le cellule che vanno a
ricostituire il t osseo. Ma se la crescita dei vasi non è in sintonia e i vasi sono ancora lontani, queste membrane
connettivali si differenziano in cellule della cartilagine che sono in grado di assorbire per diffusione i metaboliti.
Quindi la ricostruzione del segmento osseo sarà preceduto dalla formazione di un callo cartilagineo, poi per
ossificazione encondrale dentro la cartilagine che verrà demolita si andrà a costituire t osseo successivo che
dapprima è un anello che congiunge la linea di frattura poi si avrà morte all’interno della struttura midollare delle
cellule che stanno ricostruendo t osseo e si ricreerà l’architettura primitiva del t osseo. Quindi durante le fratture
se è composta subito t osseo per ossificazione diretta o membranosa, se è scomposta e la crescita dei vasi nuovi
non va in sintonia con il differenziamento delle cellule, prima callo cartilagineo, poi per ossificazione indiretta o
condrale la formazione di t osseo.
ISTOLOGIA – 8° LEZIONE
Tratteremo in questa lezione il tessuto connettivo
specializzato nella funzione trofica del sangue e daremo
dei cenni sul tessuto muscolare.
Il sangue viene definito come un tessuto connettivo
altamente specializzato. Vedremo che la sua
specializzazione lo rende un connettivo sui generis
perché è costituito da una porzione corpuscolata, ovvero
i vari elementi del sangue che vengono anche chiamati
elementi figurati, e da una sostanza in cui questi elementi
si trovano, che è liquida e prende il nome di plasma.
Il plasma non è una tipica sostanza intercellulare, vuoi
perché è liquida e non è costituita dalle tipiche
componenti della sostanza intercellulare, cioè le fibre
della sostanza fondamentale anista, vuoi perché non è prodotta dalle cellule del sangue, cosa che invece accade
per i connettivi propriamente detti e le forme di sostegno in cui cellule specializzate (vedi fibroblasti e fibrociti
per il connettivo propriamente detto, condroblasti e condrociti per la cartilagine, osteoblasti e osteociti per il
tessuto osseo) depongono questa sostanza intercellulare e la tengono a controllo. Pertanto il sangue va inquadrato
nei limiti che abbiamo appena delineato.
Le funzioni del sangue sono molteplici. Ne vediamo le più importanti.
Trasporto dei gas respiratori.
Il
sangue trasporta l’ossigeno dall’alveolo polmonare ai tessuti, legando l’ossigeno all’emoglobina dei globuli rossi.
Dai tessuti all’alveolo polmonare porta l’anidride carbonica, ovvero il prodotto della respirazione cellulare (ne
abbiamo parlato a proposito dei mitocondri), anidride carbonica che è trasportata in quota parte legata ai globuli
rossi, e molta trasportata sottoforma di bicarbonati nel plasma. Quindi trasporto dei gas respiratori.
Trasporto
dei
cataboliti.
Trasporto, cioè, dei prodotti di rifiuto del metabolismo cellulare dai tessuti al rene, organo che si incarica della
loro eliminazione attraverso il filtrato renale, ovvero con la produzione di urina.
Trasporto
degli
ormoni.
Quando abbiamo fatto il tessuto ghiandolare endocrino, abbiamo sottolineato come gli ormoni abbiano come
veicolo per arrivare a tutti i tessuti e quindi a tutte le cellule bersaglio, il circolo ematico. Lo stesso accade per le
molecole regolatrici come le citochine, i fattori di crescita, tutte sostanze che le cellule riversano per armonizzare,
regolare la funzione reciproca, per richiamare cellule della difesa, per tenere a controllo la sostanza intercellulare
che è sostanza biologica, costituita da molecole biologiche, ma non vivente, quindi non autoregolante.
Trasporto delle cellule coinvolte nei processi di difesa.
Tutte le
cellule e, in particolare i globuli bianchi che sono coinvolti in questi processi, sono trasportati attraverso il circolo
sanguigno.
Inoltre le sostanze che vengono assorbite a livello intestinale, vengono portate come prima stazione al
fegato e da lì vengono poi portate al resto dell’organismo, intendendosi per tale, gli organi e, dunque, i tessuti che
vanno a costituire gli organi.
Siamo
animali
a
sangue
caldo;
il
sangue
termoregola.
Aiutato da altri meccanismi, mantiene costante la nostra temperatura.
Mantiene un equilibrio acido-base che è importantissimo per tutti i fenomeni metabolici, mantiene la
pressione osmotica ed oncotica controllando i fluidi tissutali nella loro concentrazione di proteine e del loro
contenuto in acqua (qualunque variazione porta a scompensi).
Quindi abbiamo definito il sangue e le sue funzioni che dobbiamo conoscere.
Ora vediamo la costituzione del plasma, cioè di quella che funge da
sostanza tra le cellule.
Il plasma è costituito in prevalenza da acqua, in quanto circa il 90% di
esso lo va a costituire. In esso sono disciolte diverse sostanze: da
ormoni ad enzimi, vitamine, componenti inorganici quali il calcio, il
sodio, il potassio e vi sono macromolecole organiche quali le proteine
e i lipidi. Tra le proteine, in massima parte prodotte dal fegato, si
possono ricordare:
1.
le albumine, importanti perché una loro variazione porta
sempre ad un richiamo di acqua da un versante all’altro e, negativa da
questo punto di vista, è il richiamo di acqua nei tessuti, con
formazione di edemi;
2.
le globuline, tra cui le gammaglobuline che sono gli anticorpi ed abbiamo già visto essere prodotte da
cellule differenziatesi nel connettivo, ma derivanti da precursori ematici. Gli anticorpi mediano l’immunità
umorale;
3.
le proteine della coagulazione e le proteine del complemento che sono l’insieme delle proteine che ci
danno l’indicazione dei fenomeni di difesa.
Infine, i lipidi: i tassi ematici ci indicano l’equilibrio che il nostro organismo sa tenere in relazione a queste
sostanze.
Un altro dato importante è che tra plasma ed elementi figurati esiste un rapporto quantitativo che è indice di buona
funzionalità. Questo si chiama “valore ematocrito” e ci dice che il plasma corrisponde circa al 55%, mentre gli
elementi figurati sono il 45%, di cui il grosso è dato da globuli rossi e dalle piastrine e, solo una minima parte, è
data dai globuli bianchi. Qualunque variazione di questo rapporto è indicativo, vuoi della funzionalità del nostro
midollo, vuoi di situazioni periferiche e, ultimamente, è stato indice sebbene molto generale, di rilevazione di
processi di doping poiché qualunque sostanza (per esempio l’eritropoietina che innalza la funzionalità midollare
per incrementare le potenzialità fisiche) viene rivelata da questa percentuale.
Prendiamo in considerazione gli elementi figurati che sono suddivisi in globuli rossi, globuli bianchi o leucociti e
piastrine.
I globuli bianchi sono a loro volta suddivisi in due categorie: i leucociti o globuli bianchi granulosi e i globuli
bianchi non granulosi.
Tra i granulosi abbiamo i granulociti divisi in tre categorie a seconda delle affinità tintoriali specifiche in
neutrofili, acidofili e basofili.
Quelli non granulosi, anche se granuli ne hanno, non hanno granuli specifici, e sono i monociti ed i linfociti.
Infine le piastrine.
Bisogna sapere le dimensioni degli elementi figurati del sangue, le concentrazioni per millimetro cubo perché
ogni variazione in più o in meno è indice di patologia o comunque di funzionalità non perfetta e le caratteristiche
morfologiche che sono alla base della comprensione della funzione.
Il sangue è una delle domande che all’orale avremo quasi tutti.
I GLOBULI ROSSI
Partiamo dai globuli rossi: il diametro è di circa 7,2 micrometri, con
concentrazione di circa 5 milioni per millimetro cubo nell’uomo e 4,5 milioni per
millimetro cubo nella donna, quindi vi è una diversità tra i due sessi.
Le caratteristiche sono le seguenti:
i globuli rossi sono degli elementi a differenziamento ultimo;
sono privi di nucleo e di organuli quando sono pienamente maturi;
sono costituiti da una membrana molto studiata perché, essendo delle strutture prive di organuli e di
nucleo, si prestano bene ad un certo tipo di indagine.
(Lei preferisce chiamarli globuli rossi e non eritrociti perché non sono cellule vere e proprie).
La membrana dei globuli rossi presenta proteine integrali (molto studiate per il trasporto di ioni) e glicoproteine di
membrana che sono alla base della costituzione genetica dei gruppi sanguigni A, B, 0 ed RH. Il citoscheletro, in
questa forma così differenziata, mantiene le sue caratteristiche, conferendo ai globuli rossi, una forma particolare,
elasticità e plasticità. I globuli rossi possono passare, pur essendo di 7,2 micrometri, anche in sezioni della rete
capillare molto più sottile, di 3-4 micrometri, il che vuol dire che sono plastici. Il contenuto tipico di questo
elemento particolare è l’emoglobina, una cromoproteina che, in alcune patologie, può presentare delle anomalie di
costituzione che sono alla base di un disturbo funzionale, spesso grave, che può portare ad una distruzione
precoce dei globuli rossi.
Ad esempio l’anemia è sempre indice di una riduzione drastica dei globuli rossi circolanti dovuta a cause varie: il
midollo che non li produce, una distruzione periferica altrettanto precoce o attivata in maniera non congrua,
dovuta a cause accidentali o a cause di emoglobina non perfettamente costituita (come avviene nell’anemia
falciforme, la quale è geneticamente determinata).
Figura: vetrino con una goccia di sangue strisciata, colorata ed osservata.
I globuli rossi sono dei dischetti che si presentano più colorati nella porzione periferica (osserviamoli laddove non
sono raggruppati, ma dove sono singoli). La porzione centrale è, invece, più chiara. Questo è dovuto al fatto che il
globulo rosso ha una forma molto particolare, cosiddetta a lente biconcava, una specie di disco in cui la parte
centrale è più sottile e per questo più bianca nello striscio. Questa forma a dischetto appiattito che forma una
specie di ciambella periferica più spessa, nella microscopia elettronica mostra il suo profilo inconfondibile.
Questa forma si è selezionata come estremamente congrua allo scambio gassoso in quanto espone un elevato
rapporto tra superficie e volume. Se, infatti, prendiamo una palla delle stesse dimensioni della nostra lente
biconcava, noteremo che il rapporto tra la superficie ed il volume è maggiore in quest’ultima. Quindi, in questo
caso, l’emoglobina è maggiormente espressa per lo scambio gassoso. Inoltre se avesse avuto la forma di una palla,
l’emoglobina contenuta centralmente avrebbe incontrato qualche difficoltà nello scambio gassoso.
Bisogna sapere quindi che la forma a lente biconcava è estremamente adatta allo scambio gassoso in quanto
esprime un elevato rapporto tra superficie e volume.
Qualunque variazione della dimensione dei globuli rossi viene chiamata sferocitosi (?) e viene detto macrocita il
globulo rosso grosso oltre i 9 micrometri, microcita quello al di sotto di 6 micrometri. Anche le variazioni di
forma possono essere indice di malattia o patologia o comunque di qualcosa che si sta alterando. Per esempio i
globuli rossi che hanno una superficie non liscia, ma dentellata sono espressione di una disidratazione, mentre i
globuli rossi a falce, sono l’espressione di un’emoglobina malformata. Quindi le variazioni di forma, di
dimensione e di numero sono sempre espressioni di qualcosa che non va, cioè di qualcosa che è extra-fisiologico.
Quali sono le funzioni dei globuli rossi?
Trasporto gassoso: all’interno dei vasi sanguigni trasportano ossigeno che si lega all’emoglobina, anidride
carbonica (solo il 20% circa è trasportato dai globuli rossi sottoforma di carbossiemoglobina, mentre l’80% è
trasportato nel plasma sottoforma di bicarbonati).
