Lezioni di
biochimica
2
Lezione 6
Il metabolismo
di lipidi e proteine
3
© Zanichelli editore, 2014
Il metabolismo dei lipidi (I)
• I lipidi sono l’altra principale
fonte di energia delle cellule,
in particolare per tessuti
muscolari e fegato.
• Possono derivare dalla
dieta o essere mobilizzati
dalle riserve presenti nelle
cellule adipose.
© Zanichelli editore, 2014
Il pesce è un alimento ricco
di grassi insaturi ω3.
4
Il metabolismo dei lipidi (II)
• I lipidi alimentari (trigliceridi) assorbiti
dall’intestino viaggiano nel sangue legati a proteine
(lipoproteine) che li distribuiscono ai tessuti.
• Gli acidi grassi mobilizzati dal tessuto adiposo,
invece, vengono trasportati dalla proteina
albumina.
© Zanichelli editore, 2014
5
Il metabolismo dei lipidi (II)
Il fegato sintetizza trigliceridi che sono veicolati alle
cellule dalle proteine VLDL (Very Low Density
Lipoprotein).
Ceduto il loro carico, esse diventano LDL (Low
Density), ricche in colesterolo, e sono internalizzate
dalle cellule che necessitano di colesterolo per la
sintesi delle membrane.
L’eccesso di colesterolo nel sangue, invece, viene
rimosso dalle HDL (High Density Lipoproteins) che lo
trasportano al fegato per la sua eliminazione.
© Zanichelli editore, 2014
6
Il metabolismo dei lipidi (III)
© Zanichelli editore, 2014
7
L’ingresso degli acidi grassi
nel mitocondrio (I)
L’ossidazione degli acidi grassi avviene nel
mitocondrio. Per arrivarci, gli acidi grassi devono
superare 3 passaggi:
1. Esterificazione con Coenzima A a formare l’acilCoA (nel citosol) con consumo di 1 ATP.
2. Cessione dell’acido grasso alla carnitina con
liberazione del Coenzima A.
© Zanichelli editore, 2014
8
L’ingresso degli acidi grassi
nel mitocondrio (II)
3. Ingresso del complesso acil-carnitina nel
mitocondrio attraverso un trasportatore
specializzato e cessione dell’acido grasso
al Coenzima mitocondriale generando un nuovo
acil-CoA e carnitina libera.
© Zanichelli editore, 2014
9
La b-ossidazione (I)
L’ossidazione degli acidi grassi è detta b-ossidazione
perché nella prima reazione due idrogeni sono
strappati al legame tra i carboni 2 e 3 dell’acil-CoA
(detti carboni a e b) con formazione di FADH2 (che
entrerà nella catena respiratoria).
Il ciclo di reazioni successive porta alla generazione di
un acetil-CoA e di un acil-CoA con 2 atomi di
carbonio in meno.
© Zanichelli editore, 2014
10
La b-ossidazione (II)
A questo punto l’acil-CoA così generato re-inizia un
nuovo ciclo. La reazione procede quindi eliminando 2
C ad ogni ciclo e generando un Acetil CoA, fino ad
esaurimento della catena dell’acido grasso.
L’acido palmitico (16 C), genera quindi 8 Acetil CoA
a 2 atomi di C.
Per ogni ciclo si generano 1 FADH2 e 1 NADH e
viene utilizzato un Coenzima A.
© Zanichelli editore, 2014
11
La b-ossidazione
La β-ossidazione
dell’acido palmitico
© Zanichelli editore, 2014
12
La biosintesi degli acidi grassi:
l’acido palmitico (I)
• Gli acidi grassi cellulari sono sintetizzati da una via
completamente diversa dalla b-ossidazione.
• La biosintesi avviene nel citosol ad opera dei 7
enzimi del complesso della acido grasso sintasi.
• La biosintesi inizia con una molecola di acetil-CoA
che funge da iniziatore.
• A questa vengono via via condensate unità a 2 C
di malonil-CoA.
© Zanichelli editore, 2014
13
La biosintesi degli acidi grassi:
l’acido palmitico (II)
Quindi, per sintetizzare un
acido palmitico (16 C)
serviranno 1 acetil-CoA e 7
malonil CoA. Serve anche
potere riducente, fornito da
NADPH+H+.
