Se vi")
Lezione 9 Maggio 2013 ( Anastasi )
Se vi ricordate avevamo parlato dei recettori vestibolari , quindi macula dell' utricolo e del
sacculo e creste ampollari dei canali semicircolari ( ragazzi badate che agli esami quando
chiedo archicerebello o vestibolo voglio sapere meccanismo di deflessione, e chinociglio,
stereociglio, con la depolarizzazione e iperpolarizzazione. Certe volte chiedo
archicerebello vedo che voi partite dai recettori vedo che rispondete bene e vi dico per
vedere meglio quello che avete fatto di parlarmi dei recettori ); da quei recettori lì noi
andiamo ai nuclei vestibolari che si trovano nel pavimento del IV ventricolo, all'altezza
dell'ala bianca esterna, e poi andiamo contestualmente o con rami assonali assestanti
oppure collaterali di quelli che vanno ai nuclei vestibolari, andiamo direttamente alla
corteccia dell'archicerebello.
Riepiloghiamo: dai recettori ho le fibre del nervo vestibolare, questo è il primo nervo che
NON è pseudounipolare ma è un opposito-polare. Il ganglio si chiama ganglio di Scarpa;
le fibre hanno corpo cellulare nel ganglio di Scarpa, ramo centrifugo ai recettori, ramo
centripeto vanno ai nuclei vestibolari dell'ala bianca esterna e direttamente alla corteccia
dell'archicerebello.
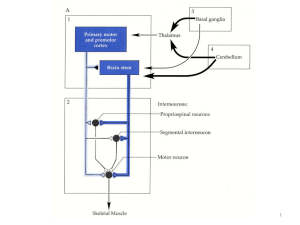
L'archicerebello proietta direttamente in uscita ai nuclei vestibolari. Stiamo attenti questo è l'unico Purkinje cerebellifugo, perché è l'unico che esce dal
cervelletto, e va agli stessi nuclei vestibolari. Allora il primo è facilitatorio, il secondo (quello che viene dalla corteccia ) è l'inibitorio, la
frequenza di spike sarà quella che caratterizzerà il vestibolo-spinale, e la somma algebrica
fra un facilitatorio ed un inibitorio ci dà il vestibolo-spinale.
Il problema è dove agisce, non basta parlare di vestibolo-spinale mediale e laterale, ma
dobbiamo anche parlare di quali motoneuroni va a beccare il pool mediale, quello cioè che
viene raggiunto dal piramidale diretto. Vi ricordate: il sistema anteriore è rappresentato dal
piramidale diretto, i due vestibolo-spinali, i due reticolo-spinali ed il tetto-spinale. Ma su quali muscoli va ? Origina dai nuclei vestibolari, (poi origina è un modo di dire dato
che abbiamo visto il circuito qual è: recettori--> nuclei vestibolari, recettori--> corteccia
dell'archicerebello, archicerebello-->nuclei vestibolari--> vestibolo spinale ).
Ma i nuclei vestibolari hanno un dispositivo somatotopico, perché mentre tutte le vie
extrapiramidali sono un confronto fra informazioni corticali ed informazioni cerebellare,
questa manca dell'informazione corticale, quindi "ragiona" senza corteccia cerebrale.
È l'unica che non è influenzata dalla corteccia cerebrale, non esistono fibre corticovestibolari. E se è autonoma deve avere il suo dispositivo somatotopico quindi deve avere
il suo homuncolus. 1
La via vestibolo-spinale laterale: termina
sulla lamina VIII, cioé non esiste una
sinapsi diretta primo neurone secondo
neurone ma è sempre mediata dalla
lamina VIII, e sulla stessa lamina VIII
agisce in effetti l'extrapiramidale ed in
questo caso il vestibolo spinale; e rare
fibre sulla lamina IX di tutto il midollo
spinale fino alla regione lombo-sacrale, ed
è importante perchè quindi voi avrete
questo per tutto il midollo spinale.
Funzione facilitatoria sui muscoli estensori
ed inibitoria sui muscoli flessori del collo, e
quindi come prima considerazione risulta che la testa risente prima di tutti le accelerazioni,
e quindi tiene la testa dritta, funzione filogeneticamente fondamentale per l'uomo che ha
come primo obiettivo quello di tenere gli occhi all'orizzonte.
