LA RADICE
La radice è quella parte della pianta che, generalmente, si sviluppa al di sotto del livello del
suolo e deve garantire l’assorbimento di acqua e sali minerali che vengono trasportati in tutti gli
altri organi e tessuti della pianta e che consente di ancorarla saldamente al terreno. La radice può
essere sito di accumulo di carboidrati che derivano dalla fotosintesi clorofilliana e che sono
immagazzinati sotto forma di amido negli amiloplasti delle cellule dei parenchimi amiliferi.
L’amido accumulato nella radice ha il significato di sostanza di riserva e viene utilizzato dalla
pianta durante la ripresa vegetativa, per sviluppare nuove foglie e nuovi rami, e nel periodo della
fioritura per sviluppare gli organi riproduttivi, fiori e frutti. Altre sostanze oltre all’amido possono
essere accumulate nelle radici dando origine così ad una varietà di specializzazioni.
La radice prende origine dall’apice radicale dell’embrione e, nelle piante a seme, la prima radice
che si sviluppa è detta radice primaria o radice principale. Da questa si sviluppa un complesso
sistema di ramificazioni di radici laterali o secondarie le quali, a loro volta, danno origine ad altre
radici secondarie. La risultante di questo modello di crescita e di sviluppo è l’apparto radicale, un
complesso sistema di radici che si irradia in tutte le direzioni nel terreno e ne assicura la capillare
captazione di acqua e sali minerali, soddisfacendo così le esigenze nutritive della pianta,
interagendo intimamente con le particelle del suolo e formando un fitto sistema di ancoraggio al
terreno . Le Gimnosperme e la maggior parte delle dicotiledoni possiedono apparati radicali in cui
la radice principale resta vitale per l’intera durata della pianta e si accresce e si sviluppa (in
lunghezza e spessore) molto di più delle radici laterali che restano di gran lunga più corte e sottili.
La tipologia di crescita di questi apparati conferisce una tipica struttura spaziale che ricorda la
forma di una piramide rovesciata e che prende il nome di apparato radicale a fittone. Possiamo
avere esempio di questa struttura negli ortaggi quali barbabietola (Beta vulgaris), ravanello
(Rafanus sativum) o carota (Dacus carota) che possiedono tutti una radice principale carnosa che
costituisce la parte commestibile .
Più raramente le radici laterali si accrescono uniformemente alla radice principale, diventando
indistinguibili, o addirittura sviluppandosi di più della principale; questo modello di crescita dà
origine all’apparato fascicolato caratterizzato da molte radici di spessore e lunghezza simile. Nelle
monocotiledoni e in alcune dicotiledoni la radice principale comunemente muore dopo poco tempo
dalla germinazione del seme e l’apparato radicale della pianta si sviluppa grazie alla produzione di
radici che prendono origine alla base del fusto, generalmente in corrispondenza delle gemme
ascellari, e definite radici avventizie in quanto non si originano dai tessuti radicali (vedi scheda di
approfondimento). Da un punto di vista funzionale esse sono del tutto analoghe alle radici
secondarie, in quanto aumentano la capacità assorbente e potenziano l’ancoraggio della pianta al
terreno. Benché la forma dell’apparato radicale risultante sia a volte simile a quello fascicolato, i
due apparati differiscono per la zona istologica da cui prendono origine le singole radici.
