Prof. Michele Segreto
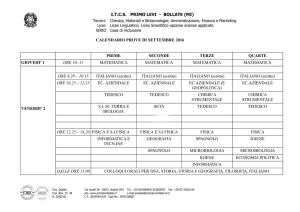
I.T.C. Primo Levi – Bollate
Ultima revisione a.s. 2012 - 2013
I Citocromi
Sono enzimi il cui coenzima presenza un gruppo eme recante uno ione ferro
Forma ossidata
Fe3+
forma ridotta
Fe2+
Esempio
2 cit.a3 (Fe2+)
+
2 cit.a3 (Fe3+)
½ O2
+ O2-
Nella respirazione aerobia il substrato di partenza, allo stato ridotto, viene
completamente ossidato con la massima produzione di energia possibile
2 H+
+
O2-
H2O
Prof. Michele Segreto
I.T.C. Primo Levi – Bollate
a.s. 2012 - 2013
Batteri ottengono energia dall’uso di substrati ridotti, sia organici
(chemiorganotrofi), sia inorganici (chemiolitotrofi).
Il substrato organico ridotto di elezione per i batteri eterotrofi è il glucosio
(C6H12O6).
La reazione globale della respirazione è
C6H12O6 + 6 O2
6 H20 + 6 CO2 + energia (ATP)
L’intero processo si svolge in due stadi
Glicolisi (o via di Embden-Meyerhof)
Ciclo di Krebs
Prof. Michele Segreto
I.T.C. Primo Levi – Bollate
a.s. 2012 - 2013
Vista la mancanza di mitocondri nei batteri, il disordine citoplasmatico porta i batteri
a svolgere il ciclo di Krebs in modo caotico e a volte ad avere una catena
respiratoria ramificata
Cit. o
Ciclo di krebs
O2
Catena respiratoria
Cit. a2
Meccanismo alternativo
in Escherichia coli
Acido lattico
Prof. Michele Segreto
I.T.C. Primo Levi – Bollate
a.s. 2012 - 2013
Batteri chemiolitotrofi - autotrofi
Usano come substrati ridotti alcuni composti inorganici come
H2
H2O
2-
H2S
SO4
NH3
NO3-
Fe2+
Fe3+
A titolo di esempio si riporta il metabolismo di due generi
batterici nitrificanti: Nitrosomonas e Nitrobacter
2 NH3 + 3 O2
2 NO2- + 2 H2O + 2 H+
NO2- + ½ O2
NO3-
Prof. Michele Segreto
I.T.C. Primo Levi – Bollate
a.s. 2012 - 2013
La respirazione anaerobia: processo catabolico tipico dei batteri chemiorganotrofi,
in cui il substrato di partenza allo stato ridotto viene ossidato attraverso un sistema di
trasporto di elettroni e idrogeno e in cui l’accettore finale non è rappresentato da
ossigeno
Gli accettori finali più comuni sono nitrato NO3- , solfato SO42-, anidride
carbonica CO2
Esempi di questo processo sono batteri denitrificanti e metanobatteri
C6H12O6 + 4 NO34 H2 + CO2
6 CO2 + 2 N2 + 6 H20 + ATP
2 H2O + CH4
Prof. Michele Segreto
I.T.C. Primo Levi – Bollate
a.s. 2012 - 2013
La fermentazione: processo catabolico in cui il substrato iniziale, donatore di
idrogeno ed elettroni, è un composto organico e l’accettore finale è rappresentato
anch’esso da un composto organico, caratterizzato da un numero inferiore di atomi
di carbonio
C6H12O6 + 2 NAD+ + 2 ADP
2 C3H4O3 + 2 NADH + H+ + 2 ATP
Acido piruvico
L’acido piruvico è la base per numerosi processi fermentativi: i più noti sono la
fermentazione alcoolica (Saccharomyces cerevisiae, Zymomonas mobilis) e lattica
(Steptococcus lactis, Lactobacillus bulgaricus)
alcoolica
lattica
C3H4O3 + NADH+H+
C3H4O3 + NADH+H+
C2H5OH + CO2 + NAD+
C3H6O3 + NAD+
