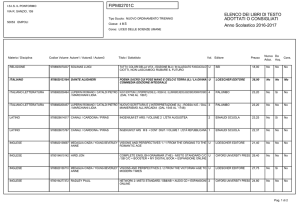
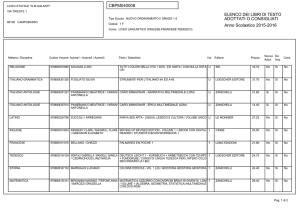
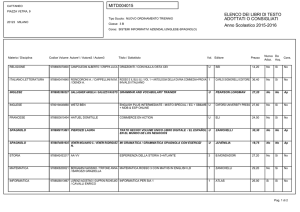
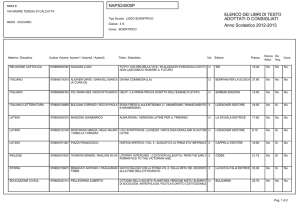
1
Il metabolismo
cellulare
2
© Zanichelli editore 2016
Che cos’è il metabolismo?
Gli organismi viventi trasformano le biomolecole che si
trovano negli alimenti per produrre energia e per ottenere i
precursori per la sintesi di macromolecole. Questo processo
di trasformazione si chiama metabolismo.
Le vie metaboliche possono essere cataboliche
(degradazione di molecole) o anaboliche (sintesi di
molecole).
3
© Zanichelli editore 2016
Il metabolismo
dei carboidrati
4
© Zanichelli editore 2016
Il catabolismo dei carboidrati:
la glicolisi
La glicolisi è la via catabolica che porta alla degradazione
del glucosio (6 atomi di C) in piruvato (3 atomi di C).
Condizioni aerobiche
Glucosio
Condizioni anaerobiche
Glucosio
Glicolisi
Piruvato
Fermentazione
Prodotti di fermentazione
Respirazione
CO2 + H2O
5
© Zanichelli editore 2016
Glicolisi: diverse fasi
La glicolisi si divide in due
fasi:
• fase preparatoria – 2
molecole di ATP sono usate
per produrre fruttosio 1,6bisfosfato;
• fase di recupero
energetico – produzione di
4 molecole di ATP e due
molecole di piruvato.
6
© Zanichelli editore 2016
Dal piruvato all’acetil-CoA
In condizioni aerobiche, il piruvato (3 C) è trasferito nei
mitocondri, dove il complesso dell’enzima piruvato
deidrogenasi lo trasforma in acetil-CoA.
piruvato + NAD+ + CoA-SH
acetil-CoA + CO2 + NADH + H+
7
© Zanichelli editore 2016
Il ciclo di Krebs
L’acetil-CoA entra nel ciclo di Krebs, o ciclo dell’acido
citrico, dove in una serie di reazioni la molecola è ossidata a
CO2, riducendo nello stesso tempo NAD a NADH.
8
© Zanichelli editore 2016
Il ciclo di Krebs: bilancio energetico
• L’ATP è prodotto indirettamente da GTP, generato quando
il succinil-CoA è idrolizzato a succinato.
• Il ciclo produce agenti riducenti: 3 NADH e 1 FADH2, che
possono essere usati per la sintesi di molecole di ATP.
9
© Zanichelli editore 2016
La catena di trasporto degli elettroni
NADH e FADH2 trasferiscono elettroni a un sistema di proteine
che si trovano sulla membrana interna dei mitocondri. Queste
proteine costituiscono la catena di trasporto degli elettroni o
catena respiratoria.
10
© Zanichelli editore 2016
La funzione della catena di trasporto
degli elettroni
La catena respiratoria è una sequenza di reazioni redox,
durante le quali le proteine che si trovano in diversi complessi
accettano elettroni e li donano subito al complesso
successivo.
L’ossigeno è l’accettore finale di elettroni.
La catena respiratoria non genera direttamente ATP, ma
produce energia sotto forma di forza proton-motrice, che
sarà necessaria per la fosforilazione ossidativa.
11
© Zanichelli editore 2016
La formazione di un gradiente
protonico
L’ossidazione di NADH libera 2 elettroni e 2 protoni (H+).
Gli elettroni entrano nella catena di trasporto, trasferiti dal
complesso I al complesso IV e all’ossigeno.
In ogni reazione redox, si
generano nuovi protoni
che vengono pompati
attraverso la membrana
mitocondriale nello
spazio intermembrana.
12
© Zanichelli editore 2016
Gradiente protonico e sintesi di ATP
Alte concentrazioni di H+ da un lato della membrana creano un
gradiente elettrochimico, per cui i protoni si riversano dallo
scomparto a concentrazione più alta nello scomparto a
concentrazione più bassa.
Il flusso di protoni passa
attraverso la regione F0 del
canale ATP sintasi.
Questa regione attiva la
subunità F1, dove ha luogo la
sintesi di ATP.
13
© Zanichelli editore 2016
Condizioni anaerobiche:
la fermentazione
In mancanza di ossigeno, il metabolismo dei carboidrati
non segue la via della respirazione cellulare: il piruvato
va incontro a fermentazione alcolica o fermentazione
lattica.