Inoltre agisce sui muscoli flessori dell'arto superiore con azione sempre inibitoria, ed in
modo opposto sugli arti inferiori, tende a mantenere l'iperestensione degli arti superiori e
tende a mantenere quella sorta di pseudo flessione morbida che si ha sugli arti inferiori
( ricordate che non si ha mai un'iper estensione degli arti inferiori, quella è una posizione
artificiosa, perché biomeccanicamente l'articolazione del ginocchio non deve mai stare
serrata ma in posizione di semi-chiusura; chi ama lo scii sa che il ginocchio è in
abbondante semi-chiusura, anzi quasi in
semi-apertura ).
Via vestibolo spinale mediale: arriva alla
regione medio-toracica, quindi mentre il
vestibolo spinale laterale va per tutto il
midollo spinale qui si ferma alla mediotoracica ed insomma si fa un po' tutto l'arto
superiore ed il tronco. Funzione facilitatoria
sui muscoli estensori, ancora una volta è
quindi un antigravitario, ed è la famosa
catena posturale che prende in
considerazione gli estensori della colonna
vertebrale e gli estensori antigravitari delle
gambe, cioè regione anteriore della coscia
e posteriore della gamba.
I meccanismi compensatori vestibolocervicali e vestibolo-spinali mantengono il capo in posizione verticale rispetto alla forza di
gravità, per cui la posizione della testa è garantita dagli extrapiramidali, fa parte di quelle
attività semiautomatiche che svolgete senza l'intervento della volontà, volontariamente
cambiate la posizione della testa.
Altre tre considerazioni fondamentali: i riflessi vestibolo-cervicali si oppongono ai
movimenti del capo in modo tale da mantenerlo in posizione stabile.
Ancora i riflessi vestibolo-spinali provocano la contrazione dei muscoli degli arti e durante
la caduta preparano il soggetto al contatto con il corpo, quando cadente infatti rannicchiate
le gambe ed estendete il tronco per fornire una superficie di impatto che attutisca la
caduta.
2
Quindi è importante il vestibolo spinale alla preparazione alla perdita dell'equilibrio.
L'inclinazione del capo in avanti, questa è un'acquisizione perché quando portate il capo in
avanti inevitabilmente ( e questo lo accentuate spesso anche nella caduta perché tendete
a rannicchiare ) determina estensione degli arti superiori e flessione di quelli inferiori.
Vedete quindi come sia importante la gestione del vestibolo-spinale sul compartimento
mediale.
Quindi in posizione normale iper-estensione della testa per il mantenimento dello sguardo
all'orizzonte iper-estensione degli arti superiori, leggera morbidezza sugli arti inferiori e
comunque colonna vertebrale dritta correttamente disposta.
Quindi questa posizione della testa nello spazio è compito del vestibolo spinale non la si
assume certo volontariamente. Invece abbasso la testa rannicchio le gambe ed estendo le braccia quando il mio corpo
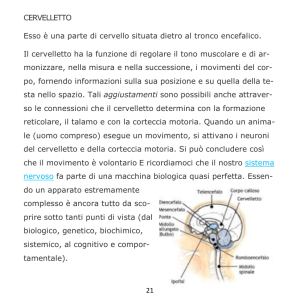
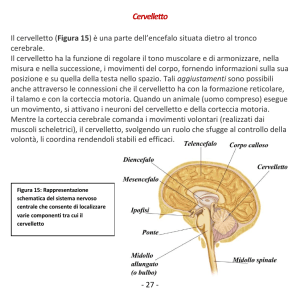
assume la posizione da opporre alla caduta. CERVELLETTO
Ha forma di cuore da carta da gioco, è una struttura sovra-assiale ed ha quindi una
corteccia esterna costituita da sostanza grigia e
la sostanza bianca invece dentro ed i nuclei
proprio.
È connesso al tronco encefalico da tre paia di
peduncoli cerebellari : • Superiori
• Medi o brachia pontis
• Inferiori o corpi restiformi
Le scissure che separano i folia cerebelli,
perché effettivamente non abbiamo dei solchi
irregolari e ripetitivi nella loro irregolarità così
come li troviamo nell'encefalo, ma abbiamo dei
solchi più profondi ma sempre ripetitivi, tanto
che aprendo il cervelletto vi darebbe
l'impressione di aprire le pagine di un libro,
come se aveste un dorso di rilegatura che è
l'ilo del cervelletto e tutto il resto sono i folia
cerebelli che si aprono fra loro.