Prof. Michele Segreto
I.T.C. Primo Levi – Bollate
a.s. 2012 - 2013
La fotosintesi – i batteri fotolitotrofi
Attraverso l’uso di batterioclorofilla, questi batteri (cianobatteri o alghe azzure)
sono in grado di captare la luce solare per svolgere un processo di riduzione della
CO2 secondo il seguente schema
CO2 + 2 H2O
(CH2O) + O2 + H2O
I batteri dei Generi Chromatium e Thiospirillum non usano acqua per ridurre
l’anidride carbonica, ma acido solfidrico
CO2 + 2 H2S
(CH2O) + 2 S + H2O
Prof. Michele Segreto
I.T.C. Primo Levi – Bollate
a.s. 2012 - 2013
La sintesi proteica
• Viene divisa in 5 fasi
– Attivazione degli aminoacidi
– Inizio
– Allungamento
– Terminazione e rilascio del ribosoma
– Ripiegamento e modificazioni post-traduzione
Prof. Michele Segreto
I.T.C. Primo Levi – Bollate
a.s. 2012 - 2013
Il codice genetico
• Il codice del DNA è formato da 4 lettere (A,
T, G e C)
• Il codice proteico è formato da 20
aminoacidi
• Il DNA può essere lo stampo su cui si
basa la sintesi proteica solo prevedendo
che triplette di basi azotate possano
codificare il singolo aminoacido
Prof. Michele Segreto
I.T.C. Primo Levi – Bollate
a.s. 2012 - 2013
seconda lettera
prima lettera
U
C
A
G
UUU
Phe
UCU
Ser
UAU
Tyr
UGU
Cys
UUC
Phe
UCC
Ser
UAC
Tyr
UGC
Cys
UUA
Leu
UCA
Ser
UAA
Stop
UGA
Stop
UUG
Leu
UCG
Ser
UAG
Stop
UGG
Trp
CUU
Leu
CCU
Pro
CAU
His
CGU
Arg
CUC
Leu
CCC
Pro
CAC
His
CGC
Arg
CUA
Leu
CCA
Pro
CAA
Gln
CGA
Arg
CUG
Leu
CCG
Pro
CAG
Gln
CGG
Arg
AUU
Ile
ACU
Thr
AAU
Asn
AGU
Ser
AUC
Ile
ACC
Thr
AAC
Asn
AGC
Ser
AUA
Ile
ACA
Thr
AAA
Lys
AGA
Arg
AUG
Met - inizio
ACG
Thr
AAG
Lys
AGG
Arg
GUU
Val
GCU
Ala
GAU
Asp
GGU
Gly
GUC
Val
GCC
Ala
GAC
Asp
GGC
Gly
GUA
Val
GCA
Ala
GAA
Glu
GGA
Gly
GUG
Val
GCG
Ala
GAG
Glu
GGG
Gly
U
C
A
G
Ala : alanina; Arg : arginina; Asn : asparagina; Asp : aspartato; Cys : cisteina; Gln: glutammina; Glu : glutammato;
Gly : gllicina; His : istidina; Ile : isoleucina; Leu: leucina; Lys: lisina; Met : metionina; Phe : fenilalanina; Pro : prolina;
Ser : serina; Thr : treonina; Trp : triptofano; Tyr : tirosina; Val : valina
Prof. Michele Segreto
I.T.C. Primo Levi – Bollate
a.s. 2012 - 2013
Tra le diverse triplette portate dal messaggero
(codoni) non ci sono segnali di interruzione.
La lettura del codone va in direzione 5’ – 3’
La terza base del codone spesso è
poco influente nel decidere
l’amminoacido corrispondente. La
sua funzione sembra essere quella di
velocizzare il distacco del tRNA
dall’mRNA, visto il suo legame
debole nell’appaiamento.
Il segnale di chiusura (codoni di terminazione)
del gene è dato dalle triplette STOP.
La tripletta complementare del
codone è chiamata anticodone.
Il fatto che più di una tripletta codifichi il singolo
aminoacido ha dato origine al termine di
“codice degenerato”.
Ogni transfer RNA (tRNA) porta un
anticodone specifico per
l’amminoacido corrispondente.
Il segnale di inizio è dato dal codone AUG,
quindi normalmente l’aminoacido iniziale è
metionina.