Organizzazione della radice
Il corpo della radice, anche se abbastanza uniforme esteriormente, presenta al suo interno diverse
zone istologiche le cui cellule differiscono per velocità di crescita e modalità di differenziamento
L’estremità della radice è provvista di un tessuto di protezione detto cuffia che ricopre le cellule
sottostanti e le protegge dall’abrasione durante la penetrazione della radice nel suolo. Al di sotto
della cuffia si trova l’apice radicale, costituito da cellule la maggior parte delle quali si divide
attivamente per mitosi ed è responsabile dell’aumento del numero di cellule della radice;
analogamente a quanto accade nel fusto, quindi, l’accrescimento per divisione nella radice è
determinato dall’attività di un meristema apicale con caratteristiche strutturali e funzionali
embrionali. Allontanandosi dall’apice le cellule gradualmente perdono le caratteristiche
meristematiche e si differenziano nelle varie tipologie di cellule adulte . La zona della radice
immediatamente al di sopra dell’apice è caratterizzata da tessuti con cellule in via di
differenziazione e per questo definita zona di differenziazione. Questa zona è anche detta zona
liscia in quanto si differenzia, per aspetto esteriore, dalla zona pilifera immediatamente al di sopra,
caratterizzata dalla presenza di numerose cellule epidermiche la cui faccia esterna sviluppa delle
strutture protendenti dette peli radicali. I peli radicali sono delle strutture specializzate per
l’assorbimento che rimangono vitali. La parete dei peli radicali è ricca di mucillagini, composti con
elevata affinità per l’acqua, che amplificano la capacità assorbente di ogni singolo elemento
cellulare raddoppiando, in genere, la capacità assorbente dell’intera epidermide radicale. Inoltre
grazie alle loro piccole dimensioni i peli radicali riescono a penetrare nei più piccoli interstizi del
suolo e garantiscono una capillare estrazione di acqua e sali minerali. Tutti i tessuti della zona
pilifera sono costituiti da cellule differenziate adulte che hanno completato la crescita e
costituiscono la zona di struttura primaria. In questa zona si originano e protrudono le radici
laterali che fuoriescono dal corpo della radice principale il cui numero e disposizione è alquanto
variabile e fortemente influenzato da fattori interni quali stimoli ormonali, e/o da fattori esterni alla
radice quali la consistenza del suolo. Nelle Gimnosperme e in numerose Angiosperme dicotiledoni,
alla zona di struttura primaria segue la zona di struttura secondaria caratterizzata da tessuti
originati dall’attività dei meristemi secondari (vedi par 8.1).
La cuffia o caliptra è come si è detto la porzione terminale dell’apice radicale e costituisce un
efficace tessuto di protezione del meristema apicale .
Il suo modello di crescita infatti, è caratterizzato da un continuo rinnovamento delle cellule che la
compongono grazie all’attività di cellule iniziali (cellule del caliptrogeno) più prossime al
meristema dell’apice, mentre nella porzione terminale si ha un costante sfaldamento delle cellule
più mature. Man mano che le cellule sono spinte in avanti modificano la loro morfologia
distinguendosi in un gruppo centrale di cellule che formano la columella e altre laterali che
costituiscono le cellule periferiche. Le cellule della columella sono caratterizzate dalle presenza di
grossi granuli di amido, gli statoliti, addensati nella parte inferiore della cellula e che sono coinvolti
nella percezione della gravità orientando la direzione di crescita verso il basso delle radici (crescita
geotropica positiva). Le cellule delle fiancate laterali vanno incontro a rapida degenerazione e si
sfaldano formando una sostanza mucillaginosa che resta attaccata alle particelle di terreno e facilita
la penetrazione della radice.
L’apice radicale è costituito da cellule meristematiche dalla cui attività mitotica prendono origine
tutti i tessuti primari della radice. Nel meristema apicale si possono distinguere file di cellule (linee
cellulari) che grazie all’attività mitotica incessante vengono spinte verso l’alto sviluppandosi lungo
la radice e arrivando fino alla zona di struttura primaria. La crescita dell’apice radicale è molto
uniforme e non è soggetta a modificazioni morfologiche periodiche come avviene invece nell’apice
del germoglio; infatti, la formazione dei primordi laterali (radici laterali) si ha in una zona distante
dal meristema, contrariamente a quanto avviene nel fusto .