Un folium cerebellum allora fa da un punto
all'altro del cervelletto percorrendone tutta la
circonferenza.
Ci sono due emisferi cerebellari che si
continuano l'uno nell'altro grazie ad una
formazione impari e mediana che è il verme
cerebellare, di forma convessa se visto
superiormente o concava se visto dal basso,
quindi è estroflesso rispetto agli emisferi se
visto superiormente, ed è introflesso se visto
inferiormente. Non è però una formazione
commessurale ma è una formazione impari costituita comunque da sostanza grigia e
sostanza bianca così come gli emisferi cerebellari.
3
L'ilo è la parte anteriore del cervelletto e guarda il IV ventricolo, cioè è il punto di
congiunzione dei sei peduncoli cerebellari.
In più in basso abbiamo nella zona di adesione al tronco encefalico, quindi in
corrispondenza dell'ilo c'è una parte che si chiama lobulo flocculo-nodulare, cioè ci sono
due flocculi, uno destro ed uno sinistro, raccordati da una porzione di corteccia cerebellare
che si chiama nodulo.
Se prendo tutto ciò e lo vedo in piano vedrò il lobulo flocculo-nodulare e la superficie
cerebellare, e da qui posso, secondo una logica filogenetica, distinguere:
• Archicerebello, che corrisponde al lobulo flocculo-nodulare ed è la porzione
filogeneticamente più antica del cervelletto. • Paleocerebello, che corrisponde alla zona del verme con gli emisferi adiacenti ed è la
porzione intermedia nello sviluppo filogenetico del cervelletto. • Neocerebello, che corrisponde alla zona degli emisferi e filogeneticamente parlando è
l'ultima porzione che si sviluppa. Quindi nel cervelletto ho una distinzione di tipo filogenetico.
Nello sviluppo embriologico abbiamo prima un'ipertrofia di crescita della parte bulbare per
cui per prima si formano i flocculi ed il nodulo, e solo dopo si assiste alla formazione del
cervelletto.
Guardandolo da dentro ci si deve rendere della profondità delle incisore che ci sono fra un
folium cerebellum ed un altro e poi come da ciascun in usura si diparta un'incisura
secondaria. A differenza della corteccia cerebrale ci si rende inoltre subito conto di come
qui si trovi una quantità di corteccia compattata, di netto fra l'altro superiore alla corteccia
cerebrale; microscopicamente infatti si parla di arbovite, perché la sostanza bianca si
introflette in tutti i folia presenti.
Quindi abbia una superficie cellulare notevolissima, ed il rapporto fra numero di neuroni
del cervelletto e della corteccia cerebrale non è direttamente proporzionale alla differenza
volumetrica, perché percentualmente abbiamo nel cervelletto una densità cellulare di gran
lunga superiore.
In immagine vedrete il bulbo con l'oliva bulbare ed il nucleo dentato del cervello detto
anche oliva cerebellare.
Della periferia al centro avrete come nuclei
• Nucleo dentato
• Nucleo emboliforme
• Nucleo globoso
• Nucleo del fastigio
Il nucleo globoso con l'emboliforme
formano il nucleo interposito.
Per la costituzione degli strati
riconosciamo tre strati:
• Strato dei granuli
• Strato delle cellule di Punkinje
• Strato molecolare
Lo strato dei granuli è lo strato di arrivo
delle fibre, le muscoidi.
Tutte le fibre infatti che arrivano al
cervelletto ( vestibolo-cerebellari, spinocerebellari, V-cerebellari, cortico-pontocerebellari ) sono muscoidi.
Più fibre su un granulo formano il
glomerulo cerebellare, e quando parlo del
glomerulo parlo di un terzo tipo di cellula presente nella corteccia cerebellare, ossia
4
l'interneurone di tipo II golgiano, una cellula che si trova nello strato dei granuli che
partecipa al glomerulo e riceve anche afferenze dal Punkinje. Questo rappresenta una
sorta di meccanismo servofreno, ossia quando arriva eccesso di segnale l'interneurone di
II tipo inibisce la sinapsi fibre-glomerulo. Quindi assoni dell'interneurone di II tipo, assoni
dei muscoidi e cellule dei granuli formano il glomerulo, se questo scarica troppo viene
inibito.
