")
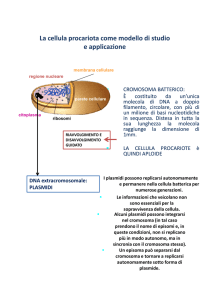
Cromosoma batterico
Plasmidi
Elementi genetici trasponibili
DNA fagico
NUCLEOIDE = unico cromosoma libero nel
citoplasma, dsDNA, circolare
superspiralizzato (lineare nei cromosomi
eucariotici), mancano gli istoni ma ci sono
proteine istone-like coinvolte nel packaging del
cromosoma e nella regolazione della
trascrizione.
DIMENSIONI:
~750 kbp in Mycoplasma spp.
~5000 kbp in Escherichia coli = 5
milioni di coppie di basi per una
lunghezza di 1,3 mm
Nell’uomo 23 coppie di
cromosomi per 2,9x109 paia di
basi pari a 990 mm
Mappa cromosomica dei geni di E. coli
Funzioni:
20% metabolismo
10% trasporto
10% regolazione/replicazione
5% strutturali
5% sintesi proteica
50% ancora sconosciute o
finalizzate a meccanismi di
resistenza e patogenicita’
Plasmidi
Elementi genetici
extracromosomici
Stabili, replicazione autonoma
(repliconi)
Sono episomi ma alcuni
possono integrarsi nel
cromosoma
Possono essere più copie per
cellula (1-50)
dsDNA circolare, da 1.5 a 400 Kb
Non codificano per funzioni vitali della
cellula e quindi non sono essenziali, ma
codificano per funzioni che possono
rappresentare un VANTAGGIO per la
cellula (scambio genico, resistenza,
virulenza, ecc)
Diffusi e ubiquitari fra Gram+ e Gram-,
sono un potente fattore di variabilità
genetica.
Forniscono le più disparate informazioni
genetiche dispensabili o indispensabili, a
volte stabili, a volte solo transitorie.
Sono un potenziale di informazione
flessibile e riarrangiabile.
Sono trasmissibili per trasformazione e
coniugazione e quindi acquisibili o
spendibili agilmente per attrezzare la
cellula ad adeguarsi ai fattori selettivi
dell’ambiente.
Plasmidi:
replicazione autonoma
La replicazione autonoma è indipendente dai tempi di
replicazione batterica.
Se il plasmide si duplica meno frequentemente del cromosoma
alcune cellule figlie possono non ricevere alcuna copia del
plasmide.
Se si duplica più frequentemente del cromosoma, le cellule
possono contenere copie multiple del plasmide.
Di alcuni plasmidi ne sono stati trovati anche 50 copie in
un'unica cellula; quindi rappresentano il veicolo ideale per la
produzione di proteine e fattori necessari in alte concentrazioni
(Es: resistenza antibiotici).
Coniugazione : Plasmide F formazione pili per trasferimento
genico (trasmissione orizzontale) attraverso un ponte
citoplasmatico.
Resistenza agli antibiotici: Plasmidi R
- degradazione enzimatica (e.g. penicillina)
- modificazioni enzimatiche (e.g. cloramfenicolo)
- alterata permeabilità (e.g. tetracicline)
- alterazione del target (e.g. streptomicine)
- via metabolica alternativa (e.g. sulfamidici)
Virulenza: fattori di invasione, produzione tossine e colicine (es:
Plasmide COL)
Metabolismo e Catabolismo: es. produzione di siderofori
Plasmide F
Il plasmide F è composto da 25 geni e regola la produzione dei pili F
di coniugazione.
Il trasferimento di un plasmide F conferisce alla cellula ricevente la
capacità di produrre pili F (cellula F+).
Il fattore F, come altri plasmidi, può essere autonomo ed
extracromosomico, oppure può integrarsi nel cromosoma batterico;
in tal caso diventa possibile il trasferimento di una porzione dello
stesso cromosoma dalla cellula a un’altra.
Plasmide R
Il plasmide R conferisce la resistenza agli antibiotici, è
caratterizzato dalla capacità di sintetizzare grandi quantità di
enzimi che distruggono il farmaco.
La resistenza dipende dalla sintesi di elevate concentrazioni di
enzimi specifici resa possibile dalla rapida duplicazione di copie
di questi geni.