La zona di differenziazione si trova in contiguità con l’apice meristematico e termina in
corrispondenza della zona di struttura primaria. Si estende per qualche millimetro ed è quindi più
estesa rispetto all’apice meristematico. Analogamente a quanto accade nel fusto le cellule in questa
zona vanno incontro ad una progressiva differenziazione e in sezione trasversale di una radice in
zona di struttura primaria si distinguono chiaramente tre parti: una esterna il rizoderma, una
intermedia il cilindro corticale e una interna il cilindro centrale. Il rizoderma è un tessuto
monostratificato e riveste esternamente il corpo primario della radice. Come l’epidermide del fusto,
il rizoderma svolge una funzione protettiva, ma a differenza del fusto, le pareti delle sue cellule non
sono cutinizzate. Inoltre nel rizoderma non sono presenti gli stomi che invece caratterizzano
l’epidermide del fusto. Questo perché importante funzione del rizoderma è quella di assorbire acqua
dal terreno e trasferirla ai tessuti sottostanti, mentre attività come traspirazione e fotosintesi sono del
tutto assenti nella radice. L’assorbimento del rizoderma è potenziato dalla presenza di peli radicali
che si originano da cellule dette tricoblasti
Al di sotto del rizoderma si trova la corteccia che è molto più estesa rispetto a quella del fusto . Si
tratta di un tessuto pluristratificato le cui cellule sono di tipo parenchimatico. Queste cellule hanno
la parete sottile e grandi vacuoli. La caratteristica principale delle cellule del parenchima corticale è
che contengono numerosi amiloplasti che spesso configurano questo tessuto come un parenchima di
riserva (parenchima amilifero). L’ultimo strato della corteccia a ridosso del cilindro centrale è
costituito da cellule più piccole che costituiscono l’endoderma o endodermide. Questo tessuto è
generalmente monostratificato ed è praticamente privo di spazi intercellulari. Una caratteristica
peculiare delle cellule di questo tessuto è che le loro pareti trasversali (superiori e inferiori) e radiali
(a contatto con le altre cellule dell’endoderma) sono impregnate di suberina, una sostanza che
rende impermeabile all’acqua e a tutto ciò in essa disciolto.
La suberina avvolge come una fascia ogni singola cellula e costituisce la banda del Caspary. Dal
momento che la suberificazione della parete interessa l’intero perimetro dell’endoderma, si viene a
formare una barriera invalicabile per l’acqua che giunge per via apoplastica dal cilindro corticale.
Infatti le soluzioni che sono penetrate attraverso le pareti corticali, giunte all’endoderma non
possono infiltrarsi nelle pareti suberificate perché impermeabili e per raggiungere il cilindro
centrale devono attraversare le pareti tangenziali delle cellule endodermiche, il che comporta
attraversare il loro citoplasma. Le soluzioni passano così attraverso il “setaccio molecolare” della
membrana plasmatica che esclude molti ioni nocivi e regola la concentrazione di altri, prima di
riversarle nel tessuto vascolare.
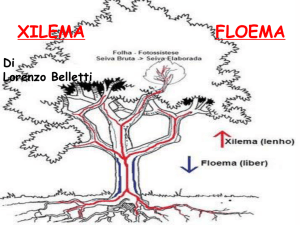
Il cilindro centrale, o stele, contiene il sistema vascolare strutturato in due tipi di tessuti di
conduzione: lo xilema o legno per il trasporto dell’acqua dalla radice alle foglie, composto dai vasi
associati a cellule parenchimatiche e fibre; il floema o libro che trasporta le sostanza elaborate dalle
foglie al resto della pianta, costituito dai tubi cribrosi, da cellule parenchimatiche e da fibre. Lo
strato più esterno del cilindro centrale a contatto con l’endoderma, detto periciclo è formato da
cellule parenchimatiche. E’ da questo tessuto che prendono origine le radici laterali e quindi viene
definito strato rizogeno. Al di sotto del periciclo è strutturato il sistema vascolare che occupa la
maggior parte del cilindro centrale. Lo xilema non è in contatto con il floema, a differenza di quanto
accade nel fusto dove i due tessuti coesistono nello stesso fascio, ma forma delle strutture raggiate o
arche xilematiche a cui si alternano le arche floematiche.
Le radici laterali si formano nella zona di struttura primaria. Esse hanno una origine endogena in
quanto si formano da un tessuto interno della radice, il periciclo. Alcune cellule del periciclo vanno
incontro a un processo di dedifferenziazione e riacquistano la capacità di dividersi per mitosi dando
origine così a tre strati di cellule. Da questi strati si originano i tre meristemi tipici dell’apice
radicale. Per attività dei meristemi la radichetta si accresce trasversalmente nel cilindro corticale
attraversando tutti i tessuti fino a fuoriuscire all’esterno .