Derivazione: tutti gli elementi del sangue circolanti derivano, come avevamo detto mentre discutevamo delle
generalità sul tessuto connettivo, da un precursore midollare, una cellula staminale ematopoietica che derivava dal
differenziamento, in questi luoghi, della cellula mesenchimale. La cellula staminale emopoietica, che si trova a
livello del midollo rosso, è totipotente, tuttavia essa, man mano che si divide, dà origine a popolazioni cellulari
diverse che, attraverso divisioni e differenziamento, danno origine alle forme ematiche. Quindi, tutte le cellule che
troviamo a livello del sangue circolante sono il risultato del differenziamento, attraverso divisioni successive, di
cellule che stanno nel midollo. Si chiama ematopoiesi il processo attraverso il quale da cellule midollari derivate
tutte da una cellula primordiale staminale, si attua il differenziamento nelle forme mature.
Quindi alla domanda “da chi derivano i globuli rossi” bisogna rispondere che essi derivano dal differenziamento
midollare.
In circolo non hanno più la potenzialità di dividersi, cioè sono a differenziamento ultimo e sono destinate ad
essere eliminate quando invecchiano.
Essi derivano dal midollo rosso emopoietico e il progenitore più antico (nel senso di divisione) proeritroblasto.
La vita media dei globuli rossi è circa 120 giorni, vengono poi distrutti dai macrofagi lienali, cioè quelli della
milza e del fegato, per fagocitosi.
Riepiloghiamo quanto detto finora:
la derivazione → a livello del midollo rosso si posso identificare cellule dette proeritroblasti come progenitori più
anziani dei globuli rossi circolanti. Queste cellule, ovvero i proeritroblasti, attraverso divisioni successive, quindi
più generazioni cellulari, si differenziano. All’inizio, il proeritroblasto è una cellula con un nucleo e molti
ribosomi, poi comincia a fabbricare tanta emoglobina riducendo i ribosomi che intanto la fabbricano;
successivamente perde il nucleo , perde gli organuli e diventa globulo rosso, cioè un sacchetto con una membrana
estremamente specializzata e con il citoscheletro pieno di emoglobina e, a questo punto, esso va in circolo. E’
caratterizzato da una forma a lente biconcava. Esiste un periodo in cui i globuli rossi circolanti, appena immessi,
possono mantenere dei residui di ribosomi e mitocondri o porzioni del nucleo e questi sono dei globuli rossi
giovani chiamati reticolociti. I reticolociti sono quindi dei globuli rossi immaturi che conservano la parte di
ribosomi, ma non devono essere più del 1 % dei globuli rossi circolanti. Nel fare l’analisi del sangue a pazienti
con problemi, ad esempio di anemia, si possono dosare i globuli rossi immaturi in quanto indice generico di buon
funzionamento o meno del midollo.
Il globulo rosso maturo è privo di qualsiasi tipo di colorazione che non sia un’acidofilia; invece i globuli rossi
immaturi, reticolociti, presentano dei reticoli all’interno del citoplasma (da cui deriva il nome).
I GLOBULI BIANCHI O LEUCOCITI.
La loro concentrazione varia tra i 4000 e gli 8000 per millimetro cubo, con una
distribuzione percentuale dei vari tipi di globuli bianchi che prende il nome di
formula leucocitaria. La formula leucocitaria è il primo indizio di funzionalità
buona o di alterazione, quindi ha valore diagnostico, seppur generico (VA
SAPUTA!). I granulociti neutrofili variano tra il 45 e il 75 %, per alcuni libri tra 50
e 70%, quindi noi riterremo giusto quest’ultimo intervallo.
I granulociti eosinofili sono l’1-5 %, mentre i basofili lo 0,5-1%.
I linfociti, che sono la seconda popolazione cellulare per concentrazione, tra il 20 e
il 45 %, i monociti il 2-10%.
I globuli bianchi non esplicano le proprie funzioni solo nei vasi, come i globuli
rossi, ma possono passare la parete endoteliale, con movimento ameboide, dando luogo alla diapedesi per poi
andare nei connettivi dove, o mantenendo le proprie caratteristiche o trasformandosi in cellule del connettivo,
danno origine ad elementi che attivamente portano avanti la funzione. Mentre i globuli rossi stanno
fisiologicamente dentro i vasi e ne fuoriescono solo per rottura di vasi e quindi per emorragia; i globuli bianchi
valicano la parete endoteliale, perché l’endotelio li riconosce attraverso una serie di sostanze che vengono
prodotte, questi si fanno varco tra le cellule endoteliali e scivolano fuori, andando a colonizzare il connettivo.
I globuli bianchi sono alla base di quella che si chiama immunità ed essa viene distinta in aspecifica (detta anche
naturale o innata) e specifica. L’immunità aspecifica coinvolge principalmente i granulociti, i macrofagi, le
proteine del complemento e la barriera costituita dalle membrane mucose. Questa immunità si instaura durante i
processi infiammatori, le ustioni, i traumi, le riparazioni delle ferite. L’immunità specifica, invece, è acquisita e
dipende, in maniera non naturale, da un contatto con un antigene, cioè con una sostanza estranea e viene portata
avanti dalle popolazioni linfocitarie.
Parliamo ora delle diverse popolazioni di globuli bianchi:
I granulociti. Abbiamo già detto che i globuli bianchi si dividono in granulosi e non granulosi in base alla
presenza o meno di granulazioni specifiche e sottolineo specifiche perché le granulazioni aspecifiche, chiamate
anche granuli azzurrofili ed hanno il significato di lisosomi, ce li hanno un po’ tutti i globuli bianchi. Gli elementi
cosiddetti granulosi, ovvero i granulociti, hanno sia granulazioni aspecifiche dette anche primarie perché per
prime vengono elaborate nel processo di differenziamento, che sono i lisosomi, sia granulazioni specifiche che
hanno attività tintoriali specifiche e da cui derivano i nomi dei vari granulociti. Quindi riconosceremo i granulociti
neutrofili che hanno affinità, cioè si colorano, con sali neutri (sono gli eosinati di blu di metilene e toluidina). Non
importa sapere questo, ma bisogna sapere su che basi si distinguono i granulociti nelle tre categorie citate.
Sottolineo nuovamente che essi vengono suddivisi in base alle affinità tintoriali conferite loro dalle granulazioni
specifiche. Questo vuol dire che i granulociti neutrofili sono dotati di granulazioni specifiche di un certo tipo,
quelli acidofili di un altro tipo e quelli basofili di un altro ancora. I granulociti neutrofili si colorano con i sali
indissociabili, eosinati del blu di metilene o degli altri, i granulociti acidofili sono detti anche eosinofili perché si
colorano con un colorante acido, rosa, detto cosina, e i granulociti basofili, invece, si colorano con i coloranti
basici, vedi il blu di metilene. Tutti i granulociti, che hanno dimensioni variabili tra i 12 ed i 14 micrometri (sono
quindi cellule più grandi dei globuli rossi), hanno un nucleo detto polilobato. Ciò significa che esso alterna zone
dilatate a zone ristrette nel proprio materiale nucleare in modo tale da dare l’idea di lobature (se mi passate il
paragone, sembrano una fila di salsicce). Questo è massimamente espresso nei granulociti neutrofili e vedremo
meno espresso negli acidofili e nei basofili. Il loro precursore midollare si chiama mieloblasto, hanno vita media
di 7-12 giorni, esplicano la loro funzione nei connettivi e da qui vengono poi eliminati per essere ripristinati da
nuove ondate di derivazione midollare.
Figura: granulocita neutrofilo. Si vede un citoplasma chiaro con dei granuli piccoli e molte lobature.
Più invecchia nei suoi 7 giorni, più le lobature aumentano.
L’eosinofilo ha al massimo due lobature e granuli più grossi.
Il basofilo ha al massimo due lobature, granulazioni molto grosse che nello striscio mascherano il nucleo.
Figura: i granulociti neutrofili hanno come tutti i granulociti una forma globosa, nello striscio
rotondeggiante, si vedono dalla figura le lobature del nucleo e le zone ristrette, con un 20% di granulociti
neutrofili di individui di sesso femminile, dove si vede un piccolo lobo aggiuntivo molto sottile che viene detto
bacchetta di tamburo o, in inglese, drumstick, poiché ha una porzione slargata ed una ristretta e si attacca ad una
delle lobature che è il corpo di bar delle cellule somatiche, cioè una delle due x del corredo cromosomico
femminile inattiva. I granulociti neutrofili sono un buon substrato dove poter mettere in evidenza questa
caratteristica femminile. Essa non appare sempre perché lo striscio può nasconderla, ma laddove appare, vuol dire
che siamo in presenza di un individuo di sesso femminile. Anche nei neuroni che hanno un nucleo particolarmente
chiaro è possibile vedere la zolla eterocromatica della x femminile.
Figura: bacchetta di tamburo con forma slargata e piccolo peduncolo che la lega ad uno dei lobi
Le lobature aumentano con l’invecchiamento dei granulociti ed esiste una formula, detta formula di Arneth, per
mettere in evidenza la percentuale di granulociti vecchi con tante lobature. Anche questo è un altro indice di
funzionalità midollare.
Quindi nel sangue ci sono tante potenzialità istologiche, anche se generiche, che permettono di evidenziare
patologie latenti o meno.
La funzione dei granulociti neutrofili è portata avanti dalle granulazioni specifiche che, in alcuni libri, vengono
suddivise in granulazioni secondarie e terziarie e grosso modo contengono, oltre ad enzimi antibatterici, le
gelatinasi e sono particolarmente adatti alla lotta contro infezioni di natura batterica. Morendo in questa loro lotta
contro i batteri, anche in condizioni prive di ossigeno, danno origine al pus che poi viene risolto ed eliminato da
altre cellule della difesa quali i macrofagi ed i linfociti. I granulociti neutrofili, tra tutti i granulociti, sono quelli
che hanno capacità di fagocitosi.
L’eosinofilo ha una forma sferica, nucleo bilobato, presenta granulazioni specifiche acidofile e anch’esse
fagocitano. Ad esempio sono addetti alla fagocitosi del complesso antigene-anticorpo. Gli anticorpi sono infatti
prodotti in risposta ad un antigene, l’anticorpo si lega all’antigene e lo disattiva ed il granulocita acidofilo leva di
mezzo il complesso antigene-anticorpo. Inoltre hanno una funzione di difesa nelle infestazioni cioè quando siamo
aggrediti da parassiti. Se una formula leucocitaria ha una variazione dei granulociti eosinofili, può darsi che un
individuo abbia subito un’infestazione, cioè sia stato aggredito da un parassita, ad esempio dalla tenia.
Figura:granulocita eosinofilo. I granuli del suo citoplasma hanno lo stesso colore dei globuli rossi, cioè si
colorano con l’eosina. Il nucleo presenta al massimo due lobature. E’ visibile la porzione sottile che le connette
Granulociti basofili. Forma sferica, nucleo bilobato, le granulazioni specifiche voluminose sono basofile. Queste
granulazioni basofile contengono eparina ed istamina, quindi hanno granuli simili ai mastociti, le cellule della
difesa che entrano in gioco nelle reazioni allergiche. I granulociti basofili collaborano con i mastociti alla risposta
nelle allergie provocando riniti, orticaria, asma fino all’evento ultimo, cioè lo shock anafilattico.
Figura: granulociti basofili. I granuli basofili quasi coprono il nucleo bilobato
I monociti sono una delle due forme non granulose, sono sferici, hanno un nucleo voluminoso, spesso
reniforme, cioè a forma di rene con un’intaccatura, vita media di 2-3 giorni, essi migrano attraverso la parete
basale e si differenziano nel sistema dei macrofagi dando origine a tutti gli elementi di questo sistema attraverso la
differenziazione in loco.
