IL CICLO CELLULARE
Mitosi e meiosi
Il ciclo cellulare comprende tutti quegli stadi attraverso i quali passa una cellula
da una divisione all’altra, di solito questo meccanismo può durare dalle 8 alle 20
ore.
Si compone sostanzialmente in due fasi:
INTERFASE: in interfase la cellula non si sta riproducendo, ma è
estremamente attiva perché sta sintetizzando lipidi, proteine
e macromolecole, questo è il periodo che intercorre tra una
fase M e la successiva
FASE M: in questa fase si ha la vera e propria mitosi cellulare e il
processo di citocinesi.
Inoltre in INTERFASE si possono riconoscere ulteriori tre stadi:
G1: (GAP 1) questo è un intervallo fra il completamento della mitosi e
l’inizio della sintesi di DNA. La crescita ed il normale metabolismo
cellulare avvengono in fase G1. Durante questo periodo la cellula
controlla l’ambiente esterno e le proprie dimensioni e, al momento
opportuno, avvia la fase decisiva di replicazione del DNA,
incrementando l’attività degli enzimi richiesti per la sintesi del DNA.
S: (SINTESI DEL DNA) durante la fase S avviene la replicazione del
DNA e la sintesi degli istoni poiché la cellula deve duplicare i propri
cromosomi.
G2: (GAP 2) la fase G2 fornisce un intervallo di sicurezza permettendo
alla cellula di assicurarsi che la duplicazione del DNA sia stata
completata prima di iniziare la mitosi. Inoltre durante questo
intervallo aumenta la sintesi proteica.
La durata del ciclo cellulare varia molto, soprattutto della fare G1, in una cellula
adulta può durare dalle 12 alle 36 ore, la differenza della durata del ciclo
cellulare, quindi, si trova proprio in G1.
Infatti, alcune cellule, come quelle nervose, si arrestano nella fase G1 per giorni,
mesi, anni o per tutta la vita, e per questi tipi di cellule, si dice, che G1 diventa
G0, e cioè una sorta di fase di congelamento mitotico.
D’altro canto, nelle cellule embrionali precoci, le fasi G1 e G2 hanno tempi
ristrettissimi, questo produce una replicazione mitotica a livelli esponenziali.
MITOSI
La fase M è il culmine del ciclo cellulare. Durante la meiosi (e cioè la fase di
duplicazione nucleare) e la citocinesi (fase di divisione citoplasmatica) il
contenuto della cellula parentale che è stato raddoppiato dall’attività biosintetica
della precedente interfase viene segregato in due cellule figlie.
A livello molecolare la mitosi è avviata da processi di fosforilazioni a cascata di
proteine che sono innescate dalla proteina chinasi che favorisce la mitosi (MPF).
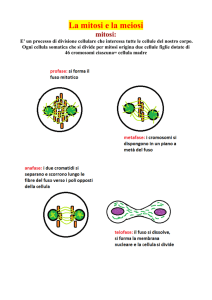
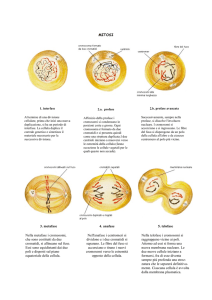
La mitosi è costituita da 6 fasi principali: Profase, Prometafase, metafase,
anafase, telofase, e la citocinesi.
PROFASE: la Profase inizia con la compattazione dei cromosomi quando i
lunghi filamenti d’eucromatina ed eterocromatina (che vanno a
formare la cromatina) diventano più corti e spessi
(SPIRALIZZAZIONE) in questo modo la cromatina può essere
distribuita alle cellule figlie senza aggrovigliarsi. La
condensazione richiede l’intervento degli ISTONI e delle
CONDENSINE. Gli istoni, e principalmente l’istone “H1” sono i
responsabili dell’impacchettamento l’uno sull’altro del DNA, la
loro attivazione è data direttamente dall’MPF, e si vanno a
collocare all’altezza del DNA di giunzione. Le condensine, invece,
sono delle proteine responsabili dell’avvolgimento e della
costituzione delle doppie anse spiralate del DNA, anche queste
sono attivate direttamente dall’MPF. Le anse spiralate vengono
compattate ancora una volta a formare un cromosoma mitotico.
