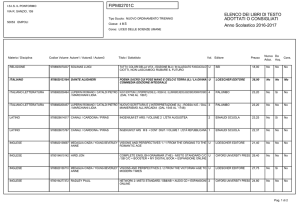
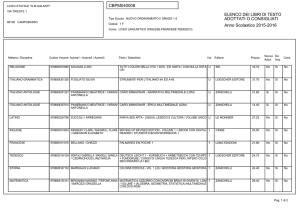
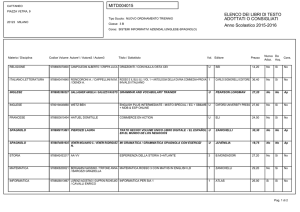
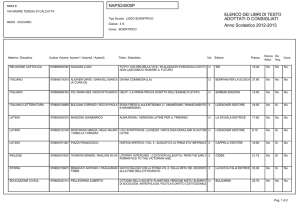
Lezioni di
biochimica
2
Lezione 5
Il metabolismo
dei carboidrati
3
© Zanichelli editore, 2014
La produzione di energia:
catabolismo dei nutrienti
La degradazione enzimatica delle sostanze nutritive
principali (carboidrati, lipidi e proteine) procede
attraverso una serie di reazioni che hanno lo scopo
ultimo di generare energia chimica.
Questa energia viene utilizzata per tutte le reazioni
cellulari, ivi comprese quelle anaboliche (biosintesi).
© Zanichelli editore, 2014
4
Il catabolismo dei carboidrati:
la glicolisi (I)
• La glicolisi è la via di degradazione del glucosio
(a 6 atomi di C) a piruvato (a 3 atomi di C).
• In condizioni anaerobie, il piruvato sarà ridotto
ad acido lattico (fermentazione lattica).
• In condizioni aerobie, il piruvato sarà usato per
generare acetil-CoA (a due atomi di C) che entrerà
nel ciclo di Krebs.
© Zanichelli editore 2014
5
Il catabolismo dei carboidrati:
la glicolisi (II)
© Zanichelli editore 2014
6
Le fasi della glicolisi
Le reazioni della glicolisi
sono organizzate in due
fasi principali:
1. La fase di preparazione,
in cui viene investita
l’energia di 2 ATP per
generare il
fruttosio 1,6-bisfosfato.
2. La fase di recupero,
in cui si generano due
piruvato e si producono
4 ATP.
© Zanichelli editore, 2014
7
La fase di preparazione
© Zanichelli editore, 2014
8
L’esochinasi
L’esochinasi è presente in tutte le cellule che fanno
la glicolisi.
È in grado di fosforilare molti esosi (come mannosio
e fruttosio) ed è inibita dal prodotto della reazione
(glucoso-6-P).
Quando la cellula ha soddisfatto il suo bisogno di
ATP, si accumula glucosio-6-P che inibisce la
glicolisi.
© Zanichelli editore, 2014
9
La glucochinasi
La glucochinasi è specifica per il glucosio ed è
presente nel fegato e non è inibita dal glucoso-6-P.
Questa differenza rispecchia i differenti metabolismi
cellulari: il fegato è il principale regolatore della
glicemia, quindi deve essere in grado di operare la
glicolisi in risposta non solo al fabbisogno energetico
ma anche alla concentrazione di glucosio del sangue.
© Zanichelli editore, 2014
10
La fase di recupero
Alla fine si generano quindi 4 ATP (due per ogni
gliceraldeide-3-P presente all’inizio) e 2 molecole
di piruvato.
© Zanichelli editore, 2014
11
La produzione di energia:
ossidazione (I)
Tutti gli organismi eterotrofi ricavano la loro energia da
reazioni di ossido-riduzione, in cui elettroni sono
trasferiti da un donatore (agente riducente) a un
accettore (agente ossidante).
Negli organismi aerobi, l’energia è ottenuta dalla
respirazione, in cui l’ossigeno è l’accettore finale degli
elettroni generati durante le reazioni di ossidazione del
catabolismo dei nutrienti.
© Zanichelli editore, 2014
12
Destino del piruvato in condizioni
anaerobie: fermentazione lattica
In condizioni anaerobie, la glicolisi procede riducendo
il piruvato a lattato (fermentazione lattica). Gli
elettroni necessari sono forniti dal NADH+H+
generatosi nella fase di rendimento.