© Zanichelli editore, 2014
14
La biosintesi degli acidi grassi:
allungamento (I)
L’acido palmitico è il precursore degli altri acidi
grassi a catena lunga. L’allungamento avviene sia
nel mitocondrio (per acidi grassi saturi e insaturi)
sia nel reticolo endoplasmatico (per acidi grassi
monoinsaturi come palmitoleato e oleato).
© Zanichelli editore, 2014
15
La biosintesi degli acidi grassi:
allungamento (II)
© Zanichelli editore, 2014
16
La biosintesi del colesterolo
• Il colesterolo è un componente essenziale delle
membrane cellulari.
• La sua biosintesi, che avviene nel fegato, è molto
complessa e richiede la presenza acetil-CoA e
NADPH + H+ nel citosol.
• Il colesterolo viene assunto anche con
l’alimentazione, e non esistendo una via catabolica a
sua carico bisogna fare attenzione a un suo eccessivo
accumulo.
© Zanichelli editore, 2014
17
Il metabolismo
degli amminoacidi (I)
• Gli amminoacidi, assunti dalle proteine della dieta,
contribuiscono in misura minoritaria al fabbisogno
energetico.
• In caso di digiuno prolungato, la demolizione delle
proteine endogene (soprattutto del muscolo
scheletrico) può diventare una fonte di riserva
importante, anche se pericolosa per l’organismo
(in quanto porta al deperimento e alla morte).
© Zanichelli editore, 2014
18
Il metabolismo
degli amminoacidi (II)
• Gli amminoacidi,
tuttavia,
forniscono
precursori
essenziali per la
biosintesi di molte
macromolecole
dell’organismo.
© Zanichelli editore, 2014
19
Rimozione del gruppo amminico
Uno degli aspetti più importanti del catabolismo
degli amminoacidi è la rimozione del loro gruppo
amminico.
Per la maggior parte degli amminoacidi questo
avviene tramite due reazioni: la transaminazione e
la deaminazione ossidativa.
Al termine il gruppo amminico è eliminato sotto
forma di ione ammonio.
© Zanichelli editore, 2014
20
Transaminazione (I)
In questa reazione, catalizzata nel fegato dalle
transaminasi (citosoliche e mitocondriali), il gruppo
amminico di un amminoacido viene trasferito a un
accettore a-chetoacido (solitamente a-chetoglutarato),
generando un amminoacido (acido glutamico) e un
a-cheotacido diverso a seconda dell’amminoacido
di partenza.
© Zanichelli editore, 2014
21
Transaminazione (II)
Le transaminasi utilizzano il cofattore piridossal
fosfato (PLP), derivato dalla vitamina B6.
© Zanichelli editore, 2014
22
Deaminazione ossidativa
L’acido glutammico prodotto dalla transaminazione
viene deidrogenato nel fegato dalla glutammato
deidrogenasi che rigenera l’a-chetoglutarato e
trasforma il gruppo amminico in ione ammonio. Gli
elettroni sono ceduti al preferenzialmente al NAD+
(ma anche al NADP+).
© Zanichelli editore, 2014
23
Transaminazione e deaminazione
ossidativa sono accoppiate
© Zanichelli editore, 2014
24
Lo ione ammonio (I)
Lo ione ammonio è tossico ed è eliminato in forme
diverse nei diversi organismi.
© Zanichelli editore, 2014
25
Lo ione ammonio (II)
Nei mammiferi e negli anfibi lo ione ammonio è
eliminato attraverso la biosintesi dell’urea (ciclo
dell’urea), secondo la reazione complessiva:
Struttura dell’urea
© Zanichelli editore, 2014
26
Metabolismo degli amminoacidi
e salute
Difetti ereditari nel metabolismo degli amminoacidi
sono correlati a numerose patologie nell’uomo.
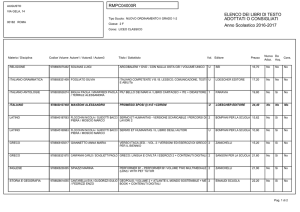
Malattia genetica
Causa
Albinismo
Difettoso metabolismo della tirosina
Cistinuria
Difettoso trasporto della cisteina
Fenilchetonuria
Difettoso metabolismo della fenilalanina
Malattia delle urine a sciroppo d’acero
Difettoso metabolismo degli a-chetoacidi
© Zanichelli editore, 2014
27