Tutti i cromosomi duplicati durante la fase S, formano delle coppie
d’unità identiche, chiamati “CROMATIDI FRATELLI”. I
cromatidi fratelli si associano tra loro tramite la coesina, una
proteina che li lega all’altezza del centromero. Ad ogni centromero
è associato un cinetocoro, e cioè una struttura proteica costituito da
due lamine più dense e una meno densa, questa è la struttura che
lega i cromosomi con i microtubuli (metafase). Oltre la
condensazione della cromatina in due cromatidi fratelli si ha la
frammentazione in vescicole più piccole del reticolo
endoplasmatico (R.E.) e dell’apparato del Golgi, in questo modo
possono essere distribuiti fra le due cellule figlie al momento della
citocinesi. L’involucro nucleare e il nucleolo iniziano a sparire,
con la scomparsa del nucleolo si ha il blocco della sintesi
dell’RNA. I microtubuli interfasici si disassemblano facendo
partire la formazione del fuso mitotico, il fuso all’inizio si
assembla tra i centrosomi (costituiti da due coppie di cetrioli) che
si separano e si spostano fuori dal nucleo disponendosi verso i due
poli cellulari.
PROMETAFASE: con la prometafase si ha la fine della disgregazione
dell’involucro nucleare e l’organizzazione del fuso
mitotico.
Il fuso è costituito dai due centrosomi, dai microtubuli del
raggio, dai microtubuli polari e dai microtubuli del
cinetocoro. I microtubuli del cinetocoro si legano al
cinetocoro da entrambi i lati del centromero di ogni
cromosoma e li iniziano a spostare verso il centro della
cellula. Per questo motivo, questa divisione è descritta
come una sfera (la struttura della cellula) con un equatore
che determina il piano equatoriale (i cromosomi disposti
lungo la stessa linea centrale della cellula) e due poli
opposti (e cioè i due centrosomi nei due settori opposti). I
microtubuli polari si originano dai due centrosomi e si
irradiano verso la piastra equatoriale, ma a differenza dei
microtubuli del cinetocoro, questi si legano con gli altri
microtubuli polari del settore opposto, non legando,
quindi, i cromosomi. I microtubuli del raggio (o
dell’astrosfera) legano saldamente i centrosomi con la
membrana cellulare, non permettendoli, quindi, altri
movimenti.
METAFASE: i cromosomi si allineano ordinatamente sul piano equatoriale
della cellula, la microtubulina cresce verso il polo positivo, e
per crescere aggiunge a questo polo altra microtubulina,
facendo questo i microtubuli si spostano verso la zona
equatoriale della cellula, nel frattempo i microtubuli polari
crescono e si sovrappongono attraverso una proteina motrice
(chinesina) subito dopo si attiva una proteina che controlla che
questo allineamento sia perfetto successivamente attiva il
complesso che promuove l’anafasa (APC)
ANAFASE: la coesina localizzata nei centromeri si disaggrega consentendo
la separazione dei due cromatidi fratelli un intero corredo
cromosomico migra verso ciascun polo. Questo movimento
viene chiamato “movimento di anafase di tipo A” comportando
l’accorciamento dei microtubuli del cinetocoro attraverso la
dineina (proteina motrice) che man mano che “indietreggia”
taglia i legami di microtubulina e si sposta lungo il polo negativo
avvicinandosi ai due centrosomi. con il “movimento di anafase di
tipo B” si ha l’allungamento dei microtubuli polari attraverso la
chinesina (proteina motrice), questo produce l’assemblamento di
altre unità di microtubulina determinando l’allontanamento
reciproco dei centrosomi che portano con loro il corredo
cromosomico. I due poli opposti devono esercitare una forza di
tensione attraverso i microtubuli dell’astrosfera collegati alla
corteccia cellulare, alla fine dell’anafase li depolimerizzano
(disassemblano), questo favorisce ancora di più la separazione
dei due poli.
TELOFASE: con la telofase i cromosomi monocromatidici arrivano ai due
poli della cellula in mitosi, i cromosomi si decondensano e si
despiralizzano, attorno ad ogni serie di cromosomi compare un
involucro nucleare costituito da piccole vescicole e da altre
componenti del vecchio involucro nucleare. Si iniziano a
riformare i pori nucleari. Le proteine iniziano a ritornare nel
nucleo e il nucleolo, quindi si inizia a riformare la sinterizzazione
dell’RNA ribosomiale. Mentre i microtubuli del cinetocoro
scompaiono, quelli polari continuano a distanziare i due nuclei.