Figura: il nucleo del monocita che può avere un’intaccatura dando origine ad una forma quasi reniforme che
talvolta può essere più accentuata.
Figura: striscio di sangue con dei monociti, ovvero cellule rotonde nello striscio e, dunque, globose, sferiche
nelle tre dimensioni, il nucleo con l’intaccatura e reniforme, un grosso apparato di Golgi e qualche granulo
azzurrofilo, cioè i granuli primari di cui tutti gli elementi della serie bianca sono costituiti .I granuli azzurrofili
sono i lisosomi.
Gli elementi che corrispondono ai linfociti possono presentarsi di piccole dimensioni, 6-9 micrometri o grandi
9-15. In circolo si trovano i piccoli linfociti. Trovare in circolo quelli di grandi dimensioni è sintomo di patologie.
La forma è sferica, il nucleo è anch’esso sferico, il citoplasma è molto ridotto e presenta ribosomi, reticolo,
apparato di Golgi e pochi granuli azzurrofili.
I linfociti si dividono in due popolazioni distinte chiamate con le lettere dell’alfabeto B e T. B deriva da Bone
Mellow (?) ora, storicamente da bursa fabricii, borsa di fabrizio che è un organo ematopoietico degli uccelli dove
furono messi in evidenza per la prima volta. Ora rimane la dicitura B che deriva da Bone Mellow (midollo osseo).
Questo deriva dal fatto che i linfociti B si differenziano da un precursore indifferenziato nel midollo osseo. Poi
vengono messi in circolo per passare poi nei connettivi.
I linfociti T derivano il loro nome dal timo, un organo molto sviluppato durante l’adolescenza, che regredisce
dando origine a questa popolazione linfocitaria che si denonima con la prima lettera di timo
I linfociti B operano la difesa umorale attraverso il differenziamento in plasmacellule e la produzione di anticorpi.
I linfociti T operano la difesa cellulo-mediata attraverso una serie di differenziamenti in cellule tutte facenti capo
a questa popolazione chiamata linfociti T.
I linfociti B riconoscono gli antigeni nella loro conformazione naturale e questo riconoscimento fa sì che loro
espongano sulla superficie dei recettori che riconoscono questi antigeni. Si dividono tante volte, cioè blastizzano,
vanno nel connettivo, si trasformano in plasmacellule e producono anticorpi sullo schema di quei recettori che
avevano riconosciuto gli antigeni. Gli anticorpi in giro trovano l’antigene, ci si legano e lo disattivano.
Quindi dobbiamo sapere come sono fatti i linfociti B e T, cosa sono, da dove derivano e quali funzioni portano
avanti.
Il linfocita B ha riconosciuto un antigene nella sua forma nativa, va nel connettivo, si trasforma in plasmacellula,
la plasmacellula attraverso il suo sistema di reticolo e Golgi produce gli anticorpi su quello schema che il linfocita
B aveva costruito, li emette, cioè li esocita e questi riconoscono l’antigene e ci si legano. Il complesso antigeneanticorpo verrà tolto di mezzo dai granulociti eosinofili.
I linfociti T riconoscono cellule alterate e sostanze che hanno valore antigenico, ma solo se legato agli antigeni di
istocompatibilità, cioè hanno bisogno di una presentazione. Inoltre aiutano la risposta dei linfociti B. A livello dei
linfociti T si possono riconoscere sottopopolazioni che vengono chiamate linfociti citotossici, helper e suppressor
ad indicare che esistono popolazioni cellulari linfocitarie (linfociti citotossici) capaci di emettere tossine che
lisano le cellule alterate o comunque non riconoscibili come nostre; i linfociti helper sono quelli che collaborano e
richiamano altre cellule della difesa tra cui anche i linfociti B; i suppressor (alcuni libri negano che siano una
popolazione a se stante) rappresentano cellule che limitano la risposta immunitaria perché la risposta immunitaria
deve avere delle caratteristiche: dev’essere specifica, cioè contro antigeni, pronta e deve poter conservare in
alcuni casi una memoria, ma deve essere anche autolimitante altrimenti la distruzione andrebbe oltre i limiti
coinvolgendo anche strutture che non devono essere coinvolte. Inoltre la risposta non deve essere perpetuata
perché tutte le risposte immunitarie di difesa portano a lesioni del connettivo e quindi una risposta che si prolunga
può avere come esito finale la fibrosi, cioè un’iper produzione di sostanza intercellulare del connettivo. Tra le
cellule linfocitarie esiste anche una popolazione detta natural killer (killer naturale) che distrugge le cellule
tumorali. Si tratta di una citotossicità spontanea. Queste cellule intercettano le cellule modificate in senso
neoplastico e le tolgono di torno.
Quindi i linfociti si distinguono in linfociti di tipo B, T e natural killer:
linfociti B risposta umorale attraverso le plasmacellule;
linfociti T cellule mediate attraverso i citotossici, helper, suppressor con il coinvolgimento dei macrofagi e di tutte
le altre cellule della difesa;
i natural killer.
Le piastrine sono frammenti cellulari che derivano dalla suddivisione del citoplasma, dei cosiddetti
megafagociti, cioè cellule midollari. La concentrazione nel sangue è di circa 200mila millimetri, la vita media di
5-9 giorni, hanno una zona centrale più colorata.
Figura: megafagociti. Hanno una zona centrale più colorata e quella periferica, lo ialomero, meno colorata.
Queste piastrine hanno un ruolo nella emostasi, cioè nel fermare il sangue in un vaso leso e nella coagulazione
attraverso l’adesione alla parete basale lesa, l’aggregazione e la secrezione di fattori che entrano in gioco nella
coagulazione.
Figura: piastrine adese alla parete di un vaso leso. Aderiscono, formano un tappo nel quale rimangono
ingabbiati i globuli rossi impedendone la fuoriuscita; inoltre, partecipando al meccanismo che porta dalla
protrombina a trombina, agiscono sul fibrinogeno che è una proteina solubile rendendola insolubile in fibrina, che
partecipa alla coagulazione del sangue.
9 Lezione istologia
TESSUTO MUSCOLARE
I tessuti muscolari sono tessuti specializzati nel fenomeno della contrazione, grazie alla presenza, nel citoplasma
degli elementi che li costituiscono, di proteine contrattili; dunque, il differenziamento di questi elementi porta
prima a fabbricare proteine del tipo contrattile e poi ad organizzare queste proteine in strutture capaci di mediare
la contrazione.
Distinguiamo due tipi di tessuto muscolare:
Striato
Liscio
TESSUTO MUSCOLARE STRIATO
La denominazione deriva dal fatto che il tessuto muscolare striato ha elementi che presentano, al microscopio
ottico e al microscopio polarizzatore, un’alternanza, lungo il decorso degli elementi costitutivi, di bande (zone più
colorate che si alternano a zone meno colorate).
Questa bandeggiatura o striatura ha dato il nome a questo tipo di muscolatura. Responsabili della bandeggiatura
sono strutture chiamate “miofibrille”, che sono il substrato morfologico della contrazione e sono costituite da
filamenti contrattili.
Il tessuto muscolare striato si divide in due categorie:
Tessuto muscolare striato di tipo scheletrico (il tessuto che va a costituire i muscoli volontari )
Tessuto muscolare striato di tipo cardiaco (il tessuto che va a costituire la muscolatura involontaria del
cuore )
TESSUTO MUSCOLARE STRIATO DI TIPO SCHELETRICO
È dunque quello che va a costituire unità anatomiche come muscoli scheletrici, quelli responsabili dei movimenti
volontari che attraverso tendini, aponeurosi, si legano al segmento osseo, la lingua, i muscoli mimici e i muscoli
estrinseci dell’occhio.
L’elemento costitutivo di questo tessuto è chiamato “fibra muscolare”. Le fibre sono strutture allungate che
possono arrivare a qualche cm di lunghezza a seconda del tipo di muscolo preso in considerazione, hanno forma
irregolarmente cilindrica e l’estremità di questi cilindri è lievemente affusolata.
Se sezioniamo la fibra trasversalmente essa appare rotondeggiante, mentre, se la guardiamo longitudinalmente,
appare come un lungo nastro.
È proprio nella sezione longitudinale che è visibile la striatura.
SEZIONE TRASVERSALE
SEZIONE LONGITUDINALE
La fibra contiene molti nuclei, parecchie centinaia, dislocati in periferia (sotto la membrana ); infatti è un esempio
di struttura polinucleata, nata attraverso la modalità a “sincizio”, cioè, durante lo sviluppo embrionale fetale, più
cellule (chiamate mioblasti ) si sono unite fra loro, hanno eliminato le separazioni citoplasmatiche, creando un
territorio citoplasmatico unico, in cui si trovano tanti nuclei.
I nuclei, inizialmente centrali, si portano ,durante lo sviluppo e differenziamento, sotto la membrana plasmatica e
la porzione centrale della fibra viene occupata da un prodotto di sintesi della fibra stessa che ha il carattere di
incluso, le miofibrille, responsabili della bandeggiatura e che quindi costituiscono il substrato morfologico della
contrazione.
Le fibre tendono a disporsi una accanto all’altra, quasi in un registro parallelo, senza sviluppare nessun tipo di
giunzione tra di loro anzi, ciascuna fibra è circondata da una membrana basale (detta endomisio) che la separa e la
unisce a quelle vicine. Viene chiamata “sarcolemma” la membrana della fibra muscolare insieme alla lamina
basale (quest’ultima viene secreta dalla fibra stessa).
Viene chiamato “sarcoplasma” il citoplasma della fibra muscolare.
Dunque i nuclei si trovano al di sotto del sarcolemma. Vicino a ciascun nucleo, il citoplasma presenta un
accumulo di organuli in cui possiamo distinguere un apparato di Golgi, cisterne di reticolo ruvido, ribosomi liberi,
mitocondri, reticolo liscio: questo si ripete ogni volta che incontriamo un nucleo, perché ogni nucleo controlla una
certa parte di citoplasma.
Il reticolo liscio e i mitocondri si ritrovano anche e soprattutto tra le miofibrille, oltre che vicino il nucleo. Le
miofibrille abbiamo detto sono degli inclusi. Come inclusi possiamo trovare inoltre granuli di glicogeno e gocce
di lipidi, come materiale di riserva. Più fasce di fibre si organizzano tra loro con connettivo, che si chiama
“perimisio” (è un fibrillare lasso), a formare il muscolo, che è circondato da una membrana di fibroso denso a
fasci intrecciati che è l’“epimisio”. Dunque,riepilogando, la composizione del muscolo è la seguente:
partendo dall’esterno troviamo l’epimisio, ovvero l’involucro del muscolo, che lo protegge dagli stimoli
meccanici che vengono applicati in superficie.
L’epimisio è in una forma densa. Dall’epimisio si dipartono setti connettivali che scompongono il muscolo in
tanti fascetti di fibre muscolari; questo connettivo diventa sempre più delicato ( connettivo lasso ) e prende il
nome di perimisio ; al suo interno si ritrovano vasi arteriosi, venosi, linfatici e nervi. Le parti ancora più interne
del perimisio si continuano con una delicata rete di connettivo ancora più sottile che forma un rivestimento (
l’endomisio ) intorno ad ogni singola fibra muscolare .
L’endomisio è a diretto contatto con la membrana della stessa fibra muscolare, il sarcolemma
STRIATURA DELLA FIBRA
Le righe chiare e scure che rappresentano la striatura della fibra sono dette “bande” o “dischi”
La banda A appare al MO più colorata della banda I.