Per alcuni si chiamano anche cellule stellate, ma più che altro per stellate ci si riferisce ad
altre cellule differenti dagli interneuroni di II tipo.
Dai granuli nascono gli assoni che vanno allo strato molecolare e si biforcano a T,
formando lo strato delle fibre parallele.
Biforcandosi a T percorrono parallelamente l'asse della lamella; dobbiamo partire dal
presupposto che ciò che stiamo descrivendo accade in un folium o in una lamella.
Le lamelle hanno una direzione circonferenziale a tutta la circonferenza del cervelletto,
quindi sono flocculo da una parte, poi diventano verme quindi paleocerebello, poi
diventano neocerebello e alla fine tornano ad essere flocculo e nodulo.
Al flocculo ed al nodulo si concentreranno tutte le informazioni che vengono dai recettori
vestibolari, poi sulla stessa lamella troviamo le informazioni che vengono dalla corteccia
cerebrale e quindi il programma motorio, poi troviamo gli spino-cerebellari ed i Vcerebellari, e le afferenze che vengono dalla corteccia visiva. Tutto ciò è su un unico
folium, avete quindi nella zona di mezzo dove arrivano gli spino-cerebellari la ricostruzione
somato-topica frammentata.
Il cervelletto presenta questa peculiarità, non presenta l'homunculus classico.
Es. Avete due uomini seduti l'uno di spalle all'altro, la linea mediana é il punto in cui le
spalle si interfacciano ed è la linea mediana del verme, quindi questi uomini hanno i
muscoli della catena vertebrale sulla linea mediana, poi gli altri che vanno verso la zona
degli emisferi accanto al verme; magari accanto alla regione del braccio hanno la gamba,
ed accanto a quella della faccia hanno le spalle o hanno la colonna vertebrale, perché si
confrontano zone diverse del corpo umano. Quando arriva il fascio spino-cerebellare non
va su un punto, ma la zona coassiale va ad un punto poi si apre ad ombrello e va a
distanza e quindi quel punto o quella zona lo proietta sia in un punto ma
contemporaneamente in altri venti punti, e quindi si trovano accanto ad altre zone del
corpo, ed avremo quindi confronto fra braccia gamba e faccia, dove avrete i toni quindi
muscolari di zone distanti fra loro. Tutto accade su un folium. C'è una situazione di equilibrio fra le varie informazioni, cioè i famosi parametri di
frequenza ormai memorizzati.
Allora sappiamo che la posizione eretta è garantita dalla frequenza di scarica dei muscoli
della mia colonna che scaricheranno in base alle mie condizioni fisiche possono essere
più o meno ottimali, lo stesso per i muscoli degli arti e del collo e così via, ed allora se mi
muovo in condizioni normali ho come schema motorio la deambulazione, ma se
improvvisamente subisco una spinta alle spalle ho un'accelerazione lineare aumentata
che resetta i parametri degli altri e consente alle altre zone del corpo di assumere il tono
corretto per l'accelerazione subita. È la legge del più forte per cui chi scarica massima frequenza resetta tutti gli altri, se a
scaricare in massima frequenza saranno i fusi neuromuscolari degli arti inferiori perché
sottoposti ad un carico diverso dal solito,questa frequenza di scarica resetterà tutte le
altre.
Le fibre parallele corrono parallelamente all'asse della lamella.
5
A livello dello strato molecolare si ha l'integrazione del segnale, considerando un sistema
inibitorio dato dagli interneuroni di II tipo, e le fibre a T che nel loro tragitto incontrano
l'albero dendridico della cellula del Purkinje che dà dieci milioni di sinapsi per cellula.
Questo albero dendridico si estrinseca su due piani solamente, ossia non è
tridimensionale mancando della profondità, ed è un ventaglio di assoni. Il piano é inoltre
ortogonale alla lamella, quindi le fibre passano attraverso le arborizzazioni dendridiche e
prendono sinapsi. Le cellule del Purkinje stimolate faranno partire l'asse e in uscita.
Quindi fibre muscoidi in entrata arriva al granulo si forma il glomerulo parte l'asse e
arriva nello strato molecolare dove si biforca a T e nella sua corsa incontra le
arborizzazioni bidimensionali della cellula di Purkinje e quando lo eccita parte
l'impulso in discesa.