Si riformano gli organelli, i mitocondri e i perossisomi si
formano per crescita e scissione degli organelli preesistenti, il
reticolo endoplasmatico e l’apparato di Golgi (divisi in vescicole
più piccole durante G2, si associano ai microtubuli del
cinetocoro durante la separazione dei cromosomi) fondono le
vesicole separate in profase.
CITOCINESI: la citocinesi, cioè la divisione del citoplasma per produrre due
cellule figlie, comincia durante la telofase (si sovrappone alla
mitosi). un anello di microfilamenti di actina contrattili con
ponti di miosina associati alla membrana plasmatica circonda la
cellula nella regione equatoriale perpendicolarmente al fuso. La
contrazione dell’anello produce un solco di divisione che
gradualmente si approfonda e separa il citoplasma in due
cellule figlie ciascuna con un nucleo completo.
SISTEMI DI CONTROLLO DELLA MITOSI
Quando una cellula entra in G1 verifica il proprio volume e l’ambiente
circostante, una volta fatto questo la cellula continua il ciclo cellulare con la fase
S, e cioè quella di duplicazione del DNA (startpoint).
Se una cellula inizia a divedersi non può più fermarsi, ma soprattutto procede
solo in una direzione e cioè G1 -> S -> G2 -> M.
La fase successiva non inizia se non è stata accertata la conclusione a buon fine
della fase precedente.
PUNTI DI CONTROLLO
G1 (startpoint): al punto di controllo di G1, la cellula controlla le sue
dimensioni, questo controllo è fatto attraverso un meccanismo
che mette in rapporto il proprio nucleo con il citoplasma
(rapporto nucleo-citoplasmatico), inoltre verifica la presenza
dei nutrienti nel meso extracellulare (glucosio…), verifica la
presenza dei fattori di crescita che possono essere comuni o
specifici (i fattori di crescita sono degli attivatori di proteine
per andare oltre lo startpoint), e infine verifica se nel DNA
non ci siano mutazioni tali da pregiudicare la buona uscita
della mitosi.
Se uno di questi non avviene la cellula si blocca nello stato G0 (stato di
quiescenza), altrimenti supera lo startpoint e inizia a dividersi.
Tra G2 e M: un altro punto di controllo è situato tra la fine di G2 e l’ingresso
di M, qui la cellula controlla, prima di entrare in mitosi, le sue
dimensioni, e cioè se ha raddoppiato il suo volume, e se il DNA,
in fase S, sia stato completamente duplicato.
Anche qui, se uno dei meccanismi non ha avuto luogo, la cellula si blocca in uno
stato di quiescenza. Altrimenti entra in Mitosi.
METAFASE: in metafase si ha l’ultimo punto di controllo e cioè quello
dell’assemblaggio del fuso, in poche parole i cromosomi
devono essere allineati perfettamente sulla piastra equatoriale.
Se questo meccanismo non avviene, la cellula blocca la fase di duplicazione e
non entra in telofase, in questo caso la cellula va in contro ad apoptosi.
PROTEINE DEI PUNTI DI CONTROLLO
Le proteine dei punti di controllo della mitosi vanno sotto un complesso
chiamato “protein-chinasi-ciclino-dipendenti”.
Le protein-chinasi-ciclino-dipendenti attivano altre proteine tramite fosforilazioni
e per fare ciò devono agganciarsi ad una ciclica, questo meccanismo avviene
durante tutto il ciclo cellulare.
I livelli delle cicline cambiano continuamente durante il ciclo cellulare, per
questo motivo la chinasi è sempre dipendente, invece la ciclina no.
La protein-chinasi-ciclino-dipendente che si lega alla proteina di tipo G1 è quella
che agisce allo startpoint ed è quella che prepara la cellula a passare da G1 a S.
Quando la cellula supera G1 e lo startpoint, si ha un nuovo complesso di proteine
che si legano ad una ciclina, questo complesso viene chiamato S-CdK che avvia
la replicazione del DNA.
L’M-CdK è un altro complesso che promuove la mitosi (MPF).
L’MPF svolge tre funzioni, degrada l’involucro nucleare, condensa i cromosomi
(MOF) e forma il fuso mitotico; se si blocca l’MPF il ciclo cellulare si blocca.
Per attivare l’MOF, l’MPF fosforila direttamente l’istone H1 permettendo
l’impacchettamento dei cromosomi, quindi le condensine sono attivate
direttamente dall’MPF.
Un altro probabile coinvolgimento dell’MPF è nell’organizzazione del fuso
mitotico, infatti si possono trovare tracce di MPF legate saldamente ai
centrosomi.