14
© Zanichelli editore 2016
La fermentazione lattica
Nella fermentazione lattica, il piruvato prodotto con la glicolisi
è ridotto ad acido lattico. Gli elettroni necessari sono forniti da
NADH + H+, generati durante la fase di recupero energetico.
Glucosio
Glicolisi
2 NAD+
2 piruvato
2 NADH + H+
2 acido lattico
15
© Zanichelli editore 2016
La fermentazione alcolica
In alcuni lieviti avviene la fermentazione alcolica: il piruvato
va incontro a una decarbossilazione ed è convertito ad
acetaldeide, poi ridotta a etanolo grazie a NADH + H+.
Glucosio
Glicolisi
2 NAD+
2 etanolo
2 piruvato
2 NADH + H+
2 CO2
2 acetaldeide
16
© Zanichelli editore 2016
Il metabolismo
dei lipidi
17
© Zanichelli editore 2016
Il metabolismo dei lipidi: dal cibo
o dalle riserve energetiche
I lipidi sono importanti fonti di energia, soprattutto nei muscoli
e nel fegato.
Possono essere introdotti nel corpo con l’alimentazione, o
possono essere presi dalle riserve energetiche delle cellule
adipose.
I lipidi presenti nei cibi (trigliceridi) sono assorbiti a livello
dell’intestino nel flusso sanguigno, dove si spostano associati
a lipoproteine, come le lipoproteine ad alta densità (HDL).
Gli acidi grassi che derivano dal tessuto adiposo sono
trasportati dalla proteina albumina.
18
© Zanichelli editore 2016
Il fegato e il metabolismo dei grassi
Il fegato sintetizza trigliceridi, che sono trasportati alle cellule
dalle proteine VLDL (very low density lipoproteins, proteine a
densità molto bassa). Dopo aver consegnato i trigliceridi,
diventano LDL, ricche in colesterolo. Sono assorbite dalle
cellule che hanno bisogno di colesterolo per le loro
membrane.
L’eccesso di colesterolo nel flusso sanguigno è rimosso dalle
proteine HDL (high density lipoproteins, lipoproteine ad alta
densità), che lo trasportano al fegato perché sia degradato.
19
© Zanichelli editore 2016
Gli acidi grassi nei mitocondri
Nei mitocondri avviene l’ossidazione degli acidi grassi.
Queste molecole sono trasportate nei mitocondri grazie al
trasportatore carnitina, con la formazione di acil-carnitina.
Nella matrice mitocondriale, la molecola reagisce con CoA
per rilasciare l’acido grasso, con la formazione di acil-CoA.
20
© Zanichelli editore 2016
Beta ossidazione degli acidi grassi
L’ossidazione degli acidi grassi è chiamata betaossidazione.
Durante il processo, le molecole di acidi grassi sono
degradate per generare acetil-CoA, FADH2 e NADH.
L’acetil-CoA può quindi entrare nel ciclo di Krebs.
21
© Zanichelli editore 2016
Il metabolismo
degli amminoacidi
22
© Zanichelli editore 2016
Il metabolismo degli amminoacidi
Gli amminoacidi rappresentano un contributo minore della
richiesta energetica.
In caso di digiuno prolungato, il corpo può distruggere le
proteine endogene (soprattutto nei muscoli scheletrici) per
ricavare una fonte di energia, ma il processo può essere
pericoloso per l’organismo.
Gli amminoacidi forniscono precursori essenziali per la sintesi
di molte molecole.
23
© Zanichelli editore 2016
La rimozione del gruppo amminico
La rimozione del gruppo amminico è una tappa importante
nel catabolismo degli amminoacidi.
Per la maggior parte degli amminoacidi, la rimozione avviene
in due reazioni:
•transaminazione;
•deaminazione ossidativa.
Il gruppo amminico è eliminato
sotto forma di catione ammonio
(NH4+).
24
© Zanichelli editore 2016
Transaminazione
La transaminazione avviene nel fegato, catalizzata dagli
enzimi chiamati transaminasi. Il gruppo amminico è trasferito
dall’amminoacido a un alfa-chetoacido (generalmente l’alfachetoglutarato), generando acido glutammico.
O
O
R1
R1
HO
HO
NH2
transaminasi
O
O
+
O
+
R2
R2
HO
HO
O
NH2
25
© Zanichelli editore 2016
La deaminazione ossidativa
L’acido glutammico generato con la transaminazione va
incontro a deidrogenazione grazie alla glutammato
deidrogenasi, che rigenera l’alfa-chetoglutarato e trasforma
il gruppo amminico in catione ammonio.
Gli elettroni sono trasferiti a NAD+ (o NADP+).
NAD(P)+
NAD(P)H + H+
alfa-chetoglutarato + NH4+
acido glutammico + H2O
glutammato
deidrogenasi
26
© Zanichelli editore 2016
Il catione ammonio
Il catione ammonio è tossico ed è eliminato dagli organismi
in forme diverse.
Nei mammiferi e negli anfibi, il catione ammonio è eliminato
con la sintesi di urea.
urea
27
© Zanichelli editore 2016