I geni R possono essere trasferiti non solo da una cellula a
un’altra, ma anche da un plasmide R ad un altro (trasposoni).
Un solo plasmide può riunire fino a 10 geni per la resistenza
quindi rendendo la cellula resistente fino a 10 antibiotici diversi.
Tali geni possono anche essere trasferiti dai plasmidi al
cromosoma batterico, ai virus e a batteri di altra specie.
Plasmide COL
Sono plasmidi che codificano per le batteriocine o colicine.
La secrezione di colicine può essere messa in evidenza su piastra
esaminando le aree di lisi che si formano in una patina batterica
intorno a colonie produttrici di colicina.
Un batterio può essere immune ad una colicina se ne possiede il
plasmide relativo, attraverso la secrezione di una proteina che lega
la colicina.
Variazioni genetiche
nei
batteri
Modificazioni geniche
Mutazioni
Ricombinazioni
Meccanismi
di scambio genico
mutazioni
Ogni cambiamento nella sequenze di basi del DNA è denominato
MUTAZIONE.
TUTTE le mutazioni sono CASUALI. Alcune mutazioni sono instabili
(cioè frequentemente ritornano al loro stato di origine), altre non
incidono sull’organismo (silenti), altre infine sono stabili e causano
cambiamenti nelle caratteristiche dell’organismo.
Mutazioni puntiformi = piccoli cambiamenti (1 coppia di basi)
Riarrangiamenti = cambiamenti più grandi (duplicazione,
delezione, traslocazione, inversione)
Le mutazioni avvengono durante la replicazione del DNA
(frequenza di errore delle DNA polimerasi), oppure in seguito a
danno chimico (Es: deaminazione della citosina in seguito a
esposizione a raggi UV, specie reattive dell’ossigeno, ecc.→
mutageni)
ricombinazioni
Avvengono tra sequenze di DNA
omologhe.
L’ appaiamento di tali sequenze
può essere seguito dalla
formazione di crossing over tra le
regioni omologhe, dando come
risultato l’inserimento della
nuova sequenza in sostituzione
di quella originale.
Il fenomeno si può verificare ogni
volta che nel batterio è introdotto
un DNA esogeno omologo
(trasformazione, coniugazione,
trasduzione).
Segmenti di DNA mobili in
grado di traslocare nell’ambito
della stessa cellula.
Non capaci di replicazione
autonoma.
Trasposizione mediata da
ricombinazione sito-specifica
(enzima Trasposasi).
A.
Sequenze IS
B.
Trasposoni (Tn)
Scambio genetico
A differenza di molti eucarioti (riproduzione sessuata mediante
gameti), i procarioti non hanno la possibilità di variare il
patrimonio genetico ad ogni generazione.
Però possono scambiare piccole porzioni di DNA tra una cellula
(donatrice) a un’altra (ricevente).
Mentre negli eucarioti lo scambio genetico avviene con un unico
meccanismo (fusione dei gameti), nei procarioti questo può
avvenire con 3 MECCANISMI DIVERSI.
1.
TRASFORMAZIONE
2.
CONIUGAZIONE
3.
TRASDUZIONE
trasformazione
Processo attraverso cui i batteri acquisiscono frammenti di DNA
nudo e lo incorporano.
I batteri devono essere “competenti”: alcuni lo sono naturalmente
alla fine della fase log di crescita (Gram+ S. pneumoniae, Bacillus),
oppure sempre (Gram- H. influenzae, Neisseria), mentre altri devono
essere forzati (Es: trasformazione di E.coli con elettroporazione o
metodi chimici).
coniugazione
Trasferimento di DNA da
un batterio donatore (♂) ad
un batterio ricevente (♀)
attraverso il pilo sessuale
o pilo F.
Il ruolo (sesso) è
determinato dalla presenza
del plasmide F.
Può avvenire in quasi tutti
i batteri.
Cosa avviene nella coniugazione
Importante:
Il plasmide F non viene
trasferito come DNA a
doppia elica, ma a
singola elica.
Questo viene poi
duplicato nuovamente in
entrambe le cellule:
donatrice (F+) e
ricevente (F-).
Esiti della coniugazione
Caso 1
F+
x
F+
F+
F+
F+
F+
Caso 2
F+
x
F-
Hfr
(High Frequency
of
Recombination)
F+
Hfr
1. Quando il fattore F si integra nel cromosoma batterico
si forma una cellula Hfr. L’integrazione avviene in un sito
casuale nel cromosoma.