Acrescimento secondario
In questa zona la radice è costituita da tessuti secondari che si originano cioè dall’attività di
meristemi secondari. Questa zona costituisce il restante corpo della radice e la sua struttura si
modifica e diviene via via più complessa con l’aumentare dell’età della pianta. Questo tipo di
accrescimento è il risultato dell’attività di due tessuti meristematici il cambio cribro-legnoso o
cambio e il cambio subero-fellodermico o fellogeno, che si originano dalla dedifferenziazione di
cellule parenchimatiche e per questo sono definiti meristemi secondari. Il cambio cribro-legnoso si
origina in fase avanzata di differenziazione primaria all’interno del cilindro centrale e segna il
passaggio alla struttura secondaria. Alcune cellule parenchimatiche in corrispondenza delle arche
floematiche vanno incontro ad un processo di dedifferenziazione e originano le prime cellule del
cambio che riacquistano caratteristiche meristematiche. Questo fenomeno ben presto interessa
diverse porzioni del parenchima tra le arche dando origine ad un sistema continuo di cellule
cambiali che, in sezione trasversale, presenta una forma sinusoidale. Il cambio racchiude
internamente le arche xilematiche e relega esternamente le arche floematiche. L’attività di questo
tessuto meristematico genera inizialmente solo xilema verso il lato interno (xilema secondario) e
successivamente, floema secondario verso il lato esterno, acquistando così una attività dipleurica.
Si originano così due anelli concentrici, uno interno di xilema secondario e uno esterno di floema
secondario. Con l’aumentare dell’età della pianta l’attività del cambio produce sempre nuovo legno
e nuovo libro secondari e la dimensione del cilindro centrale aumenta progressivamente
determinando l’accrescimento in spessore tipico delle piante legnose perenni .
Anche il rizoderma subisce profonde modificazioni grazie all’attività dipleurica del fellogeno che
origina esternamente sughero, a costituire i tegumenti della struttura secondaria, e internamente il
felloderma, un tessuto di natura parenchimatica. Analogamente a quanto avviene nel fusto l’insieme
di sughero, fellogeno e felloderma costituisce il periderma
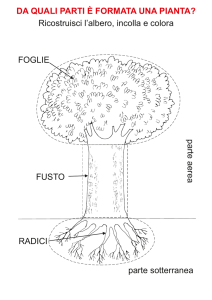
IL FUSTO
Il fusto è l’organo di sostegno delle piante e stabilisce il collegamento tra foglie e radici. In fase
giovanile può svolgere attività fotosintetica, anche se a livelli più bassi rispetto alle foglie.
Il fusto è deputato anche al trasporto attraverso
il sistema conduttore di questo organo che è
connesso con quello degli altri organi. Le sostanze assorbite dal terreno (acqua e nutrienti minerali)
sono trasportate, via xilema, dalla radice al fusto e da questo alle foglie, mentre i prodotti della
fotosintesi sono condotti, attraverso il floema, dalle foglie al fusto e da questo a tutti gli altri organi
della pianta
Il fusto può, inoltre, accumulare sostanze di riserva in cellule parenchimatiche contenenti
amiloplasti. Alcune piante, come la patata (Solanum tuberosum) ed il gladiolo (Gladiolus
glandiflorus), presentano fusti sotterranei la cui principale funzione è quella di accumulare sostanze
di riserva per garantire la ripresa dell’attività vegetativa al termine della stagione sfavorevole.
Con il termine “germoglio” ci si riferisce al sistema di organi comprendente fusto, rami, foglie e
gemme apicale e ascellari; esso rappresenta, nella generalità dei casi, la porzione aerea della pianta
La gemma apicale è costituita dal apice vegetativo (un gruppo di cellule meristematiche da cui
traggono origine i tessuti primaridel fusto), e da una serie di bozze fogliari (foglie ai primi stadi di
sviluppo) che lo avvolgono. Le foglie si inseriscono sul fusto al livello dei nodi; le regioni
comprese tra un nodo e quello successivo sono definite internodi. La distanza tra i nodi aumenta al
progredire della distanza dall’apice vegetativo, fino a raggiungere un valore massimo che tende,
poi, a mantenersi costante. Le regioni del fusto localizzate al di sopra delle inserzioni fogliari sono
dette ascelle fogliari; in corrispondenza di tali regioni si trovano le gemme laterali (o ascellari),
strutture simili alla gemma apicale, da cui traggono origine i rami laterali. Questi ultimi presentano
sviluppo, morfologia e anatomia del tutto simili a quelli del fusto. In talune piante le gemme laterali
si sviluppano formando rami di primo ordine. Le gemme laterali di questi ultimi, a loro volta, si
sviluppano in rami di secondo ordine e così via. Il fusto principale, è molto più sviluppato dei rami
di primo ordine e questi più di quelli di secondo ordine e via di seguito. Questo tipo di
Formattato: Tipo di carattere: Corsivo
ramificazione è detto monopodiale e da origine alle chiome delle conifere e, tra le angiosperme, a
Formattato: Tipo di carattere: Corsivo
quelle delle querce, dell’acero e del frassino, In altre piante il fusto principale arresta presto il suo
accrescimento e prendono a svilupparsi i rami di primo ordine, finché anche questi ultimi arrestano
il loro sviluppo e iniziano a svilupparsi quelli di secondo ordine e così di seguito. Questo tipo di
ramificazione è detta simpodiale e da origine a chiome come quelle del tiglio, dell’olmo e del
carpino e in molte piante erbacee,
Nel corpo primario del fusto, osservato in sezione longitudinale, si possono riconoscere le seguenti
zone: apice vegetativo, zona di differenziamento e zona di struttura primaria
L’apice vegetativo dà origine al corpo primario del fusto, grazie all’attività di divisione di che
derivano dalle cellule meristematiche dell’embrione, localizzate al di sopra del punto di inserzione
dei cotiledoni, dove danno origine alla gemma embrionale, la plumula. Per enfatizzare il fatto che
le cellule iniziali del cono vegetativo, così come quelle del meristema apicale radicale, derivano
dalle cellule meristematiche dell’embrione, i meristemi apicali sono anche definiti meristemi
primari.