Consideriamo che abbiamo due tipi di interneuroni importanti sopratutto per dimensioni,
prima di tutto abbiamo la cellula dei canestri che si trova nello strato molecolare e corre
lungo addirittura più lamelle, e si chiama così perché periodicamente emette degli assoni
che si ramificano a canestro ed inglobano una cellula del Punkinje, ed hanno funzione
inibitoria e sono facilitate al massimo dai fili dove corre la massima frequenza di scarica.
Quindi se lungo una fila di cellule a T o un gruppo di file ho la massima eccitazione si
stimolano le cellule dei canestri che vanno ad inibire le cellule del Purkinje della lamella
accanto in un senso e nell'altro; quindi se una lamella scarica al massimo le lamelle
contigue vengono inibite. È l'inibizione cento-periferia dei campi recettorali, per cui se ho
un recettore massimamente eccitato in un punto inibisco i recettori accanto.
Questa è massima discriminazione della zona elettiva sulla quale devo agire, per cui. Per
agire su quella zona ho bisogno che le lamelle accanto siano inibite così che il segnale
parta sola da quelle lamelle. Ecco la legge del più forte.
Ancora abbiamo le cellule stellate che vanno a discriminare in un gruppo e selezionano
due cellule del Purkinje adiacenti, lavorando su due fila con lo stesso modus operandi
della cellula a canestro.
Tutto sommato lo schema è elementare: ingesso ( fibre muscoidi ), granulo, gromerulo, fibra a T, stimolo della cellula del Purkinje,
fibra in uscita; in più un controllo forzato del glomerulo grazie agli interneuroni di Golgi
posti nello strato granulari, fibre molecolari con fibre dei canestri ( inibizione a grande
distanza) fibre stellate ( inibizione a piccola distanza ).
Ad un certo punto ci si è resi conto che le cellule del Purkinje avessero azione inibitoria sui
nuclei cerebellari [ identificazione di Ito ]. L'ingresso lascia le fibre collaterali ai nuclei
cerebellari, le collaterali sono facilitatorie; il Purkinje é inibitori e dalla somma algebrica
nasce la frequenza in uscita.
In ingresso abbiamo un altro tipo di fibre che si chiamano rampicanti, queste vanno
direttamente al Purkinje senza passare dai granuli, e danno un potenziale di azione
particolare detto a denti di sega: si ha uno spike di depolarizzazione, poi abbozza la
ripolarizzazione ma mentre si trova in ripolarizzazione parte in sequenza un altro spike più
basso del primo, torna in ripolarizzazione ma mentre si trova in questa fase subisce un
altro spike. Quindi abbia tre/quattro potenziali decrescenti in fila e fra questi non c'è mai
l'isoelettrica raggiunta sola con l'ultimo piccolo spike. C'è una sola rampicante che è l'olivocerebellare, oliva intensa come oliva bulbare. Con questo tipo di potenziale la fibra è
mantenuta in iperpolarizzazione, e la mantengono refrattaria per un tempo più lungo. Il
tempo è almeno 20 volte più lungo, ed in quel momento tutte le fibre del Purkinje non sono
più eccitabili perché sono in fase di depolarizzazione continua, e quindi si bloccano le
muscoidi.
6
NON esiste il fascio olivo-spinale MA esiste lo spino-olivare, che porta propiocettivitá
incosciente e cosciente, cioè porta la propiocettivitá muscolare in genere. Al l'oliva bulbare
arriva anche la classica informazione dalla corteccia cerebrale motoria, cioè il programma
motorio dalla fibra cortico-olivare. Il risultato di integrazione fra le ultime fibre citate che ci
danno lo stato di contrazione dei nostri muscoli periferici, e lo spino-olivare che ci informa
sulla tensione presente si avrà l'informazione olivo-cerebellare. Esperimento: il primate
impara a sollevare sulla parte volare degli avambracci una sbarra dal peso x, l'operatore
gliel'abbassa e il primate la rialza. Ripetendo l'esercizio si ha uno spike complesso ( lo
spike a dente di sega ) ogni 20 spike semplici. Aumentando il peso della sbarra il primate
proverà a rialzarla ma non ci riuscirà perché applica la forza precedente, ed allora
l'operatore lo aiuterà ad alzare la sbarra istruendolo fin quando l'animale non andrà a
seguire lo stesso schema motorio di prima. In questo tempo è accaduto che al momento di
cambiamento della sbarra abbiamo avuto 20 spike complessi ed uno semplice, man mano
che l'animale però apprende diminuiscono gli spike complessi sostituiti dai semplici, finché
l'animale ha imparato e torniamo alla condizione di 1spike complesso e 20 semplici.