La sequenza è quindi:
banda A molto colorata, banda I meno colorata che si ripetono lungo tutto il decorso della fibra.
Al centro della banda I si trova una linea scura, la linea Z.
La banda A non appare uniformemente colorata, ma presenta una parte centrale meno colorata, detta banda H,
segnata al centro dalla linea M.
Definiamo “sarcomero” la porzione che va da una linea Z a quella successiva, comprendente un emidisco I , un
emidisco A , un altro emidisco I che si ripetono periodicamente lungo il decorso della fibra.
La bandeggiatura, come vediamo dall’immagine, interessa solo le miofibrille ma poiché esse, all’interno della
fibra, sono “in registro” (cioè tutte le linee Z,tutte le bande A ,etc, sono alla stessa altezza), la bandeggiatura della
miofibrilla diventa bandeggiatura della fibra muscolare.
MIOFIBRILLE
Le miofibrille sono strutture disposte longitudinalmente rispetto al decorso della fibra e sono visibili, come
strutture a sé, solo al microscopio elettronico.
Appaiono come funi al cui interno si trovano altri elementi, i “miofilamenti”, i quali si distribuiscono in modo
diverso, determinando la formazione delle varie zone appena descritte.
I miofilamenti sono di due tipi :
Sottile
Spesso
Il miofilamento sottile è costituito da actina; l’actina sarcomerica è di isoforma diversa da quella ubiquitaria dei
microfilamenti citoscheletrici non muscolari, ma la struttura è uguale, globulare: le molecole di actina si uniscono
a formare 2 fili che si attorcigliano: la F-actina
Nel miofilamento sottile si trovano, oltre all’actina, altre due proteine: la troponina e la tropomiosina.
Quest’ultima attorciglia il primo filo dell’actina e si lega alla troponina attraverso un sito di riconoscimento; la
troponina, poi, si lega anch’essa all’actina e avrà un altro sito disponibile al calcio.
Riepilogando, il miofilamento sottile è costituito da:
Actina di tipo sarcomerico
Tropomiosina
Troponina
La troponina tiene insieme le prime due e riconosce il calcio.
Il miofilamento spesso è dato dalla miosina, quella proteina che interagisce sempre con l’actina per provocare la
contrazione.
La miosina ricorda una mazza da golf; presenta una porzione slargata, detta “testa” e una allungata, sottile (sono
la parte pesante e leggera della miosina).
A formare il filamento spesso, le mazze da golf si riuniscono attraverso le porzioni sottili, facendo emergere le
teste da questo insieme.
Ora mettiamo filamento sottile e filamento spesso all’interno della miofibrilla, cioè dentro il sarcomero.
Quando il filamento spesso si va a collocare dentro il sarcomero abbiamo la congiunzione dei filamenti spessi,
attraverso le porzioni sottili della miosina, al livello della linea M; successivamente si dirigono specularmente
esponendo le teste nelle porzioni periferiche della banda A. Quindi la zona H è occupata dalla miosina senza testa.
L’actina dove si trova? Dove si trova cioè il filamento sottile?
Esso parte dalle linea Z, cioè a metà della banda I e si dirige specularmene ad occupare tutto il disco I, ma non gli
basta ed entra, infatti, anche nelle porzioni periferiche del disco A, laddove ci sono le teste della miosina.
Sarà infatti il sinergismo tra le teste della miosina e l’actina a provocare la contrazione ( lo scorrimento dei
filamenti ). Ricapitolando:
Il disco I contiene solo filamenti sottili, le porzioni periferiche del disco A
contengono entrambi i tipi di filamento, la porzione lineare della banda H contiene solo filamenti spessi; la linea
M , invece, non è altro che una zona di raccordo tra i filamenti spessi degli emidischi A e tra loro stessi: questo è
il sarcomero.
Gli elementi scheletrici appartenenti alla categoria intermedia, nella fibra muscolare sono tantissimi.
Quale funzione hanno?
Legano nella parte superiore e nella parte inferiore le miofibrille e quindi sono solidali con la membrana
plasmatici
Tengono le linee Z tutte alla stessa altezza e le legano alla membrana laterale.
Legano il disco A a varie componenti del citoscheletro.
Raccordano le varie parti tra loro.
Quindi hanno un ruolo estremamente importante.
Ricordiamo , tra questi, la “desmina” che tiene le linee Z tutte alla stessa altezza e la “distrofina”, che appartiene
alle proteine citoscheletriche intermedie e che lega alla membrana ed agli altri elementi del citoscheletro la
miofibrilla.
Esiste una malattia, la distrofia muscolare, che è data da una cattiva struttura della distrofina; una distrofina non
ben congegnata non gioca un ruolo positivo nella trasmissione della forza contrattile e non previene gli effetti
traumatici di contrazioni troppo forti, il che vuol dire che i muscoli delle persone affette da distrofia muscolare
vanno incontro ad una degenerazione che rende prima molto faticoso il movimento poi lo impedisce totalmente.
RETICOLO SARCOPLASMATICO
Le fibre muscolari hanno molto sviluppato, oltre ai mitocondri ( e questo è capibile in quanto i mitocondri
forniscono energia, ed essendo la contrazione muscolare un lavoro attivo,richiede molto ATP ), il reticolo liscio (
che prende il nome di reticolo sarcoplasmatico ).
Lungo il decorso della miofibrilla si trovano dei tubuli del reticolo liscio che, a mò di calza, circondano la
miofibrilla stessa .Questi tubuli, cioè, ogni tanto confluiscono insieme e formano delle cisterne che circondano la
miofibrilla a livello del punto di passaggio tra disco A e disco I. Si formano due cisterne ravvicinate ogni volta
che il disco A passa nel disco I e viceversa ( due per ogni sarcomero ).
Alla confluenza di due cisterne terminali è presente, tra le due cisterne una formazione tubulare detta tubulo T, si
tratta di una invaginazione del sarcolemma il cui lume comunica con l'ambiente extracellulare ma non con il lume
del reticolo sarcoplasmatico.
Quest’introflessione ,infilandosi tra le cisterne, forma una struttura a tre oggetti chiamata “triade”.
La muscolatura striata si contrae in risposta ad uno stimolo volontario che avviene ad opera di motoneuroni che si
trovano nel midollo.
La fibra muscolare, dunque, riceve una specie di informazione, la contrazione, che corre sulla membrana,
trovando i tubuli T, i quali a loro volta passano l’informazione alle cisterne , con le quali sono a contatto.
L’informazione che arriva alle cisterne blocca pompe di membrana al calcio che sono attive fino a quando non
arriva l’impulso alla contrazione pompando calcio all’interno dei tubuli del reticolo liscio;all’arrivo dell’impulso
queste pompe al calcio si inibiscono e il calcio dilaga nel citoplasma, dove incontra la troponina (sensibile al
calcio ), la quale cambia la propria conformazione e libera i siti attivi dell’actina per la miosina nelle porzioni
laterali del disco A.
Quando il muscolo è rilasciato, la miosina non tocca l’actina; appena arriva il segnale, la miosina scinde l’ATP, in
quanto è ad attività ATP-atica, e lega il sito attivo dell’actina che si è reso evidente; poi ruota passando da una
posizione che è a 90° circa ad una più inclinata e quindi avvicina i filamenti sottili alla metà del disco A; se questo
viene fatto in sequenza tante volte, come il remo nell'acqua, si verifica un accorciamento del sarcomero fino a che
la porzione laterale, quindi i dischi I, scompaiono e scompare la zona H.
Si verifica dunque un incunearsi dei filamenti sottili nel disco A, spinti dalla forza della miosina, fino a che, a
contrazione completa, il disco I non si nota più e non si nota più neanche la diversità di colorazione al centro
della zona A.
L'arrivo di altro ATP scinde il complesso tra actina e miosina e rende possibili altre contrazioni. Se l' ATP non
ritorna, come ad esempio dopo la morte dell'organismo, i complessi actina-miosina diventano stabili per un certo
quantitativo di tempo ( il cosiddetto rigor mortis ).
Ciò che abbiamo detto finora altro non è che una sintesi di caratteristiche generali, ma i nostri muscoli non sono
fatti tutti dall'insieme di fibre uguali ma da un miscuglio di fibre un pò diverse tra loro, di cui i capi di questa
diversità si chiamano “ fibre rosse e fibre bianche”.
Al centro ( diciamo ) di questa diversità ci sono tante fibre con caratteristiche intermedie. I nostri muscoli sono,
dunque, un insieme di fibre in parte geneticamente determinate; con l'esercizio possiamo enfatizzare la grandezza
delle fibre, farle diventare ipertrofiche ma non ne aumentiamo il numero.
FIBRE ROSSE E FIBRE BIANCHE
Identifichiamo con il termine di fibre rosse, delle fibre muscolari ricche di una cromo proteina, la “ mioglobina”
che gli conferisce questo colore.
La mioglobina lega l'ossigeno, che si rende poi inattivo durante la contrazione.
Le fibre rosse sono lente nella contrazione ma poco affaticabili, facendo ottenere il tono muscolare.
Quindi i muscoli della nostra schiena sono in prevalenza fibre rosse.
Le fibre rosse sono ricche di mitocondri e sono a prevalente metabolismo ossidativo, il che vuol dire che hanno
tantissimi mitocondri che producono ATP.
Le fibre bianche, pallide perché hanno poca mioglobina, sono rapide nella contrazione ma facilmente affaticabili;
infatti entrano in gioco laddove i movimenti sono veloci.
Hanno un minor contenuto di mitocondri e per questo motivo sfruttano prevalentemente il metabolismo
glicolitico.
Quindi hanno anche grosse riserve di granuli di glicogeno mentre, le fibre rosse, caso mai, utilizzano riserve di
grasso.
Vista al microscopio ottico la fibra rossa presenta sotto il plasmalemma tanti mitocondri, i quali si trovano anche
tra le miofibrille che sono poche al centro della fibra.
La fibra bianca, invece, presenta pochi mitocondri, i quali non si trovano sotto il plasmalemma ma solo tra le
miofribille, che sono tante ed occupano la maggior parte del citoplama.
Questi elementi muscolari hanno possibiltà di ripresa dopo un danno?
Sono elementi stabili; possono, tuttavia essere soggetti ad una riparazione, se pur con possibilità modeste, da un
compartimento germinativo, detto “ cellule satelliti”, le quali devono questo nome al fatto che stanno in posizioni
satelliti, cioè appiccicate lungo la fibra, tra la fibra e la membrana basale .Le cellule satelliti hanno più che altro
un grosso significato nella ricerca perché si cerca di convincerle a differenziarsi verso una muscolatura che non ha
questo compartimento germinativo, come il cuore.
Questa possibilità modesta di riparo deve avvenire, però , a membrana basale integra.
Se si rompono le membrane basali entra il connettivo del perimisio e fa cicatrice connettivale, che ripristina
l'integrità ma non la contrazione e quindi quelle fibre non si contrarranno più in maniera corretta.
TESSUTO MUSCOLARE CARDIACO
È responsabile di una contrazione involontaria e costituisce il cosiddetto
miocardio.
Noi ci occuperemo del miocardio “comune”, quello, cioè, che contraendosi dà
origine al lavoro cardiaco. Gli elementi costitutivi della muscolatura striata
cardiaca sono cellule chiamate “miocardiociti” , strutture mononucleate ( hanno
al massimo due nuclei posti in posizione centrale ).
Il miocardiocita ha una struttura che ripete lo schema della fibra rossa; ha grosso
metabolismo aerobico, quindi molti mitocondri, che sono i più grossi mitocondri dell'organismo, disposti sia sotto
il plasmalemma che tra le miofibrille ( gli altri organuli si trovano nelle zone vicino il nucleo ).
