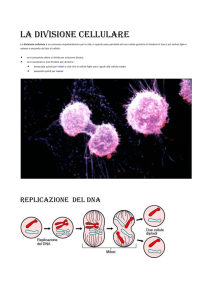
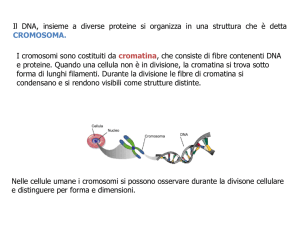
Ogni singola cellula, sia che appartenga ad un
organismo unicellulare o che sia parte di un organismo
pluricellulare, deve essere capace di crescere e di
dividersi in modo controllato.
La crescita cellulare deve essere seguita dalla divisione
cellulare, in cui da una cellula vengono generate due
cellule figlie.
Negli organismi unicellulari, la divisione cellulare fa
aumentare il numero totale degli individui di una
popolazione.
Negli organismi pluricellulari, la divisione cellulare fa
aumentare il numero di cellule e quindi permette la
crescita dell’organismo oppure serve a rimpiazzare le
cellule morte.
Il ciclo cellulare ha inizio con la formazione di due nuove cellule
figlie a partire da una cellula parentale e termina con la successiva
divisione di una di queste due.
L’interfase occupa oltre il 90% dell’intero ciclo
cellulare.
Nelle cellule dei mammiferi la durata del periodo
interfasico dipende dalla durata di G1 in quanto la
durata dei periodi S e G2 è relativamente costante.
G1: 8-10h
S:
6-8h
G2: 4-6h
M: 1-2h
Totale 18-24h
LA LUNGHEZZA DEL CICLO CELLULARE VARIA TRA I
DIVERSI TIPI CELLULARI
LABILI : Cellule del sangue
Cellule epiteliali della pelle
Cellule epiteliali che rivestono il canale digerente
La durata di G1 è di poche ore.
STABILI: fibroblasti
cellule endoteliali
cellule della muscolatura liscia
cellule epatiche
Il periodo G1 si prolunga per giorni, anche anni (Fase G0)
G2
In presenza di topoisomerasi II e di ATP
La condensina, in vitro, è in grado di legarsi al
DNA e ripiegarlo in anse superavvolte
positivamente, in presenza di una topoisomerasi
di tipo II e di ATP. Anche in vivo la
topoisomerasi è una delle principali proteine
non istoniche dell’impalcatura del cromosoma.
Sul cromosoma mitotico, esiste una marcata rientranza, detta
costrizione primaria, che individua la posizione del centromero
(sequenze di DNA altamente ripetute necessarie per il legame
con proteine specifiche). Sul lato esterno della superficie
centromerica si trova una struttura appiattita, detta cinetocore.
Questo si unisce al centromero durante la profase.
IL FUSO MITOTICO E’ RESPONSABILE DEI MOVIMENTI
CROMOSOMICI DURANTE LA MITOSI
Assemblaggio del fuso ed attacco dei cromosomi
(PROFASE e PROMETASAFE)
centromero
Microt. del cinetocore
Microt. polari
Microt. astrali
I MT polari prendono contatto con i MT provenienti dal
centrosoma opposto. Quando le due estremità positive di
due MT di polarità opposta iniziano a sovrapporsi,
intervengono delle proteine che le legano insieme
formando dei legami crociati. Come il legame tra
cinetocore e i MT del cinetocore, questi legami crociati
stabilizzano i MT polari.
Possiamo immaginare i MT che durante la profase e la
prometafase escono rapidamente a raggiera dai due
centrosomi. Quelli che riescono a incontrare un
cinetocore o un MT di polarità opposta si stabilizzano;
gli altri si disassemblano e si ritirano.
Allineamento dei cromosomi (PROMETAFASE e METAFASE)
I cromosomi migrano verso la regione centrale del fuso con una
serie di movimenti avanti e indietro generati da almeno due
forze differenti:
1. La forza di trazione esercitata dai microtubuli del cinetocore
che muove i cromosomi verso il polo a cui sono attaccati
2. La forza che tende a spingere i cromosomi lontano dai poli
del fuso
La combinazione di queste due forze fa disporre i cromosomi
sulla piastra metafasica (congressione)
Sebbene sembri che i cromosomi smettano di muoversi, l’attenta
osservazione al microscopio di cellule viventi dimostra che in
realtà i cromosomi fanno dei continui piccoli movimenti e che
essi sono sottoposti a continue tensioni in entrambe le direzioni.
Formazione della
piastra metafasica
“flusso dei microtubuli”
Mancanza di una proteina motrice (Kid),
presente lungo i bracci cromosomici in
prometafase e necessaria per il movimento
dei cromosomi dai poli verso il centro.
Proteine motrici e movimenti cromosomici in ANAFASE
All’inizio dell’anafase, i due cromatidi di ciascun cromosoma si
separano e cominciano a muoversi verso i poli opposti del fuso.
Le coesine vengono degradate da proteasi prima dell’inizio
dell’anafase, consentendo così la separazione dei cromatidi fratelli.
Le proteine motrici hanno almeno tre distinti ruoli nel movimento
dei cromosomi anafasici:
CITOCINESI: divisione del citoplasma
La citocinesi non è necessariamente connessa alla mitosi:
• in alcuni casi può trascorrere un intervallo di tempo
significativo tra la divisione nucleare e la citocinesi
• alcuni tipi di cellule possono andare incontro a molti
cicli di replicazione cromosomica e di divisione nucleare
in assenza di citocinesi dando origine a grosse cellule
multinucleate (es. sviluppo delle uova degli insetti)
La posizione del fuso
determina il punto in
cui si dividerà il
citoplasma.
Citocinesi in un uovo fecondato di rana
Il solco di divisione divide la
cellula lungo un piano che passa
per la regione centrale del fuso
(equatore del fuso).
Uovo di echinoderma diviso in 2
cellule embrionali.
la divisione avviene più
rapidamente nella cellula
cilindrica perché la distanza tra i
poli e il sito di divisione è ridotta,
diminuendo quindi il tempo
necessario affinché il segnale di
divisione raggiunga la superficie
della cellula
Il sito di assemblaggio dell’actina, e quindi il piano della
citocinesi, è determinato da segnali che provengono dai
poli del fuso. Questi segnali viaggiano lungo i microtubuli
astrali fino al cortex.