In molte piante (nelle monocotiledoni e in un certo numero di dicotiledoni) il corpo è costituito
esclusivamente da tessuti di origine primaria; in altre (tutte le gimnosperme e numerose
dicotiledoni), oltre al corpo primario è possibile riconoscere un corpo secondario della cui
formazione sono responsabili due meristemi laterali (il cambio cribro-legnoso e cambio suberofellodermico).
Le cellule che derivano dall’apice vegetativo vanno incontro a graduale differenziamento
assumendo caratteristiche citologiche tipiche dei tessuti che costituiscono il corpo primario del
fusto. Da ciò deriva che l’apice del germoglio, durante lo sviluppo del fusto, avanzerà lasciandosi
dietro tessuti in via di differenziamento. I tessuti che compongono la struttura primaria del fusto
sono : l’epidermide che è, nella maggior parte dei casi, formata da un monostrato di cellule
appiattite e allungate in senso longitudinale, strettamente addossate le une alle altre in modo da non
lasciare spazi intercellulari. Queste La parete esterna è rivestita dalla cuticola, uno strato più o
meno spesso di sostanze idrofobe (cutina e cere). L’epidermide del fusto è dotata di stomi (sebbene
in numero inferiore rispetto alla foglia) che assicurano gli scambi gassosi; le cellule di guardia,
differentemente dalle circostanti cellule epidermiche, sono provviste di cloroplasti. Analogamente
all’epidermide fogliare, quella del fusto può presentare tricomi. Le cellule epidermiche mantengono
la capacità di dividersi mitoticamente, ciò consente a questo tessuto di adattarsi alle variazioni di
diametro del fusto. Al di sotto dell’epidermide si trova la corteccia che è formata da un certo
numero di strati di cellule, prevalentemente parenchimatiche, tra le quali si trovano ampi spazi
intercellulari. Finché il fusto è giovane e di colore verde, gli strati esterni del parenchima corticale
Formattato: Tipo di carattere: Corsivo
possono contenere cloroplasti e svolgere attività fotosintetica (in tal caso il parenchima è definito
clorofilliano) . Nella corteccia possono essere presenti, specialmente in posizione periferica, tessuti
meccanici collenchimatici e/o sclerenchimatici. Nella parte più interna della corteccia si trova, il
Cilindro centrale che è costituito dal sistema conduttore e dal midollo.
Il sistema conduttore è rappresentato dall’insieme dei fasci conduttori, ognuno dei quali è
costituito da tessuti conduttori (xilema e floema) ai quali si accompagnano frequentemente tessuti
parenchimatici (parenchima del floema e dello xilema) e meccanici (collenchima e sclerenchima).
Il numero, la posizione e la tipologia dei fasci conduttori variano nelle diverse categorie
tassonomiche. Nei fasci delle piante che presentano accrescimento secondario (tutte le
gimnosperme e parte delle angiosperme dicotiledoni), definiti collaterali aperti, è presente un
tessuto meristematico, il procambio, localizzato tra xilema e floema (il termine “collaterali” allude
al fatto che i fasci sono formati da tessuto xilematico, verso l’interno, e floematico, verso l’esterno.