L’MPF, inoltre, attiva il “complesso che promuove l’anafase” (APC).
MEIOSI
Nella maggior parte delle forme di riproduzione asessuata tutte le cellule sono
prodotte per MITOSI, cosicché i loro geni ed i caratteri ereditati sono identici a
quelli del genitore. Un tale gruppo di organismi geneticamente identici è
chiamato CLONE.
La riproduzione asessuata permette ad individui ben adattati al loro ambiente di
riprodurre nuove generazioni di individui ugualmente adattati la riproduzione
asessuata avviene in modo rapido ed efficiente, in parte perché l’organismo non
deve spendere energia per trovare un partner.
La riproduzione sessuata, invece, è l’unione di due cellule specializzate chiamate
GAMETI, per formare una singola cellula chiamata ZIGOTE.
I gameti, di solito, derivano da due diversi genitori: uno spermatozoo e una
cellula uovo.
La riproduzione sessuata porta a variabilità genetica all’interno della prole e cioè
sono individui geneticamente non identici ai genitori, cosicché alcuni possono
sopravvivere a variazioni ambientali e a stress meglio do loro, mentre altri che
presentano una diversa combinazione di caratteri sono meno adatti a
sopravvivere.
Un set di cromosomi contiene un cromosoma per ogni tipo, cioè un membro di
ciascuna coppia di cromosomi omologhi.
In un corredo di cromosomi dipolide ci sono 46 cromosomi (23 coppie) per ogni
cellula, in un corredo di cromosomi aploide ci sono 23 cromosomi ( non più 23
coppie, ma 23 singoli cromosomi) per ogni cellula.
La MEIOSI è una particolare divisione cellulare che riduce il numero dei
cromosomi dei gameti, questo comporta 2 divisioni citoplasmatiche e 2 divisioni
nucleari con potenziale produzione di 4 cellule aploidi.
Il DNA e gli altri componenti nucleari subiscono una sola duplicazione durante
l’interfase che precede la prima divisione meiotica; ognuna delle 4 cellule
prodotte dalla meiosi contiene un numero aploide di cromosomi; durante la
meiosi l’informazione genetica che proviene da entrambi i genitori viene
rimescolata: ogni cellula aploide prodotta contiene una combinazione di geni
potenzialmente unica.
La meiosi consiste in due divisioni nucleari e citoplasmatiche, quindi si possono
riconoscere 2 fasi meiotiche, chiamate MEIOSI I e MEIOSI II
MEIOSI I: i membri di ogni coppia di cromosomi omologhi prima si
uniscono, poi si separano e vengono distribuiti in nuclei distinti.
MEIOSI II: i cromatidi fratelli che costituiscono ciascun cromosoma omologo
si separano e vengono distribuiti ai nuclei delle cellule figlie.
MEIOSI I
Nella meiosi I troviamo a grandi linee tutte le fasi della mitosi, per cui anche qui
ci saranno: profase I, metafase I, anafase I e telofase I.
PROFASE I: durante la fase S che precede la meiosi, ciascun cromosoma
viene duplicato come nella mitosi, per cui ogni cromosoma
duplicato è costituito da due cromatidi. In profase I, mentre i
cromatidi hanno forma di lunghi e sottili filamenti, i cromosomi
omologhi si separano longitudinalmente (con complessi chiamati
SINAPSI), formando le TETRADI (in una cellula ci sono 46
coppie di cromosomi, di cui 23 coppie paterne e 23 coppie
materne, in pratica si ha un totale di 92 cromatidi). Le sinapsi
sono il punto di incontro di 2 coppie cromosomiche, una materna
e una paterna. Durante la sinapsi, i cromosomi omologhi si
associano strettamente formando quello che è chiamato
COMPLESSO SINAPTILEMALE. Si ritiene che il complesso
sinaptilemale giochi un ruolo fondamentale nel CROSSINGOVER, che è un processo in cui i cromosomi omologhi appaiati
si scambiano materiale genetico (DNA). Il crossing-over produce
nuove combinazioni di geni, la ricombinazione genetica aumenta
notevolmente la variabilità genetica, ciè le nuove combinazioni
di caratteri tra i discendenti della riproduzione sessuata. Come
nella profase mitotica, anche nella profase I meiotica si forma il
fuso costituito dai microtubuli e dalle altre componenti; una
coppia di cetrioli migra ad ogni polo e si costituisce l’aster; in
tarda profase I scompare l’involucro nucleare e si evidenziano le
tetradi. In tarda profase I, i cromatidi fratelli continuano ad
essere associati lungo il loro asse maggiore, mentre i centromeri,
e di conseguenza i cinetocoro, dei cromosomi omologhi si
separano l’uno dall’altro, restando uniti solo in corrispondenza di
regioni particolari, denominati CHIASMI. Ogni chiasma
corrisponde ad un sito in cui i cromatidi omologhi, che hanno
subito precedentemente la rottura e lo scambio di materiale
genetico (crossing-over), si sono ricongiunti e mostrano una
configurazione a χ (x).