Il codice genetico è universale, ossia
praticamente tutte le specie viventi, compresi i
batteri, studiate gli aminoacidi sono 20 e sono
codificati dalle stesse triplette.
Prof. Michele Segreto
I.T.C. Primo Levi – Bollate
a.s. 2012 - 2013
Fase 1 : attivazione aminoacidi
• Avviene nel citoplasma
• Serve ad attivare il gruppo carbossilico
dell’amminoacido
• Questa fase è seguita da enzimi chiamati
aminoacil-tRNA sintetasi
• Gli enzimi sono attivati da Mg2+
Prof. Michele Segreto
I.T.C. Primo Levi – Bollate
a.s. 2012 - 2013
Fase 2 : inizio
• L’mRNA si lega alla subunità minore del
ribosoma
• Alla tripletta di inizio si lega il primo tRNA
• Le proteine che seguono questo processo
sono dette “fattori di inizio”
Prof. Michele Segreto
I.T.C. Primo Levi – Bollate
a.s. 2012 - 2013
Fase 3 : allungamento
• Il secondo codone viene letto dal ribosoma, che richiama
il tRNA complementare. Il primo aminoacido, di solito
metionina, viene legato con il legame peptidico al
secondo aminoacido.
• Il primo tRNA, libero dall’aminoacido trasportato, si
stacca dal codone e torna libero nel citoplasma.
• Il rRNA si sposta di 3 basi lungo il messaggero. Al terzo
codone si lega il tRNA corrispondente.
• L’amminoacido corrispondente viene legato al secondo
amminoacido
• Il secondo tRNA, libero dall’aminoacido trasportato, si
stacca dal codone e torna libero nel citoplasma.
Prof. Michele Segreto
I.T.C. Primo Levi – Bollate
a.s. 2012 - 2013
Fase 4 : terminazione
• Quando il ribosoma legge il codone Stop
termina di leggere il messaggero e libera
la catena polipeptidica prodotta
• Le proteine che avviano il distacco
vengono chiamate “fattori di rilascio”
• Il ribosoma inizia un nuovo ciclo produttivo
Prof. Michele Segreto
I.T.C. Primo Levi – Bollate
a.s. 2012 - 2013
Fase 5 : ripiegamento
• Il polipeptide, per poter essere
funzionante, deve prendere la sua forma
tridimensionale definitiva
• A volte alla catena amminoacidica
vengono aggiunti gruppi chimici funzionali
(gruppi acetile, fosfato, metile, ecc.)
• Altre volte vengono rimossi atomi o
molecole, anche amminoacidi, in grado di
attivare o rendere più efficiente la proteina
Prof. Michele Segreto
I.T.C. Primo Levi – Bollate
a.s. 2012 - 2013
Alcune curiosità
• Una cellula di Escherichia coli contiene oltre 15.000
ribosomi, che costituiscono il 25% del peso secco della
cellula;
• I 2/3 del peso del ribosoma è dato da RNA ribosomale
ed 1/3 da proteine;
• La funzione delle proteine non è stata ancora
completamente chiarita, ma probabilmente la più
importante è di tipo strutturale;
• Il ribosoma batterico è costituito da 2 subunità,
denominate rispettivamente 50 e 30 S, con un diametro
di 23 nm, mentre il ribosoma eucariotico è più grande
con velocità di sedimentazione di 60 e 40 S
Prof. Michele Segreto
I.T.C. Primo Levi – Bollate
a.s. 2012 - 2013
RNA transfer
•
•
•
•
•
Si tratta di molecole RNA
piccole, formate da 73 – 93
nucleotidi, con pm da 24.000 –
31.000
Ogni cellula contiene almeno
32 tipi di tRNA (alcuni
riconoscono più di un codone)
La forma, definita a trifoglio,
viene schematizzata con un
braccio lungo a cui è legato in
cima l’amminoacido
corrispondente all’anticodone
Dalla parte opposta si trova il
braccio con l’anticodone
specifico
A rinforzo della struttura
troviamo 2 anse o bracci
Prof. Michele Segreto
I.T.C. Primo Levi – Bollate
a.s. 2012 - 2013
Attivazione del tRNA
• Avviene nel citoplasma
• L’enzima responsabile dell’operazione è chiamato amminoacil-tRNA
sintetasi
• Per ogni amminoacido c’è un diverso enzima responsabile