Il miocardicita ha una forma a cilindretto, ma le estremità sono sfrangiate. Queste estremità vengono in contatto
con altre strutture simili, formando una specie di rete detta “plesso”.
Dobbiamo dunque presupporre che i miocardiociti siano legati tra loro, ma da che cosa?
Da giunzioni particolari che nel loro complesso prendono il nome di “ disco intercalare” o “ stria scalariforme”
perché sembra fatta come i gradini di una scala ( scala che si crea nel punto di giunzione tra due miocardiociti ).
La scaletta ha una porzione trasversale e lì i miocardiociti vicini sono legati da giunzioni aderenti, cioè giunzioni
che garantiscono l’ancoraggio di miocardiociti vicini che nella contrazione rimangono, così, solidali e legano sul
versante citoplasmatico le varie strutture cistoscheletriche.
La porzione longitudinale invece, quella parallela al maggior asse della struttura e quindi alle miofibrille e che
rappresenta l’alzata del gradino, è fatta da giunzioni comunicanti, punti di bassa resistenza elettrica che facilitano
il passaggio degli ioni responsabili della contrazione, che come impulso è nata nel miocardio specifico.
Quindi il disco intercalare è una struttura che non solo lega tra loro i miocardiociti a formare il plesso ma facilita
anche la trasmissione dell’impulso, attraverso le giunzioni comunicanti.
Ma il plesso presenta una sorta di buchi, allora dobbiamo ipotizzare che i miocardiociti siano circondati da che
cosa? Da una membrana basale che si continua con un fibrillare lasso che riempie le maglie della rete. Cosa ci
andrà in questo fibrillare lasso?
L’innervazione che modula il ritmo cardiaco e la circolazione che garantisce il buon trofismo di questo tessuto: la
circolazione coronarica.
Qualunque disturbo in questa circolazione, per esempio un’ischemia, provoca un danno al miocardio, con morte
dei miocardiociti ( il cosiddetto infarto ).Poiché non esiste un compartimento germinativo, quindi possibilità di
riparo , se la lesione del miocardiocita è compatibile ancora con una funzionalità cardiaca , ci si forma una
cicatrice , altrimenti la prognosi è nefasta.
Dunque mentre la muscolatura scheletrica, se pur con regole molto strette e per danni piccoli, si ripara, il cuore
ancora no.
I miocardiociti atriali sono più sottili e danno origine ad una struttura più esile di quella ventricolare. Attorno a
ciascun miocardiocita c’è una struttura che non è altro che la rivelazione della membrana basale , che è una
specie di endomisio , che si continua, poi , negli spazi lasciati liberi dal plesso, in un fibrillare lasso.I
miocardiociti atriali hanno capacità endocrine ( questa capacità rientra nel cosiddetto fenomeno endocrino in
elementi non epiteliali ), infatti vicino il nucleo, dall’apparato di Golgi, emergono granuli electrondensi che sono
il fattore natriuretico atriale che viene riversato nel sangue e partecipa al fenomeno della regolazione della
pressione
Come nella muscolatura striata scheletrica il reticolo liscio di membrana forma una struttura chiamata triade, nel
cuore è presente invece una diade, stessa struttura, ma i tubuli T invece di dare origine a delle cisterne, formano
solamente una dilatazione che crea una struttura a due oggetti, la diade appunto.
MUSCOLATURA LISCIA
La muscolatura liscia è una muscolatura involontaria e deve il suo nome alla mancanza di miofibrille, in quanto
la sua struttura contrattile non le sintetizza, per cui non si vedono striature.
Quali sono gli elementi che la costituiscono?
L’elemento di base è la cellula muscolare liscia, che ha una forma detta a
“fuso” , che presenta una parte equatoriale più slargata mentre ai poli si
assottiglia. Nella parte slargata si trova il nucleo ai cui poli si trova il
citoplasma cha dà origine a due zone dette “ coni sarcoplasmatici” o “ calotte
polari” , zone in cui si trovano la maggior parte degli organuli, escluso il
reticolo liscio che, “come al solito” si trova un po’ ovunque.
Nella porzione periferica del citoplasma si trova la componete contrattile, la quale è costituita dal filamento di
actina ( actina propria della muscolatura liscia ).
Questi filamenti si legano da un lato alla membrana attraverso proteine di legame, le cosiddette “placche dense”,
poi vanno a finire dentro il citoplasma in una zona densa detta “corpo denso” in cui si raccordano con altri
filamenti e proseguono il loro percorso all’interno del citoplasma andandosi ad ancorare al lato opposto della
membrana.
Quindi , attraverso il meccanismo di placche dense che agganciano la membrana, corpi densi che stimolano delle
linee Z , che altro non sono che punti di raccordo tra filamenti sottili, si crea una rete di filamenti sottili che
avvolge tutta la porzione centrale della cellula occupandone la zone periferica. Sappiamo che per contrarsi è
necessario che l’actina interagisca con la miosina; qui manca il sarcomero perché la miosina è sotto una forma
molecolare detta “ inattiva” ( in quanto la coda si ripiega su se stessa e rende così le teste della miosina non attive
per il contatto con l’actina ).
Gli elementi intermedi, sotto forma di vari tipi di proteine , legano le strutture sottili alla membrana, le raccordano
al livello delle placche dense, seguono il decorso dei filamenti che sono a forma di una specie di scheletro rigido
che segue le maglie ( maglie, come prima, delimitate dai filamenti sottili ).
Se andiamo a vedere la superficie di queste cellule affusate noteremo che ogni tanto ci sono delle introflessioni
del tipo di vescicole di micropinocitosi, ma queste non si chiudono mai per entrare nella cellula, restano aperte e
sono ricche di una proteina, detta “caveolina”, da cui il nome caveole ( somigliano ai tubuli T della muscolatura
striata ; infatti le caveole sono in stretto contatto con il reticolo liscio e sembrano avere lo stesso ruolo, nella
gestione del calcio, che la triade o la diade hanno nella muscolatura striata ).
Quando le cellule muscolari lisce si aggregano a formare lamine, come a circondare, a formare una conca negli
organi cavi, si organizzano ponendo la parte slargata a contatto con quella più sottile di quelle vicine, formando
così una struttura detta a “ mosaico”.
Ciascuna cellula è circondata dalla membrana basale che si interrompe solo nei punti in cui le cellule muscolari
lisce vengono in contatto giunzionale.
Ma come si mettono in contatto giunzionale?
Attraverso due tipi di giunzioni:
Una è quella aderente, le altre sono giunzioni comunicanti, punti di bassa resistenza elettrica che avranno un ruolo
nella trasmissione dell’impulso nervoso per la contrazione. Poi esiste un interstizio molto sottile, connettivale, che
porterà vasi e nervi per il trofismo della muscolatura.
Anche qui c’è un arrivo di ioni calcio, al momento della contrazione, nel citoplasma.
Il calcio, legandosi ad una proteina, detta “calmodulina”, prima provoca la fosforilazione della miosina che da
quella famosa struttura ripiegata passa a quella a coda rilasciata, e quindi rende attivi i siti di legame; poi si ha il
legame con l’actina e appena actina e miosina si trovano in contatto, la miosina ruota, passando da 90° a 45° e
scoprendo i filamenti.
Questo termina in una contrazione che viene detta “ peristaltica” perché è ad onde lungo, per esempio, la parete
intestinale.
Poi va nella parete dei vasi, quindi è un tipo di contrazione che ha degli effetti meno forti rispetto alla
muscolatura striata anche se molto duraturi e ha la cellula liscia a riposo mentre quando si contrae la cellula non
solo diminuisce di lunghezza, aumentando di volume, ma ruota lievemente perché è tutta ingabbiata dentro il
citoplasma non solo da filamenti contrattili ma anche da quelli rigidi e quelli non possono scivolare;quindi la
cellula sembra un po’ tutta a porzioni rigonfiate come un asciugamano strizzato.
Il legame actina-miosina avviene come nella muscolatura striata, con il sito attivo; sull’actina, cioè, c’è un sito
attivo che viene riconosciuto dalla miosina, la quale scinde l’ATP.
La muscolatura degli organi cavi è data da lamine, con andamento delle cellule su piani o longitudinali rispetto
all’organo cavo o trasversali, anche se sembra che di fatto siano disposte a scala a chiocciola con passo diverso,
una molto piatta e quindi le cellule sembrano disposte su un piano, una molto rigida e quindi le cellule sembrano
disposte su un piano ortogonale; in questo modo si formano le pareti.
Per concludere:
La muscolatura liscia ha potenzialità di ripresa, perché nel suo contesto esiste un compartimento germinativo,
anche qui con delle regole abbastanza precise, dato da cellule mesenchimali, il quale sotto stimolo appropriato in
sede può dare origine ad un differenziamento ex-novo di cellule muscolari.
ISTOLOGIA X LEZIONE:
TESSUTO NERVOSO
Definizione in base alla struttura:
Tessuto caratterizzato dalla presenza di cellule specifiche chiamate neuroni, e da cellule di
accompagnamento a queste, chiamate nella loro unione cellule di nevroglia o gliari.
Il neurone è quindi l’elemento specifico di questo tessuto nervoso che, anatomicamente, ne
costituisce il sistema nervoso, che come sapete è diviso in 2 grosse trance: il SNC costituito dall’encefalo e dal
midollo spinale, e il SNP costituito da gangli, nervi e terminazioni nervose.
Prima affronteremo la struttura del neurone come unità cellulare specifica d questo tessuto, dopodichè lo caliamo
nella realtà anatomica e lo facciamo accompagnare dalle cellule gliari.
NEURONE: Elemento specializzato nel portare
avanti le funzioni tipiche del tessuto nervoso e
quindi del SN.
Le funzioni da esso svolte sono:
Eccitabilità
Conducibilità
Polarizzazione funzionale
L’eccitabilità, è l’espressione massima di una
funzionalità comune a tutta la materia vivente
(abbiamo detto che tutta la materia vivente è irritabile, cioè in grado di rispondere in maniera opportuna a uno
stimolo). L’irritabilità del neurone risponde a basse soglie di solidificazioni e alla base di questo fenomeno di
questo fenomeno ci sono quelle proteine intrinseche di membrana che svolgono trasporto attivo di ioni, qui la
pompa implicata è la pompa Na+K+ che porta all’esterno della cellula Na+ e all’interno K+, contro gradiente di
concentrazione. Questa pompa, quindi, è un esempio di trasporto sincrono di ioni diversi verso distretti opposti
della cellula; ma mentre trasporta 3 ioni Na+ all’esterno della cellula, trasporta 2 ioni K+ all’interno portando
così a una formazione ineguale di cariche sui 2 versanti della membrana che è alla base del cosiddetto potenziale
di riposo (negativo all’interno rispetto all’esterno).
Quando arrivo uno stimolo, anche di bassa soglia, si ha una rapida attivazione di queste pompe con conseguente
ingresso e uscita di questi ioni, questa volta secondo gradiente; così si capovolge questa differenza di potenziale e
si crea quello che va sotto il nome di potenziale di azione, cioè una corrente elettrica che nasce in un punto e viene
contagiata a punti continui vicini della membrana del neurone, fino alla sua ultima diramazione.
Quindi il neurone, oltre ad essere irritabile, è anche capace di condurre questo stimolo elettrico che corrisponde ad
una informazione; pertanto potremmo dire che il neurone è irritabile e capace di conducibilità (ha una velocità di
propagazione che va da 0.5 a 103 m/s).