METAFASE I: nella metafase I le tetradi si allineano lungo la piastra
equatoriale. I cinetocori fratelli di un cromosoma sono
connessi mediante le fibre del fuso ad uno dei due poli,
mentre quelli del cromosoma omologo sono collegati all’altro
polo, questa è una sostanziale differenza nei confronti della
mitosi. Poiché i cromosomi sono orientati a caso ogni cellula
figlia riceverà casualmente una parte dei cromosomi paterni e
una parte dei cromosomi materni.
ANAFASE I: in anafase I i cromosomi omologhi di ogni coppia si separano e
migrano verso i poli opposti della cellula. Ogni polo riceve
un’insieme casuale di cromosomi materni e paterni, ma un solo
rappresentante di ogni coppia di cromosomi omologhi è
presente ad ogni polo. I cromatidi fratelli sono ancora uniti
nella regione centromerica, questa è un’altra differenza con la
mitosi.
TELOFASE I: durante la telofase I i cromatidi si decondensano, l’involucro
nucleare di riorganizza ed ha luogo la citocinesi. Il nucleo
contiene il numero aploide di cromosomi (23 singoli) ma ogni
cromosoma è duplicato, perciò si avranno 23 cromosomi doppi.
INTERFASE: l’interfase è un breve periodo che si interpone tra la meiosi I e
la meiosi II; in questo periodo non si duplica il DNA, quindi
non c’è una seconda fase S, il tempo dell’interfase è
brevissimo, fino a scomparire in alcuni organismi.
PROFASE II: la profase II è molto breve perché i cromosomi sono già
condensati nel nucleo cellulare; non si ha l’accoppiamento dei
cromosomi omologhi e, soprattutto, non si ha il crossing-over.
METAFASE II: durante la metafase II, i cromosomi si allinenano sul piano
equatoriale delle rispettive cellule. Se in metafase I i
cromatidi erano disposti in gruppi di 4 (tetradi), in metafase
II i cromatidi sono disposti in gruppi di due, analogamente
alla metafase mitotica.
ANAFASE II: durante l’anafase II, i cromatidi, attaccati alle fibre del fuso
tramite i cinetocori, si separano e migrano ai poli opposti,
situazione analoga all’anafase mitotica. Ogni cromatide viene
indicato ora come cromosoma
TELOFASE II: nella telofase II i cromosomi omologhi si sono spostati ormai
nei due poli differenti, ri ricostruiscono gli organelli e il
nucleo.i cromosomi monocromatici si despiralizzano per
formare i nuovi filamenti di cromatina e si verifica la
citocinesi. Ormai la cellula madre (spermatozoo o oocita) si è
duplicata in 4 cellule figlie, ciascuna con un corredo aploide
di cromosomi, le informazione geniche delle cellule figlie,
però, sono potenzialmente diverse da quelle della madre.
GAMETOGENESI
SPERMATOGENESI: la spermatogenesi è la formazione di 4 spermatozoi
aploidi per ogni cellula. Lo spermatozoo che va in
contro a spermatogenesi è chiamato
SPERMATOCITA PRIMARIO.
OVOGENESI: la cellula uovo che va in contro a ovogenesi è definita
OOCITA PRIMARIO, l’ovogenesi è la formazione di una
singola cellula uovo per ogni oocita primario che entra in
mitosi. Questo è determinato da un processo che indirizza
tutto il citoplasma ad uno solo dei due nuclei in ciascuna
divisione meiotica. Alla fine della prima divisione meiotica
un nucleo viene mantenuto, mentre l’altro, chiamato PRIMO
GLOBULO POLARE, degenera. In modo simile alla fine
della seconda divisione meiotica, un nucleo diventa il
SECONDO GLOBULO POLARE, mentre l’altro sopravvive,
tale nucleo riceve la massima parte del citoplasma e dei
nutrienti della cellula madre che è entrata in meiosi.