• L’aggancio dell’amminoacido al suo tRNA richiede il consumo di 1
ATP degradato però ad AMP
• ATP
AMP + 2 Pi
• L’operazione raggiunge due scopi: attivare il messaggero e fare in
modo che possa arrivare nella posizione corretta sul messaggero
• I possibili errori nello svolgimento del processo sono corretti dalla
tRNA sintetasi (proofreading o correttore di bozze)
• La correzione è possibile solo a questo livello, una volta fatto lo
sbaglio e non corretto successivamente nelle fasi successive non
c’è modo di porre rimedio
Prof. Michele Segreto
I.T.C. Primo Levi – Bollate
a.s. 2012 - 2013
Inizio sintesi proteica
• L’importanza del codone AUG, e quindi
della metionina, nell’inizio della sintesi
proteica è testimoniato dal fatto che
troviamo 2 diversi tRNA per la metionina,
uno solo per il codone di inizio sequenza
(tRNA Met), l’altro per i codoni di centro
messaggio (tRNA fMet)
Prof. Michele Segreto
I.T.C. Primo Levi – Bollate
a.s. 2012 - 2013
Formazione del complesso di inizio nei
batteri
I fattori di inizio IF-1 e IF-3 si legano al sito
P, lo attivano e a quel punto il messaggero
si lega alla subunità minore del ribosoma.
Al sito P si appoggia il codone AUG che
codifica metionina.
Al codone si attacca il transfer UAC che
trasporta fMet. Il processo è controllato
dal fattore di inizio IF-2, attivato da GTP
che nel dare energia al processo si
trasforma in GDP, liberando una molecola
di Pi
Prof. Michele Segreto
I.T.C. Primo Levi – Bollate
a.s. 2012 - 2013
• Il secondo amminoacil-tRNA entra nel
ribosoma, si lega al sito A su cui si
trova la seconda tripletta di nucleotidi.
• Il tRNA viene attivato da un fattore di
allungamento, indicato con Tu, a sua
volta caricato con GTP, che perdendo
un Pi si trasforma in GDP fornendo
l’energia necessaria al processo
Prof. Michele Segreto
I.T.C. Primo Levi – Bollate
a.s. 2012 - 2013
• Tappe di inizio sintesi proteica
• La subunità minore del ribosoma viene attivata
mediante l’attacco di due fattori di inizio (fattore
1 e 3) al sito amminoacilico A ed in luogo
periferico (negli eucarioti i fattori di inizio sono
almeno 9)
• Il messaggero viene posizionato sulla subunità.
La direzione 5’ viene indicata dalla presenza ad
inizio messaggero di 4 – 9 basi puriniche
(sequenza di Shine-Dalgarno)
• Il tRNA Met viene posizionato sul codone di inizio
sul sito P
• Vengono liberati i fattori di inizio
• Il ribosoma si ricompone con l’aggiunta della
subunità maggiore
Prof. Michele Segreto
I.T.C. Primo Levi – Bollate
a.s. 2012 - 2013
Allungamento
• Il tRNA corrispondente alla seconda tripletta, al momento
appoggiata sul sito A, viene posizionato;
• Il GTP (guanosina tri-fosfato), legato al tranfer come attivatore,
viene idrolizzato
• La peptidil trasferasi unisce i due amminoacidi con il legame
peptidico; il primo tRNA resta privo del suo amminoacido
(traslocazione);
• Il ribosoma scivola di una tripletta verso la terminazione 3’;
• Giunto sul sito E, il tRNA scarico (deacilato) si stacca dal
messaggero
• Un nuovo tRNA, complementare alla tripletta letta dal sito A, si
appoggia sul messaggero;
• Una nuova traslocazione ha inizio
Prof. Michele Segreto
I.T.C. Primo Levi – Bollate
a.s. 2012 - 2013
Terminazione
• Dopo una serie di traslocazioni, il ribosoma legge il
segnale di fine messaggio
• Un fattore di rilascio occupa il sito A
• A questo punto i fattori di terminazione (o di rilascio)
idrolizzano l’ultimo legame che tiene unita la catena
polipeptidica con l’ultimo tRNA, in posizione P