© Zanichelli editore, 2014
13
Destino del piruvato in condizioni
anaerobie: fermentazione alcolica
In alcuni lieviti (per esempio il lievito di birra) avviene
invece la fermentazione alcolica, in cui il piruvato
è prima decarbossilato (perde 1 C) ad acetaldeide
(+ CO2) che viene poi ridotta (usando il NADH+H+)
a etanolo.
© Zanichelli editore, 2014
14
Destino del piruvato in condizioni
aerobie: l’acetil-CoA
In condizioni aerobie, il piruvato è importato all’interno
dei mitocondri. Qui, il complesso della piruvato
deidrogenasi opera una decarbossilazione, in cui il
piruvato a 3C viene trasformato in acetil-CoA (a 2C)
+ CO2. Gli elettroni generati sono ceduti al NAD+.
L’acetil-CoA così formato entra nel ciclo di Krebs.
© Zanichelli editore, 2014
15
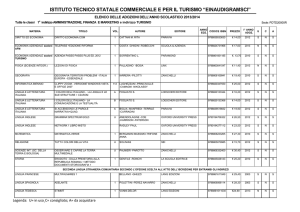
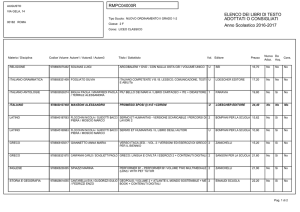
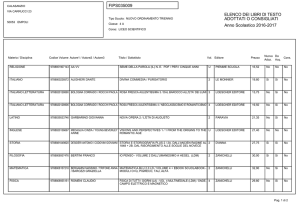
Considerazioni finali sul bilancio
energetico della glicolisi (I)
Nella glicolisi abbiamo visto il trasferimento di 2
fosfati da ATP a un esoso (glucosio e fruttosio) e di
due fosfati dall’acido fosforico a due triosi
(gliceraldeide-3-P).
Questi 4 fosfati vengono poi donati nuovamente all’ADP
per formare 4 ATP. Dato che due ATP erano stati
consumati, il bilancio netto è di 2 ATP.
© Zanichelli editore, 2014
16
Considerazioni finali sul bilancio
energetico della glicolisi (II)
Il passaggio chiave che consente il guadagno
energetico è l’utilizzo del fosfato per generare
l’1,3-bisfosfoglicerato.
La delucidazione di questo meccanismo ha fornito
la prima prova che l’energia derivata dall’ossidazione
di una molecola organica può essere conservata
sotto forma di ATP.
© Zanichelli editore, 2014
17
La gluconeogenesi riparte
dal piruvato
La biosintesi del
glucosio nel
fegato parte da
piruvato, che il
fegato ricava sia
dal lattato, sia
da molecole
come gli
amminoacidi.
© Zanichelli editore, 2014
18
La gluconeogenesi non è
l’inverso della glicolisi
La biosintesi del glucosio non è l’inverso della sua
degradazione, le due vie sono indipendenti.
Via glicolitica
© Zanichelli editore, 2014
Via gluconeogenetica
19
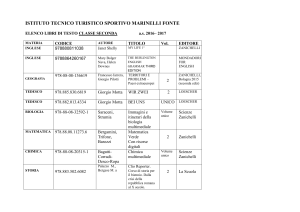
La glicogenosintesi
Il glucosio-6-P
può venire
utilizzato in
molte cellule
(fegato,
muscolo) per
sintetizzare il
glicogeno,
un’importante
riserva di
energia.
© Zanichelli editore, 2014
20
La demolizione del glicogeno
Il glicogeno può essere demolito generando unità di glucosio1-P, grazie alla glicogeno fosforilasi.
Queste sono poi trasformate in glucosio-6-fosfato.
© Zanichelli editore, 2014
21
La regolazione di glicolisi e
gluconeogenesi è speculare
La glicolisi è regolata a livello della
fosfofruttochinasi (da fruttosio 6-fosfato a fruttosio
1,6-bisfosfato) che è inibita da ATP e citrato e
attivata da AMP e ADP.
Specularmente, la reazione corrispondente nella
gluconeogenesi (da fruttosio 1,6-bisfosfato a fruttosio
6-fosfato) è regolata dalla fruttosio bisifosfatasi,
che viene attivata da ATP e citrato e inibita da AMP
e dal fruttosio 1,6-bisfosfato.
© Zanichelli editore, 2014
22