1.
2. La cellula Hfr è capace di iniziare la coniugazione
con una cellula F-.
3. Quando inizia il trasferimento del DNA, la cellula Hfr
cerca di trasferire TUTTO il suo cromosoma alla cellula F-. Il
primo DNA ad essere trasferito è quello cromosomico,
mentre l’ultimo è il fattore F.
4. Il trasferimento quasi mai è completo. I pili sono
strutture fragili, ed è frequente che si rompano prima che
l’intero cromosoma sia trasferito. Quindi il fattore F stesso
non è quasi mai trasferito, e la cellula ricevente resta F-.
Questa cellula però riceve nuovo DNA dalla cellula Hfr, e
questo DNA può ricombinarsi con alta frequenza con il
cromosoma ospite (Hfr= alta frequenza di ricombinazione).
La ricombinazione può risultare in eventi di conversione
genica, se il DNA trasferito e la corrispondente regione del
DNA ospite contengono alleli diversi dello stesso gene.
2.
3.
4.
La quantità di
geni trasferiti
dipende dal
TEMPO di
coniugazione.
trasduzione
Ricombinazione genetica attraverso un’infezione fagica.
Capside o
testa
Sistema
contrattile
Fibre della
coda
Coda
Piastra
basale
Due tipi di infezione
Ciclo litico: infezione con fagi virulenti e conseguente lisi cellulare
Ciclo lisogenico: infezione con fagi temperati ed integrazione del
genoma virale (profago) nel genoma batterico senza lisi cellulare
Il ciclo lisogeno può trasformarsi in ciclo litico con l’uscita del profago
dal genoma
N.B.
IMMUNITA’ FAGICA:
durante lo stato
lisogeno la cellula
batterica e’ immune da
infezioni di altri fagi
dello stesso tipo del
fago integrato
Infezione
litica
L’infezione di un batterio da parte di un fago virulento provoca
inevitabilmente la morte della cellula mediante lisi, con il rilascio di
nuove particelle fagiche.
Il ciclo litico dura circa 20 minuti.
Una cellula batterica infettata da un solo fago può rilasciare centinaia
di fagi progenie.
Infezione lisogenica
Nell’infezione di un
batterio da parte di
un fago temperato,
il DNA del fago si
integra nel
cromosoma
batterico. Il batterio
si moltiplica
indisturbato (batterio
lisogenizzato). A
volte il DNA
integrato può
excidersi dal
cromosoma e ridare
un ciclo litico.
I nuovi fagi possono portare, nel loro DNA, frammenti di cromosoma
batterico contenenti le sequenze vicine al sito di integrazione.
Generalizzata
(associata al
ciclo litico):
trasduzione
nella quale ogni
gene batterico
puo’ essere
potenzialmente
trasferito
Specializzata
(associata al
ciclo lisogeno):
trasduzione
nella quale
specifici geni
batterici sono
trasferiti
1. Infezione della cellula batterica (donatore)
2. Replicazione del fago e degradazione del DNA dell’ospite
3. Assemblaggio delle particelle fagiche
4. Rilascio dei fagi neoformati
5. Infezione di una nuova cellula batterica (ricevente)
6. Ricombinazione omologa
Trasduzione specializzata
1. Infezione della cellula batterica (donatore)
2. Integrazione del fago (lisogenia)
3. Riattivazione del DNA fagico, replicazione e rilascio dei nuovi fagi
4. Infezione di una nuova cellula batterica (ricevente)
5. Ricombinazione omologa
CONVERSIONE FAGICA
Batteri che assumono nuovi caratteri fenotipici quando
vengono lisogenizzati con un profago ricombinato
(contenenti geni di origine batterica).
ESEMPI DI CONVERSIONE FAGICA:
La trasformazione di batteri avirulenti in batteri produttori di
tossine, eg. Corynebacterium diphtheriae (fago beta con
proteina Tox)
Amplia il corredo di tossine batteriche, eg. Clostridium
botulinum (almeno due delle sette tossine botuliniche),
Streptococcus pyogenes (alcune tossine pirogene)
Modificazioni antigeniche che si riscontrano in alcune
salmonelle, shigelle ed altri batteri
Base genetica dei fattori di virulenza
")