• Il tRNA scarico viene rilasciato dal sito P
• Il ribosoma si divide nelle due subunità
• Tutti i componenti vengono rilasciati e liberano il
messaggero, che può ricominciare tutta l’operazione
Prof. Michele Segreto
I.T.C. Primo Levi – Bollate
a.s. 2012 - 2013
Regolazione dell’espressione genica
• Processo 1: trascrizione
• Processo 2: modificazioni
del mRNA post-trascrizione
• Processo 3: degradazione
del mRNA
• Processo 4: traduzione
• Processo 5: modificazione
delle proteine posttraduzione
• Processo 6: trasporto della
proteina
• Processo 7: degradazione
proteica
Prof. Michele Segreto
I.T.C. Primo Levi – Bollate
a.s. 2012 - 2013
I principi della regolazione genica
• Alcuni geni in alcune cellule sono sempre attivi: questi
sono chiamati geni costitutivi
• Il carattere non sottoposto a regolazione è chiamato
espressione genica costitutiva
• I caratteri sottoposti a regolazione ambientale sono detti
espressione genica regolata
• I prodotti dovuti a caratteri regolabili sono detti inducibili
• Il processo di attivazione dei geni regolabili è detto
induzione
• Quando il gene viene bloccato si parla di carattere
reprimibile
• Il processo che blocca il gene che determina il carattere
da fermare è detto repressione
Prof. Michele Segreto
I.T.C. Primo Levi – Bollate
a.s. 2012 - 2013
• RNA polimerasi si lega al DNA
• La trascrizione avviene solo se l’enzima riesce a
legarsi al sito promotore
• L’interazione tra promotore ed RNA non è monodimensionale, può variare a seconda delle
richieste della cellula: più il prodotto è richiesto e
maggiore è la velocità di sintesi
• I fattori di specificità modificano la capacità
dell’RNA polimerasi di legarsi al promotore e
dare inizio alle operazioni
• I repressori bloccano l’interazione tra enzima e
promotore
• Gli attivatori facilitano l’operazione
Prof. Michele Segreto
I.T.C. Primo Levi – Bollate
a.s. 2012 - 2013
• Nei batteri i siti a cui si legano i repressori sono detti
operatori e in genere sono vicini al promotore
• Se il repressore è legato all’operatore, l’RNA polimerasi
si blocca
• Il blocco dell’attività enzimatica è detto regolazione
negativa
• L’effettore è una molecola generalmente di piccole
dimensioni che funge da segnale molecolare: si lega al
repressore e lo modifica strutturalmente
• Il legame effettore – repressore modifica la velocità della
trascrizione.
• A volte l’interazione effettore – repressore modifica la
conformazione tridimensionale della molecola finale e la
lettura può avere inizio
Prof. Michele Segreto
I.T.C. Primo Levi – Bollate
a.s. 2012 - 2013
Regolazione negativa
a) Il repressore è legato all’operatore, che è
parte del promotore. La molecola segnale si
lega al repressore e si libera il promotore per
cui inizia la sintesi proteica
b) Il repressore è legato all’operatore insieme
alla molecola regolatrice. Il segnale esterno
provoca il distacco della proteina regolatrice
dal repressore che quindi libera il promotore
per cui inizia la sintesi proteica
• Gli attivatori, una volta legati al DNA, aumentano
la velocità di sintesi (regolazione positiva)
• I siti a cui si legano gli attivatori, nei procarioti, è
normalmente vicino al promotore
• A volte negli eucarioti si trovano attivatori legati a
siti detti enhancer (incrementatori) distanti dal
promotore
• A volte gli attivatori vengono attivati dalla
molecola segnale, altre volte vengono bloccati
da tale molecola. Pertanto la velocità di
trascrizione può essere sia velocizzata, sia
decrementata dalla presenza del segnale.