L'oliva bulbare quindi organizza i parametri del gesto motorio perché gli dà lo schema
motorio di partenza e la resistenza che é il risultato. Non appena le due cose non sono più
bilanciate l'oliva inizia ad avere gli spike complessi fin quando l'animale non ri-memorizza,
grazie al fatto che man mano che si hanno più spike complessi le informazioni precedenti
non vengono più percepite, data l'iperpolarizazzione del Purkinje ad opera della
rampicanti. Man mano che ha imparato ecco che le muscoidi ricominciano a lavorare col
nuovo schema motorio appreso.
Tornando ad una suddivisione verticale si identificano
• Flocculo e nodulo, dove si denotano le afferenze vestibolari
• Lo spino-cerebello, dove troviamo si afferenze visive ed uditive, che spinali e trireminali
• Il neocerebello.
( Con riferimento
all'immagine ).Riferendoci solo
alla porzione del verme del
paleocerebello; nuclei reticolari
bulbopontini, la famosa reticolare
di cui abbiamo già parlato ( fasci
spino-reticolari ) che raccoglie
tutte le collaterali di tutte le
informazioni sensitive.
Quindi corteccia cerebrale sui
nuclei reticolari bulbo-pontini ( e
su questo si trova molta
letteratura, incentrata sul fatto che
mentre si immagina di compiere
un movimento, a ritmo basale si
accendono tutti quei meccanismi
che governano il movimento
immaginato, anche se non lo
compiete ), venendo dall'area IV e
dall'area VI della corteccia
cerebrale, per dare il piano
motorio.
7
Il verme riceve afferenze spino-cerebellari che ci danno informazioni sul tono di
contrazioni, in riferimento quindi ai fusi neuromuscolari; quindi riceve anche i
quintocerebellari, le uditive e le visive ( che sono in rosso perché ed vengono da tre fonti :
• Dalla corteccia visiva
• Dal nucleo genicolato
• Dal quadrigemello superiore ) Le afferenze daranno le collaterali al nucleo proprio (ed ecco dov'è nucleo del tetto che
non sta con l'archicerebello ma con la zona del verme ), e poi arrivano al paleocerebello.
Avevamo detto che c'erano delle afferenze anche al vestibolocerebello, al flocculo ed al
nodulo, e da questo si avevo poi i nucleo vestibolari, ma le afferenze alla corteccia vanno
comunque al nucleo del tetto. Dai recettori vestibolari andiamo ai nuclei vestibolari ed
all'archicerebello, ma andando all'archicerebello andiamo anche alle collaterali al nucleo
proprio. Quindi è vero che vanno all'archicerebello ed è vero che l'archicerebello va
direttamente ai nuclei vestibolari; ma il nucleo del tetto le ha le informazioni, riceve infatti le
informazioni delle accelerazioni lineari ed angolari, le stesse che andranno
all'archicerebello per raggiungere poi i nuclei vestibolari. Quindi a livello del nucleo del
tetto si ha un confronto fra vestibolo e tono basale, e si ha confronto anche delle afferenze
visive. Ed allora il nucleo del tetto si trova in territorio di archicerebello ma ha le afferenze
anche dal paleocerebello. A questo punto il nucleo del tetto va ai nuclei reticolari bulboponti, dai quali nasce la formazione reticolo spinale.
A ciò si aggiungono anche le vie spino reticolari; quindi si ha confronto dello schema
motorio con le correzioni dovute alle interazioni cerebellari di vestibolo e spino-cerebello,
le vie spino-reticolari, senza cui non potremmo avere la posizione eretta o armonica
distribuzione delle forze e masse corporee.
Questo è il sistema che confronta il sistema dell'atto motorio con il sistema vestibolare e le
afferenze muscolari. Ma non le confronta solo col piano strategico motorio ma anche con
la realtà in cui ciò si attua.
8
Alessandro Marra
 Se vi")