Questo per quanto riguarda le prime 2 caratteristiche del neurone; la 3^, che è quella della polarizzazione, può
avere delle eccezioni, ma in generale, questa corrente elettrica che si forma a seguito di una eccitazione della
membrana del neurone ha generalmente una direzione che può essere: dai prolungamenti dendritici verso il corpo
cellulare e dal corpo cellulare versi la periferia; ha quindi 2 possibilità diverse di conduzione. Questo ci porta
immediatamente a prendere visione del fatto che il neurone è una cellula estremamente complessa: ha un corpo
cellulare chiamato anche soma o pirenoforo, ha prolungamenti dendritici detti anche dendriti (che possono essere
da uno a +) ed è provvisto di un solo prolungamento chiamato massonico o assone o neurite o, potete aggiungere,
cilindrasse.
Quindi il neurone ci appare come una cellula caratterizzata sa un corpo cellulare, 2 tipi di prolungamenti: i
dendriti (da 1 a +) e l’assone (che è unico); la direzione di propagazione è verso il corpo cellulare nei dendriti (e
viene detta cellulipeta) mentre sfugge dal corpo cellulare lungo l’assone fino alla
periferia (ed è quindi cellulifuga). Come notate l’assone o neurita cilindrasse,
prende origine dal corpo cellulare in una zona particolare fatta ad imbuto che
prende il nome di cono di emergenza.
STRUTTURA E PROPRIETA’ DEL NEURONE: Il neurone, cellula di
dimensioni variabili, è costituito da una porzione chiamata corpo cellulare che
contiene il nucleo e che appare estremamente poco colorabile (quindi ricco di
eucromatina), la cromatina che esso trascrive prende il nome di nucleo da lavoro,
intendendo che questa cellula è implicata nella sintesi proteica.
Il nucleolo, appare molto sviluppato ed emerge molto colorato in seno ad un
nucleo che si colora pochissimo; le uniche zone di eucromatina sono facilmente
evidenziabili e una di queste viene studiata perché è la rappresentazione di una delle X eterocromatiche negli
individui di sesso femminile; ciò per dire che il neurone, così come il granulocita necrofilo, sono cellule in cui è
facile studiare questo ammasso di eterocromatina degli individui di sesso femminile.
Spostiamoci ora al citoplasma; si può notare che il colore depositato sul nucleo in questa cellula va a colorare il
citoplasma del corpo cellulare dando origine a quelle che si chiamano zolle di Niesl (??), N. è un ricercatore che
aveva messo a punto una colorazione specifica per i neuroni e aveva così scoperto che colorando il nucleo si
poteva anche colorare il citoplasma.
Che cosa c’è in questo citoplasma di così importante da potersi colorare con i coloranti nucleari? Tanti ribosomi,
sotto forma di polisomi liberi e ribosomi adesi alle cisterne del reticolo; quindi il corpo del neurone è ricco
dell’apparato proteosintetico, sia per quanto riguarda proteine strutturali di polisomi liberi, sia per quanto riguarda
riserve di membrana, lisosomi e proteine destinate a fuoriuscire dalla cellula. La sostanza di Niesl (???) si può
presentare sotto forma di zolle nei grossi neuroni perché lì c’è concentrazione diversa di reticolo e di polisomi che
danno l’immagine di un mantello di felino , da cui il nome di sostanza tigroide.
Quindi alla domanda che cos’è la sostanza tigroide, risponderete che sono cumuli di ribosomi liberi o adesi alle
cisterne del reticolo attraverso una semplice colorazione di microscopia ottica.
Il neurone, oltre ad avere tanti ribosomi in entrambe le forme, è anche una cellula che mostra un apparato di Golgi
molto sviluppato. Per meglio dire, sono tanti apparati di Golgi che si dispongono a formare una specie di sistema
circolare attorno al nucleo.
Anche i lisosomi sono molto sviluppati e ciò ci porta ad ammettere che questi neuroni effettuano un consistente
riciclaggio di sostanze e in particolare, il riciclaggio di membrana, può dare origine a corpi residui che poco
frequentemente vanno ad occupare il citoplasma dei neuroni (in soggetti specialmente anziani).
Il neurone è caratterizzato anche da tanti mitocondri di piccole dimensioni ma altamente sviluppati che porta il
neurone ad avere tanta energia per portare aventi le sue funzioni.
Anche il Reticolo Liscio sembra avere un grosso sviluppo e le cisterne del reticolo tendono a distribuirsi vicino al
Reticolo Ruvido il quel ruolo nel neurone non è ancora ben chiaro ma essenzialmente sembri essere collegato con
il deposito di ioni calcio indispensabili per la trasmissione dell’impulso nervoso da una cellula all’altra. Inoltre, in
alcuni neuroni ci possono essere degli accumuli di neurosecreto, mi riferisco ai neuroni dell’ipotalamo che
esocitano ossitocina, in vicinanza di capillari sanguigni, e questi verranno portati in giro cercando le cellule target
(cellule bersaglio che presentano i recettori specifici).
Le cellule nervose, possono presentare inclusi, e in alcune localizzazioni del SNC (tipo il locus ceruleo) possono
presentare anche grani di melanina e quindi apparire pigmentati di scuro.
Per ora abbiamo visto una cellula che si presenta attiva; se noi facciamo una colorazione specifica possiamo
mettere in evidenza nel corpo cellulare delle strutture filamentose che possono avere un andamento a maglia nel
corpo cellulare, mentre possono disporsi parallelamente nei prolungamenti dendritici prendendo così il nome di
neurofibrille, che viste al M.E. altro non sono che gli elementi del citoscheletro del neurone e cioè microtubuli e
neurofilamenti.
Invece, i microfilamenti di actina, tendono a disporsi al di sotto del plasmalemma. Per riassumere, le neurofibrille
altro no sono che il citoscheletro del neurone.
I PROLUNGAMENTI DENDRITICI: I dendriti possono essre da uno a + e sono delle ramificazioni chiamate
anche ramificazioni protoplasmatiche; nelle porzioni + grosse, i dendriti, hanno la stessa struttura del corpo
cellulare e quindi presentano tutto il corteo di organuli o inclusi appena descritti. Man mano che si assottigliano,
l’apparato protosintetico tende a diminuire.
I dendriti sono i prolungamenti del neurone che si dipartono dal corpo cellulare e a breve distanza possono dare
dei rami collaterali che si dipartono dal ramo principale con angoli inferiori a 90°, quindi con angoli acuti.
Pertanto, i prolungamenti dendritici, si possono riconoscere nelle sezioni istologiche dal fatto che hanno un
citoplasma con organuli uguali al corpo cellulare e dal fatto che le ramificazioni partono dal ramo principale con
angoli inferiori a 90°.
Le ramificazioni dendritiche, presentano delle strutture a pera dette spine dendritiche, che dimostrano gli enormi
contatti che si formano tra i neuroni nell’economia della trasmissione degli impulsi nervosi da un neurone
all’altro.
ASSONE: E’ sempre unico, si diparte dal corpo cellulare in una zona ad imbuto chiamata cono di emergenza che
non risulta colorata e quindi se ne deduce che è una parte priva di ribosomi. A partire dal cono di emergenza, e giù
per tutto l’assone, l’apparato protosintetico no c’è, si interrompe in una zona del corpo cellulare.
Nell’assone ritroviamo:
Ribosomi
Vescicole golgiane
Elementi del citoscheletro paralleli tra loro
L’assone, a differenza dei prolungamenti dendritici, si ramifica a distanza dal corpo cellulare e dà ramificazioni ad
angolo retto.
IL TRASPORTO ASSONICO: Movimento veloce lungo l’assone di molecole, organuli o vescicole. Questo
trasporto veloce può andare dal corpo cellulare di assone alla periferia (trasporto anterogrado) o può avere un
andamento inverso, dal terminale massonico al corpo cellulare (trasporto retrogrado).
In senso anterogrado, vengono trasportati enzimi, riserve di membrana e neurosecreti; in senso retrogrado può
tornare al corpo cellulare tutto ciò che dev’ essere digerito e tutto ciò che l’assone capta per endocitosi.
Come si attiva questo trasporto? Attraverso gli elementi del citoscheletro, principalmente i neurotubuli disposti
parallelamente tra di loro.
Qual è la forza propulsiva? Sono delle proteine dette motrici che avendo l’attività ATP-asica scindono l’ATP,
legano queste strutture da trasportare, si legano a loro volta ai costituenti del citoscheletro e come delle
locomotive agganciano questi trenini e li trasportano lungo questi binari dati dal citoscheletro.
Il trasporto anterogrado si avvale della cinesina, come molecola trasportatrice, mentre quello retrogrado si avvale
della ghineina, proteina già incontrata nelle ciglia.
Questo era il trasporto veloce, la lungo l’assone esiste anche un trasporto detto lento: è un flusso unidirezionale
dal corpo alla periferia che porta alla progressione di ricambi dei costituenti della matrice del citoplasma e delle
proteine del citoscheletro.
Torniamo a parlare del neurone in generale. Il neurone, porta avanti la sua funzione integrandosi con altri, ma allo
stesso tempo è anche indipendente dagli altri. Dal punto di vista del rinnovo è una cellula perenne, non capace di
rigenerarsi.
C’è da dire, che la plasticità neuronale è capace di ripristinare entro certi limita la funzione del neurone in quanto,
durante la nostra crescita, si verifica in noi lo sviluppo del SN attraverso i contatti che questi neuroni sviluppano
tra di loro; quindi man mano che i neuroni muoiono la plasticità ne’urinale cambia. Quando, però, per trauma o
malattia perdiamo un n. consistente di neuroni, si forma di nuovo la continuità tra questi grazie alle cellule di
nevroglia, ma non se ne ripristina la funzione.
I neuroni vengono classificati in base a:
N. di prolungamenti
Lunghezza dell’assone
Loro ruolo funzionale
In base al n. di prolungamenti dendritici abbiamo:
Neuroni unipolari con unico prolungamento che è l’assone ( es. coni e bastoncelli).
Neuroni dipolari con 2 prolungamenti: 1 dendrita e un assone. In questo caso i 2 prolungamenti
emergono dai poli opposti della cellula (es. i gangli).
Neuroni pseudounipolari o a T nei quali il corpo cellulare durante lo sviluppo emette 1 prolungamento
che a breve distanza dal corpo cellulare si ramifica e va in direzione opposta dando alla cellula l’immagine di una
T. Questi neuroni presentano + dendriti dando al corpo cellulare 1 aspetto stellato e definiscono quelli che si
chiamano neuroni multipolari.
Abbiamo detto che il neurone funziona in modo indipendente ma si integra con altri a formare strutture chiamate
sinapsi, ossia zone di continuità tra neuroni a livello delle quali si realizza la trasmissione dell’impulso nervoso.
Attraverso la situazione che si registra nel punto di contiguità tra neuroni, le possiamo distinguere in sinapsi
elettriche e chimiche.
La struttura delle sinapsi elettriche è di 2 neuroni che vengono in contatto e tra le 2 membrane si formano
giunzioni comunicanti, ossia punti a bassa resistenza elettrica. Le s. elettriche sono in numero estremamente
ridotto e caratterizzano solo alcune zone del SNC, nelle altre situazioni si parla solo di sinapsi chimiche che
prevedono la presenza di un neurotrasmettitore che propaga l’impulso nervosa da un neurone all’altro.
A livello delle sinapsi si ha: l’elemento che precede chiamato pre-sinaptico; l’elemento che segue chiamato postsinaptico e tra i 2 elementi uno spazio chiamato fessura o spazio sinaptico.
Al terminale massonico dell’elemento pre-sinaptico si va a concentrare il neurotrasmettitore sotto forma di
vescicole golgiane a contenuto diverso a seconda del tipo di sinapsi e delle zone interessate.
Ci interessa sapere quali parti del neurone vengono in contatto: noi abbiamo l’elemento pre-sinaptico che è il
neurone che ha già l’informazione e gli corre sulla membrana; il suo terminale assonico può venire in contatto o
con i dendriti (asso-dendritica) o col corpo (asso-somatica) o con un altro assone (asso-assonica).