Prof. Michele Segreto
I.T.C. Primo Levi – Bollate
a.s. 2012 - 2013
Regolazione positiva
c) l’attivatore si lega al promotore e parte la
trascrizione. La molecola segnale si lega
all’attivatore, l’attivatore si stacca dal promotore e la
sintesi si blocca
d) l’attivatore è legato al promotore insieme alla
molecola segnale. Quando il segnale si stacca
dall’attivatore, questo si stacca dal promotore e la
sintesi si blocca
Gli operoni
• Nei procarioti si è trovato che diversi geni legati
ad un unico processo di sintesi proteica sono
riuniti in un’unica unità detta operone
• L’operone comprende diversi geni (da 2 a 6, a
volte 20), un unico sito per l’attivatore, uno per
l’operatore-repressore ed un solo promotore
• Lo studio di tali operoni è iniziato con
l’osservazione dell’operone lac in Escherichia
coli nel 1941 da J. Monod
Prof. Michele Segreto
I.T.C. Primo Levi – Bollate
a.s. 2012 - 2013
• Monod allestì colture di batteri chemioeterotrofi: il terreno di
coltura era minerale e veniva aggiunto uno zucchero per
volta
• L’aggiunta di glucosio portava ad una crescita piuttosto
veloce della colonia batterica, mentre l’aggiunta di lattosio
(un disaccaride formato da galattosio e glucosio) causava un
periodo di latenza più lungo ma una crescita altrettanto
rigogliosa
In blu la crescita normale
della colonia con
glucosio, in rosa con
substrato di difficile
assimilazione
00
00
00
00
00
00
00
00
00
0
1
3
5
7
9 11 13 15 17 19 21 23 25 27 29 31 33 35 37 39
• Mettendo entrambi gli zuccheri nel terreno di coltura si aveva una
prima crescita veloce, sovrapponibile a quella con il glucosio, a cui
segue un momento di pausa ed una seconda fase di crescita
• L’ipotesi di Monod fu che E.coli utilizzava glucosio fintanto
che trovava questo zucchero come fonte di carbonio, e solo
quando il glucosio termina, il battero rivolge le sue attenzioni
al lattosio.
• A questo tipo di crescita venne dato il nome di crescita
diauxica
• L’ipotesi, poi confermata dai lavori di laboratorio, era che
E.coli utilizzava enzimi costitutivi, quelli per l’utilizzo del
glucosio che il battero ha sempre presenti nella cellula, e
quando lo zucchero semplice termina, vengono prodotti
gli enzimi in grado di utilizzare il lattosio (enzimi
inducibili)
• Nel 1961 Jacob e Monod (premi Nobel per la medicina
nel 1965) presentarono il primo modello di regolazione
genica, l’operone lattosio.
• Lo studio parte dall’osservazione che in E.coli un solo
difetto genetico porta alla perdita dei 3 enzimi codificati
da quello che sarà poi chiamato operone lattosio
Prof. Michele Segreto
I.T.C. Primo Levi – Bollate
a.s. 2012 - 2013
L’operone lattosio
• Comprende i geni b-galattosidasi, galattoside permeasi e
tiogalattoside transacetilasi
• b-galattosidasi: sistema enzimatico complesso in grado
di catalizzare due reazioni: la principale porta all’idrolisi
del lattosio con liberazione di galattosio e glucosio, e la
secondaria in grado di convertire il lattosio in allattosio
• Allattosio è l’induttore per l’intero processo
• galattoside permeasi: è una proteina di membrana con
la funzione di catturare il lattosio presente nel mezzo di
coltura
• tiogalattoside transacetilasi: sembra che abbia la
funzione di eliminare i tiogalattosidi tossici fatti entrare
dalla permeasi
Prof. Michele Segreto
I.T.C. Primo Levi – Bollate
a.s. 2012 - 2013
• Questo modello è detto di regolazione per induzione
• I 3 geni sono adiacenti
• Il modello è di regolazione negativa: il repressore, legato
al DNA, inibisce la trascrizione in assenza di lattosio.
• Quando entra lattosio nella cellula, viene trasformato in
allattosio dalle poche molecole di galattossidasi presenti
nel citoplasma.
• La molecola segnale (l’induttore allattosio) si lega al
repressore e lo rimuove: da quel momento inizia la
lettura del DNA
Prof. Michele Segreto
I.T.C. Primo Levi – Bollate
a.s. 2012 - 2013
Ma in che modo l’associazione lattosio +
glucosio causa la crescita diauxica?