Il termine “aperti” indica la presenza di cellule meristematiche tra floema e xilema). Il procambio è
coinvolto nel passaggio dalla struttura primaria a quella secondaria. I fasci delle piante che non
hanno accrescimento secondario (tutte le angiosperme monocotiledoni e parte delle angiosperme
dicotiledoni), denominati collaterali chiusi, non presentano il procambio e in essi xilema e floema
sono a diretto contatto. Nelle gimnosperme e in gran parte delle angiosperme dicotiledoni, i fasci
conduttori sono disposti in modo tale da formare un anello interposto tra corteccia e midollo. In
ogni singolo fascio lo xilema è sempre rivolto verso l’interno del fusto e il floema verso l’esterno.
Nelle angiosperme monocotiledoni la corteccia ha uno spessore molto ridotto, i fasci conduttori
sono sempre collaterali chiusi e sono immersi in tessuto parenchimatico, disposti in apparente
disordine.
Il fusto della maggior parte delle monocotiledoni, anche quello delle palme più alte, raggiunge il
diametro definitivo in pochi anni, dovuto esclusivamente ad accrescimento primario. Solo un
numero limitato di monocotiledoni, come la dracena, la yucca e l’aloe, presentano un particolare
tipo di accrescimento secondario dovuto alla formazione di un cambio.
Nella zona centrale del fusto si trova il midollo, principalmente costituito da cellule
parenchimatiche ricche di amiloplasti; la sua principale funzione è, infatti, quella di riserva. Nel
midollo si possono trovare canali resiniferi, tubi laticiferi ed anche cellule che accumulano tannini o
cristalli di ossalato di calcio. Talvolta le pareti cellulari degli strati più esterni del midollo possono
lignificare. In alcune piante, ad esempio in diverse specie della famiglia delle labiate e nella
maggior parte delle graminacee, il midollo può mancare ed il fusto risulta cavo.
Il cambio cribro-legnoso e il passaggio alla struttura secondaria
Il cambio cribro-legnoso è il meristema da cui traggono origine le cellule dello xilema secondario,
e del floema secondario. Il cambio cribro-legnoso si forma dal procambio localizzato nei fasci
conduttori fra floema e xilema primario e dalle cellule del parenchima delle regioni fra un fascio e
l’altro. Quello che si forma all’interno dei fasci conduttori primari è detto cambio intrafasciale,
mentre quello che si forma tra i fasci conduttori per sdifferenziamento delle cellule dei raggi
midollari è detto cambio interfasciale.
Le cellule del cambio dividendosi secondo piani longitudinali periclinali, originano due cellule, una
delle quali rimane iniziale, mentre l’altra si differenzia in una cellula madre del floema
secondario, se è rivolta verso l’esterno, o in una cellula madre dello xilema secondario, se rivolta
verso l’interno (vedi schema). Le cellule madri dello xilema e del floema potranno dividersi una o
più volte prima di differenziarsi, rispettivamente, in cellule xilematiche e floematiche. Per
evidenziare il fatto che le cellule del cambio cribro-legnoso producono nuovi tessuti su entrambi i
lati, si dice che questo meristema ha attività dipleurica. L’attività cambiale è più intensa verso
l’interno del fusto, producendo un maggior numero di vasi xilematici rispetto ai floematici, di
conseguenza, nell’arco di un anno viene prodotto molto più xilema che floema.
Le cellule del floema e dello xilema sono, come le iniziali fusiformi da cui derivano, allungate
secondo la direzione dell’asse maggiore del fusto e costituiscono il sistema assiale. Le cellule dei
raggi midollari, come le iniziali dei raggi da cui derivano, hanno l’asse maggiore orientato
radialmente e costituiscono il sistema radiale.
La formazione dello xilema secondario comporta un progressivo spostamento del cambio cribrolegnoso verso l’esterno e aumenta la circonferenza del fusto. Nei climi tropicali le cellule del
cambio di numerose specie si dividono in modo continuo durante l’anno. Nelle specie di regioni
temperate l’attività cambiale si interrompe durante la stagione invernale per riprendere in
primavera. I primi elementi xilematici (legno precoce), per migliorare la conduzione dell’acqua,
hanno un diametro maggiore rispetto a quelli che si formano nella stagione estiva (legno tardivo).
Questo consente di distinguere gli anelli di accrescimento del legno, ognuno dei quali si forma
durante un anno di crescita.