Poi esistono in alcune porzioni del SNC contatti tra dendriti quindi sinapsi dendro-dendritiche.
Abbiamo finito di descrivere il neurone, l’elemento caratterizzante del tessuto nervoso.
Il tessuto nervoso, però, è caratterizzato anche da cellule di nevroglia, una famiglia di popolazioni cellulari che nel
loro insieme portano portano avanti tutte le funzioni che in altri distretti spettano al connettivo.
Mi spiego meglio: il SNC non ha stroma connettivale, ha solo le membrane basali; tutte le funzioni che aveva il
connettivo vengono assunte dalle cellule nevroglia del SNP. Dove il connettivo va a formare lo stroma ci sono
cellule di nevroglia ad accompagnare in tutte le sue parti il neurone.
Nel SNC le cellule di nevroglia portano avanti come si è detto, tutte le funzioni che in altri distretti vengono
assolte dal connettivo e cioè: funzioni di supporto meccanico, f. di riparo, f. trofica e f. di difesa.
Le cellule di nevroglia del SNC sono:
Gli oligodendrociti
Gli astrociti
Le cellule ependimali
La microglia (???)
- Gli oligidendrociti sono cellule dotate di prolungamenti che possono trovarsi nella sostanza grigia e nella
sostanza bianca. Nella sost. Grigia sono associati ai corpi cellulari dei neuroni e per questo si chiamano
perineuronali. Nella sost. bianca sono coloro che si incaricano di produrla e vengono detti oligodendrociti
interfascicolari.
Un oligodendrocita può produrre mielina e formare un tratto internodale a + assoni. Lungo il decorso dell’assone
ci saranno + oligodendrociti in fila a formare + segmenti internodali fino a che questo assone non fuoriesce dai
SNC.
Tra un segmento internodale e un altro abbiamo una zona chiamata nodo di Ranvier che indica i tratto di assone
nudo di centri, in cui termina l’avvolgimento dato da un oligodendrocita e ne comincia un altro.
- Gli astrociti sono una popolazione cellulare fatta a stella, cioè con dei prolungamenti connessi tra loro mediante
giunzioni comunicanti. Essi hanno, quindi, una funzione meccanica e strutturale tra i neuroni, ma soprattutto
partecipano alla costituzione della barriera ematoencefalica.
Gli astrociti possono trovarsi sia nella sost. bianca che nella sost. grigia ed hanno il ruolo di formare delle
espansioni terminali a ventosa a mò di piedino succhiatore che va a circondare, mettendosi in contatto con
estorciti vicini, l’endotelio che forma la parete dei vasi capillari.
Si realizza così una specie di barriera con l’endotelio (con giunzioni occludenti, che media attraverso la pinocitosi
tutti gli scambi), con i piedi succhiatori degli astrociti e con la membrana basale; è una barriera che filtra ciò che
passa dal torrente sanguigno al medium in cui si trovano i neuroni, realizzando così una fine regolazione delle sue
concentrazioni e anche della qualità. La barriera si realizza non solo nei confronti dell’endotelio, q quindi dei vasi,
ma anche nei confr del liquido cefalorachidiano contenuto nei ventricoli, nel canale midollare e nelle meningi.
Quindi il medium dove si trovano i neuroni è altamente controllato da questa barriera, che però può essere anche
problematica per quanto riguarda la somministrazione di farmaci che vengono riconosciuto come sostanze
estranee e, quindi, non vengono assimilati dall’organismo.
LEZIONE XI ISTOLOGIA
Continuando la trattazione delle cellule di nevroglia del SNC iniziamo a descrivere le cellule ependimali. Sono
cellule che hanno un aggregazione di tipo epiteliale. Già parlando degli epiteli di rivestimento avevamo detto che
erano cellule riferite ad altri contesti come una aggregazione di tipo epiteliale. Vanno a costituire delle membrane
che rivestono le cavità interne del SNC quali il canale centrale del midollo e i ventricoli cerebrali, cioè tutte quelle
zone in cui è contenuto il liquor.
Le cellule ependimali hanno una specie di microvilli, cioè delle estroflessioni molto lunghe che hanno una
struttura microscolare (?) ed hanno come significato quello di assorbimento. Queste cellule si dispongono in
assetto circolare formando tra di loro giunzioni. La parte sottostante si incunea nella membrana basale, a ridosso
della membrana basale troviamo espansioni terminali degli ….etrociti (?) a formare pedicelli.
L’ultimo appartenente alle cellule di nevroglia : microglia.
Sono cellule che appartengono al sistema dei macrofagi tanto che riconoscono il monocita come cellula
progenitrice e svolgono la loro funzione attraverso la fagocitosi.
Si trovano sia in posizione peribasale a controllare l’ingresso di eventuali sostanza estranee che provengono dal
circolo sanguigno. Si ritrovano anche vicino ai neuroni.
Oltre ai neuroni abbiamo anche altre cellule che nel loro complesso portano avanti funzioni che in altri distretti
esplica lo stroma connettivale:
▪Oligodendrociti
▪Astrociti
▪Cellule ependimali
▪Microglia
Passiamo ora al SNP dove dobbiamo prendere in considerazione tre strutture anatomiche:
▪Gangli
▪Nervi
▪Terminazione nervose periferiche
Gangli
Associazioni di cellule nervose in periferia. Quale è la struttura del ganglio?sono un insieme di neuroni dislocati
in un tessuto periferico. Questa struttura è circondata da una capsula di natura connettivale fibrosa densa forma le
capsule di tutti gli organi, ed è il tipo di connettivo che esprime al massimo le funzioni meccaniche e di difesa.
All’interno della scatola abbiamo un tessuto fibrillare lasso molto vascolarizzato che forma lo stroma della
struttura.
I neuroni non hanno diretto contatto con lo stroma connettivale ma sono circondate, a livello del corpo, da cellule
di nevroglia periferiche che prendono il nome di cellule satelliti, dal fatto che sono poste perifericamente in
posizioni satellite. Quindi i corpi appiattiti delle cellule satelliti circondano il neurone e il ganglio e formano la
lamina basale che continua separando questa struttura dalla parte connettivale che fa da stroma.
Cellule satelliti: sono le prime cellule di nevroglia trattate del SNP, associate ai corpi cellulari dei neuroni
gangliari (cellule satellite le abbiamo trovare anche in un altro tessuto).
I gangli sono dislocati ai lati del rachide cervicale e formano i così detti gangli cerebrospinali, sono costituiti
come natura specifica da neuroni a T o pseudobipolari, cioè neuroni che hanno un solo prolungamento che a
distanza si divide in due, dal corpo cellulare, e prendono direzione opposta. Morfologicamente sono simili ma uno
ha significato di dendrite, cioè porta informazioni dalla periferia al corpo del neurone, l’altra branca entra nel
SNC prendendo rapporto con altri neuroni o risalendo le vie.
Il corpo del neurone è circondato dalle cellule satelliti che lo proteggono meccanicamente e mediano gli scambi
poi si continuano con una membrana basale prodotta in parte anche da cellule del connettivo. Il prolungamento
sarà protetto da cellule di nevroglia di tipo diverso.
Affrontiamo ora l’argomento delle fibre nervose.
Le fibre nervose vengono definite come insieme dell’assone e guaine, invece nel SNP le guaine sono prodotte da
altre cellule di nevroglia, chiamate cellule di Schwann. Il comportamento delle cellule di Schwann è diverso a
seconda che noi parliamo di fibre mieliniche (bianche) o di fibre amieliniche (grigie). In entrambi i casi sono
rivestite da una guaina da un numero di cellule di Schwann.
Ora noi prendiamo in considerazione il comportamento delle cellule di Schwann nel formare la fibra mielinica.
Vi ricordo che la mielina era prodotta nei centri dagli oligodendrociti, i quali per progressivo accrescimento di
lembi del citoplasma davano luogo ad avvolgimenti intorno a un elemento dell’assone formando un accollamento
di membrana che viene chiamato mielina.
In periferia la cellula di Schwann si comporta in maniera simile. Durante lo sviluppo abbiamo che la cellula di
Schwann si avvicina all’assone, forma una specie di goccia che tende a circondare l’assone, e forma una specie di
accollamento tra i lembi della goccia che prende il nome di mesassone. Per accrescimento di uno dei lembi del
mesassone si ha il progressivo avvolgimento dell’assone da parte della cellula di Schwann , che si avvolge attorno
scivolando tra una delle branche della goccia e l’assone stesso. Ad ogni giro il citoplasma viene riportato in
periferia attorno al nucleo. Quando la mielina è stata costruita di fatto altro non è che una serie di accollamenti
della membrana della cellula di Schwann con pochissimo citoplasma e un nucleo interposto, struttura di natura
lipoproteica con un mosaico fluido estremamente poco fluido che quindi impedisce molto gli scambi.
Si crea così la guaina che ha il significato di isolamento dell’assone da restante ambiente e quindi si crea un
rivestimento fatto da un insieme di manicotti ciascuno prodotto da una cellula di Schwann e chiamato segmento
internodale. Dove finisce il rivestimento di una cellula di Schwann e ne comincia un altro si chiama nodo di
Ranvier , esattamente come nei centri. Quindi le cellule di Schwann sono coinvolte nella formazione della guaina
mielinica del singolo assone .
Se vi ricordate nei centri un oligodendrocita poteva dare la guaina mielinica a più assoni contemporaneamente
per quello che era il segmento internodale (questa è la prima differenza). La seconda differenza è che nei centri il
nodo di Ranvier è nudo, cioè l’assone non presenta alcun rivestimento (teniamo presente che nei centri esiste una
barriera che tiene a controllo: barriera ematoencefalica). In periferia oltre ad essere diverso il rapporto numerico
tra cellula di Schwann e assone (1:1), il nodo è anche protetto da lembi del citoplasma della cellula di Schwann
che precede rispetto a quella che segue che si interdigitano, non formano un involucro isolante perché non danno
origine a mielina, la teoria saltatoria dell’impulso nervoso è ancora valida ma l’assone è protetto dalla presenza
del citoplasma delle cellule di Schwann interdigitate, inoltre all’esterno si viene a formare una membrana basale
poichè la cellula di Schwann è un grado di formare la lamina basale e il connettivo che fa da stroma provvederà a
formare la porzione reticolare.
Quindi la fibra mielinica che viene etichettata come l’assone e le guaine disposta perifericamente è così intuita:
partire dall’interno verso l’esterno, assone, rivestimento della cellula di Schwann che dà origine alla mielina per
un tratto internodale, la membrana basale che prende il nome di endometrio detto anche endonevrio.
Esattamente come succede nel muscolo.
Nel caso delle fibre amieliniche , cioè prive di guaina mielinica , le fibre così dette grigie una cellula di Schwann
dà origine a dei rivestimenti formando gocce nel proprio citoplasma nel quale accoglie più assoni , all’esterno
ancora la membrana basale chiamata endonevrio. Nel caso delle fibre amieliniche la cellula di Schwann si pone al
centro, non avvolge ma accoglie in gocce del proprio citoplasma + assoni , all’esterno ancora, l’endometrio.
Definiamo nervi un insieme di fasci di fibre nervose. Quindi un insieme di fibre amieliniche o meliche, come è
dettato da quel particolare nervo, si raccolgono in fasci, + fasci uniti tra di loro vanno a formare il nervo.
Il nervo è quindi un insieme di fibre nervose e guaine connettivali che procedendo dall’esterno verso l’interno si
chiamano:
● epinevrio che sta sopra, attorno, è un connettivo nella porzione periferica denso, perché deve
ammortizzare le forze che vengono portate sulla superficie di questa struttura, all’interno si fa
lasso o fibrillare lasso formando uno stroma connettivale che porterà i vasi all’interno del nervo.