• Si è scoperto negli anni ’70 che l’operone lac è attivato
dall’induttore ma è sottoposto ad un ulteriore
meccanismo di controllo dovuto ad una proteina detta
attivatore CAP o CRP
• La proteina CRP è codificata da un gene distante
dall’operone lac
• Questa proteina viene sintetizzata in forma inattiva e
attivata da AMP ciclico (cAMP)
• La concentrazione di CRP inattivo nella cellula è
costante
• La concentrazione di cAMP è basso con concentrazioni
intracellulari alte di glucosio, si alza con l’abbassarsi
delle concentrazioni di glucosio
• Quindi cAMP funge da messaggero in grado di avvisare
E.coli della situazione cellulare del glucosio
Prof. Michele Segreto
I.T.C. Primo Levi – Bollate
a.s. 2012 - 2013
• Quindi il controllo dell’operone è doppio: alta concentrazione di
glucosio porta a basse concentrazioni di cAMP
• Viene consumato il glucosio a disposizione
• Quando il glucosio è ormai poco concentrato, si alza la
concentrazione di cAMP
• A questo punto viene attivata la proteina CRP
• CRP attivata si lega all’operone su un sito a lato del promotore
• Parte la sintesi delle proteine codificate dall’operone
• Ma se non c’è lattosio nell’ambiente l’operone è comunque bloccato
• Se c’è lattosio la galattosidasi ne trasforma una piccola parte in
allolattosio che toglie il repressore all’operone legandosi ad esso
• A questo punto parte davvero la sintesi degli enzimi dell’operone e il
lattosio viene consumato
• Viene liberato il repressore che blocca l’operone
Prof. Michele Segreto
I.T.C. Primo Levi – Bollate
a.s. 2012 - 2013
Modello dell’operone triptofano
• È la tipica repressione da prodotto finale
• In pratica quando manca il prodotto codificato dall’operone (5 geni),
il repressore è inattivo
• L’alta concentrazione del prodotto (in questo modello il triptofano
che funge da corepressore) porta all’attivazione del repressore
• Il repressore attivato si lega all’operatore e si blocca la sintesi
proteica legata all’intero operone
• Anche per questo operone troviamo un attenuatore: si tratta di una
sequenza leader (162 nucleotidi all’estremità del messaggero) che
comprende una coppia di triplette che codifica due codoni per il
triptofano, disposti in successione
• Se il triptofano è ben presente saranno presenti tRNA carichi
dell’amminoacido
• Il processo di trascrizione è contemporaneo al processo della
traduzione
Prof. Michele Segreto
I.T.C. Primo Levi – Bollate
a.s. 2012 - 2013
• La caratteristica peculiare del modello operone trp è
legata al fatto che la regolazione non è solo “produco –
non produco” ma c’è anche la possibilità di regolare la
velocità di tale processo
• Il controllo su tale meccanismo è dato da un processo
detto “attenuazione della trascrizione”
• La trascrizione viene attenuata quando la richiesta
dell’amminoacido è scarsa o ci sono molte molecole
presenti nel substrato
• Il meccanismo funziona in base alla presenza nella
regione leader di 4 sequenze (chiamate semplicemente
sequenze 1, che comprende i due codoni per triptofanotRNA, 2, 3 e 4) in grado di regolare la velocità di
produzione dell’amminoacido
• Modelli simili vengono applicati per tutte le catene
metaboliche procariotiche che portano alla produzione di
amminoacidi o di molecole che si trovano in condizioni
simili
Prof. Michele Segreto
I.T.C. Primo Levi – Bollate
a.s. 2012 - 2013
• Se nella cellula il livello del triptofano è alto, viene letta
tutta l’area del leader ma i segmenti successivi (chiamati
3 e 4) si chiudono formando una forcina (o stelo) con
l’impedimento alla continuazione della trascrizione dei
geni strutturali. Quindi si ha di fatto la creazione di un
terminatore in grado di bloccare la trascrizione degli altri
geni
• Se la quantità di triptofano è bassa lo stelo viene formato
tra i segmenti 2 e 3, quindi più a monte; il ribosoma, in
seguito alla carenza del tRNA legato al triptofano, trova i
codoni iniziali liberi (segmento 1) per cui inizia la lettura
dei geni codificati dal codone
• Questo modello non è esportabile pari pari agli eucarioti,
che avendo una membrana nucleare e la divisione tra i
processi di trascrizione e traduzione,
Prof. Michele Segreto
I.T.C. Primo Levi – Bollate
a.s. 2012 - 2013