● Perinevrio ricco di vasi
● endonevrio cioè la membrana basale, che circonda ciascuno strato di mielina
A complicare ulteriormente le cose ora interviene un’altra struttura. Come notate a circondare fasci di fibre
nervose, quello che noi chiamiamo il perinevrio che forma tutto lo stroma, a ridosso delle fibre sembra formare
delle bucce di cipolla. Quindi il perinevrio non è un semplice fibrillare lasso in quanto oltre ad avere cellule
chiamate fibroblasti e fibrociti, a questi livelli si arricchisce di cellule particolari che si chiamano cellule
perinevrali, cioè cellule del perinevrio.
A circondare ogni fibra nervosa c’è l’endonevrio che è una membrana basale poi è tutto perinevrio che è un
fibrillare lasso che fa da stroma e si insinua tra le fibre nervose, ma a circondare un fascetto si arricchisce di
cellule che hanno una disposizione particolare. Sono cellule estremamente appiattite che si dispongono attorno al
fascetto di fibre nervose costruendo una specie di involucro a bucce di cipolla creando tra di loro delle giunzioni
occludenti, cioè quelle giunzioni che fanno si che non possa passare niente tra una cellula e l’altra o quanto meno
deve essere tenuto a controllo, sono ricche di vescicole di micropinocitosi ad indicare un’attività di grosso
trasporto tra un versante e l’altro, sono dotate di lamina basale. Quindi noi avremo una struttura attorno ai fascetti
di fibre nervose data dalle cellule perinevrali, che ripeto presentano lamina basale, giunzioni occludenti, vescicole
di micropinocitosi. Dicevo involucro che media lo scambio che crea quindi un microambiente per le fibre
altamente selezionate.
Abbiamo visto il SNC nella sua struttura istologica, abbiamo visto strutture periferiche quali gangli e nervi che
sono gruppi di fibre nervose raccordate insieme.
Ora proviamo a pensare che queste strutture nervose hanno un loro target, cioè vanno a terminare dentro i tessuti o
strutture organizzate da tessuti.
Parliamo delle così dette terminazioni nervose, cioè quelle strutture che prendono raccordo con le strutture
periferiche (tessuti) e che possono avere come dimensione di trasmissione delle informazioni o dai centri della
periferia e quindi parliamo di terminazioni effettrici (cioè portano un’informazione sottoforma di segnale elettrico
che corre sulla membrana della fibra nervosa che verrà poi trasmessa all’elemento non nervoso) che possono
portare un’informazione alla motilità sia essa volontaria o no, oppure alla secrezione, formando quelle che sono le
fibre motrici o eccitosecretorie.
Poi abbiamo le così dette terminazioni nervose sensitive (afferenti) cioè che ricevono un’informazione dai tessuti
perifericamente e le portano ai centri. Questa sensibilità noi la tratteremo da un punto di vista della sensibilità
generale, cioè la sensibilità dolorifica, termica, tattile o di postura.
Le cellule che accompagnano sempre le terminazioni nervose prendono il nome di cellule di teloglia o nevroglia
terminale o in alcuni libri, Schwann modificate. Quindi dalla sua nascita a laddove terminano sono sempre
accompagnate da cellule di nevroglia.
Per le cellule effettrici ci occupiamo in particolar modo di quelle che portano informazioni alla motilità nella
fibra muscolare. Le fibre muscolari, cioè il tessuto muscolare striato di tipo scheletrico, ricevono innervazione
così detta multiunitaria, cioè neuroni disposti nelle corna anteriori del midollo spinale, neuroni motori, mandano
una fibra nervosa che da origine a più ramificazioni ciascuna delle quali prende rapporto con una fibra muscolare,
nel muscolo abbiamo così dei settori commisti ad altri che ricevono terminazioni nervose da un motoneurone e
altri da altri.
A livello delle singole fibre muscolari si crea quella che si chiama una giunzione mioneurale, cioè da un elemento
nervoso (una fibra) che porta l’informazione (che è una depolarizzazione) alla motilità e la fibra muscolare. Nel
punto di vicinanza spaziale si crea un rapporto a tipo sinapsi. La fibra è coperta da mielina ma a livello di contatto
con la fibra muscolare si interrompe la mielina (ultimo nodo di Ranvier) e la fibra continua dando delle
ramificazioni terminali con sopra varie cellule di teloglia che coprono la terminazione nervosa (proteggendola
laddove è scoperta) dalla porzione che non guarda la fibra muscolare, il perinevrio ha fatto tutt’uno col perimisio,
l’endonevrio fa un tutt’uno con l’endomisio quindi le 2 strutture si legano tra di loro, la fibra nervosa e quella
muscolare, attraverso le guaine di rivestimento. Tra le ultime ramificazioni e la fibra muscolare si crea un
brevissimo spazio detto fessura sinaptica (primaria), dove le ramificazioni terminali prendono vicinanza stretta
con la superficie della fibra. La fibra muscolare a questo punto presenta un citoplasma in cui non si trovano
miofibrille, ma molti nuclei, mitocondri, perché è una struttura che riceverà informazione alla contrazione. Nel
punto allora terminale noi avremo che la superficie della fibra muscolare si innalza in tante tegoline (le fessure
sinaptiche secondarie) nel tentativo di amplificare la superficie di membrana della muscolatura a contatto con la
fibra nervosa che porta l’informazione alla motilità.
A livello della fessura sinaptica primaria, cioè quell’avvallamento che riceve la vicinanza della terminazione
nervosa, si alza in tante piegoline, le fessure sinaptiche secondarie, quindi la superficie di membrana che mostra il
contatto con la fibra nervosa è molto amplificata, lo spazio intersinaptico è riempito dalle 2 lamine basali.
Come avviene la trasmissione dell’impulso che ha una depolarizzazione di membrana che corre sulla fibra
nervosa? Quando arriva il terminale assonico crea una serie di eventi che scatenano il meccanismo della esocitosi
del neurotrasmettitore (NT) e in questo caso è l’acetilcolina. Il NT è rilasciato nello spazio intersinaptico,
l’acetilcolina trova i recettori sulla fibra muscolare, ci si lega, si scatena così la depolarizzazione di membrana
sulla fibra muscolare che è informazione alla contrazione.
A vari livelli della fibra muscolare la membrana si infila all’interno tra le miofibrille a formare tubuli T , quindi la
depolarizzazione che corre sulla membrana corre nei tubuli T, ma i tubuli T sono vicini alle membrane del
reticolo liscio, quindi questa informazione passa a queste membrane, si ha uno stop del pompaggio all’interno del
reticolo liscio, le pompe ferme permettono il dilagare del calcio nel citoplasma per fenomeni di diffusione, cioè
secondo gradiente di concentrazione, si cambia la configurazione della troponina che tira fuori la tropomiosina dal
mascheramento del sito attivo dell’actina , la miosina ci si attacca ed abbiamo la contrazione. Quindi una serie di
eventi che non deve essere un imparaticcio ma che dovrete avere chiari come meccanismo.
Quando passa, c’è lo stop alla informazione della contrazione, l’acetilcolina si stacca dal recettore e viene captata
da un enzima (acetilcolinaesterasi) che scinde le 2 parti (acetil dalla colina) e questa viene riciclata.
Quindi la muscolatura striata di tipo scheletrico presenta le proprie fibre muscolari parallele tra di loro e innervate
ciascuna attraverso una placca motrice che la giunzione citoneurale cioè il punto di contatto ravvicinato tra la
componente nervosa e quella muscolare.
La muscolatura liscia può avere un’innervazione di questo tipo anche se i neuroni che sono coinvolti non sono
neuroni volontari ma di contrazione involontaria. Questa innervazione multiunitaria permette un più stretto
controllo nervoso sulla contrazione delle cellule muscolari lisce e questo si ha nella parete dei vasi sangugni.
Nella, invece, muscolatura liscia viscerale noi abbiamo che la componente nervosa che può arrivare ad alcune
cellule muscolari lisce ma non a tutte, è poi coadiuvata nella contrazione da stimoli miogeni che possono essere di
natura ormonale oppure addirittura insorti spontaneamente a livello della muscolatura liscia e passare alle altre
cellule attraverso giunzioni comunicanti, tipo quello che accade nel cuore.
Se parliamo di terminazioni nervose sensitive, le possiamo classificare in base al tipo di tessuto a livelli del quale
queste terminazioni arrivano o partono perché in questo caso si parla di terminazioni afferenti, cioè che portano
informazioni dal tessuto ai centri. Queste possono essere nei tessuti epiteliali, nei tessuti connettivi, nei tessuti
muscolari.
Nei tessuti epiteliali si parla di terminazioni libere poiché le fibre nervose giungono in prossimità della membrana
basale, perdono la guaina mielinica, e nude senza cellule di teloglia corrono tra le cellule epiteliali che le
accolgono in gocce quindi qui esiste un rivestimento che è fornito da cellule epiteliali a contatto tra di loro.
Oppure possiamo avere terminazione che prendono contatto con cellule specifiche, vi ricordo il disco tattile di
Merkel, tra cellula epiteliale e terminazione nervosa si stabilisce un contatto sinaptico con un interazione del NT
da parte della cellula. Richiamo l’attenzione sul neuroepitelio (terminazioni associate a cellule
neuroepiteliali)dove le cellule di questo tipo di epitelio hanno stretto rapporto con terminazioni nervose
adiacenti(?).
Le terminazioni libere si ritrovano anche a livello dei connettivi dove però sono circondate dalle cellule di
teloglia, tanto che nei connettivi e nello stroma connettivale e del muscolare non esistono cellule a mutuo contatto
e quindi qui si forma un involucro delle cellule di teloglia. Ma a livello dei tessuti connettivi e muscolari noi
abbiamo anche le così dette terminazioni capsulate. Sono struttre in parte connettivali e in parte nervose che si
collocano nel connettivo propriamente detto o nel connettivo propriamente detto stromale dell’organizzazione
muscolare del muscolo striato.
Come sono fatte?innanzitutto hanno una capsula che è costituita da un insieme di lamelle di sostanza
intercellulare connettivale fatta da sostanza fondamentale anista e fibrilline collagene. Tra una lamella e l’altra ci
stanno i fibroblasti che tengono a controllo la struttura connettivale, ma più internamente alle lamelle ci sono le
cellule del perinevrio che hanno accompagnato la fibra nervosa fino al suo ingresso nel corpuscolo, ora
l’abbandonano e si dispongono tra le lamelle a formare il microambiente. Queste lamelle vanno a disegnare uno
spazio interno detto clava che è tutto foderato dall’endonevrio che correva internamente, qui c’è la fibra nervosa.
La fibra penetra nel corpuscolo dando origine a delle ramificazioni terminali
circondate dalle cellule di teloglia.
Questa è la struttura generale delle terminazioni nervose capsulate.
Le terminazione capsulate sono diverse come morfologia cioè possono avere
più o meno lamelle, una clava interna più grande o più piccola, le fibre nervose
che si ramificano in un certo modo o in un altro a seconda del contesto in cui
andranno a collocarsi.
Abbiamo recettori di questo tipo nel connettivo e recepiscono sensazioni tattili sia superficiali che profonde.
L’abbiamo nel tendine dove recepiscono il grado di contrazione attiva del muscolo quindi di stiramento del
tendine che è la struttura che lega il muscolo alle ossa oppure nei fusi neuromuscolari che registrano invece
l’attività tonica dei muscoli. E quindi la contrazione muscolare messa insieme può essere tenuta a controllo oltre
che dalle terminazioni efferenti anche da quelle afferenti per mezzo dei corpuscoli nel contesto del tendine e della
muscolatura striata.