Il cambio subero-fellodermico
Il cambio subero-fellodermico o fellogeno è un meristema laterale costituito da un monostrato di
cellule con forma cubica che si dividono dando origine, verso l’esterno, a cellule del sughero e,
verso l’interno, a cellule che formano il felloderma. Fellogeno, sughero e felloderma formano un
complesso di tessuti denominato periderma.
Il fellogeno si origina, generalmente, per sdifferenziamento di cellule del parenchima corticale
localizzate immediatamente al di sotto dell’epidermide, ma può anche trarre origine da cellule
epidermiche, del floema primario o secondario.
Ripetute divisioni del fellogeno portano alla formazione, verso l’esterno, di più strati di sughero, un
tessuto costituito da cellule rese impermeabili da lamelle alternate di suberina e cere depositate sulla
superficie interna della parete cellulare. A maturità le cellule del sughero muoiono formando un
tessuto impermeabile specializzato per la prevenzione della perdita di acqua e dell’attacco da parte
di fitofagi. L’impermeabilità del sughero fa sì che tutti i tessuti esterni ad esso muoiano per
mancanza di nutrienti ed acqua.
L’attività del fellogeno è di breve durata per cui lo strato di sughero è, generalmente, abbastanza
sottile. Dopo uno o più anni, nuovi strato di fellogeno si formano nel floema secondario originando
molti strati di sughero sovrapposti che nel complesso costituiscono la scorza o ritidoma. Con
l’aumento di diametro del fusto, il ritidoma viene lacerato e tende a distaccarsi. Il ritidoma è perciò
formato interamente da tessuti morti.
Il sughero usato a scopi commerciali è ottenuto dalla quercia da sughero (Quercus suber), una
specie dell’area mediterranea. Questa pianta accumula molti strati di sughero originati da successivi
cambi subero-fellodermici. Il sughero viene asportato una volta ogni dieci anni, in questo modo non
si producono danni alla pianta.
L’impermeabilità del sughero comporterebbe il blocco degli scambi gassosi, se non fossero presenti
le lenticelle, insiemi di cellule prodotte dal fellogeno che nel differenziamento si arrotondano
lasciando spazi intercellulari nei quali possono circolare i gas. La lenticella si forma, generalmente,
in aree in cui, nell’epidermide, era presente uno stoma o un gruppo di stomi. Si possono rinvenire
lenticelle anche sulla buccia di alcuni frutti, come esempio pere e mele, in cui si presentano come
piccole macchie.
Le cerchie annuali
Il cambio cribro-vascolare non rimane ininterrottamente attivo durante la vita della pianta, ma
possiede un’attività periodica. Nei periodi di attività esso appone, esternamente a quelli già
esistenti, un nuovo strato di xilema. Nella sezione trasversale di un fusto è, infatti, possibile
osservare una serie di strati concentrici di xilema secondario definiti anelli di crescita. Se ciascun
anello corrisponde alla crescita di un anno, come avviene in climi caratterizzati dall’alternarsi di
stagioni favorevoli e sfavorevoli, questi sono detti cerchie annuali. È il caso dei climi temperati in
cui il cambio cribro-vascolare entra in quiescenza durante l’inverno per riprendere la sua attività in
primavera. Da quanto detto risulta che le cerchie annuali che si contano nella sezione trasversale
della base di un fusto corrispondono approssimativamente all’età della pianta.
Le cerchie annuali possono essere riconosciute grazie alle differenze strutturali tra la parte più
interna e quella più esterna di ogni strato, che si formano, rispettivamente, in primavera (legno
primaverile) e in estate (legno estivo). Generalmente, il legno primaverile è più ricco di vasi, mentre
scarseggiano le fibre, presenta cellule più larghe e con pareti meno ispessite rispetto al legno estivo.
In alcune specie, il cui legno è definito a porosità anulare, tali differenze sono molto marcate; in
altre specie con legno a porosità diffusa, le differenze tra legno formato in diversi periodi sono
meno marcate e si osserva una certa omogeneità all’interno di ciascuna cerchia.
LA FOGLIA
La foglia è uno dei tre organi costituenti il corpo della pianta ed assieme al fusto forma il
germoglio. Le sue principali funzioni sono la fotosintesi e la traspirazione; va, comunque,
sottolineato che si tratta di un organo con elevatissima plasticità evolutiva, e oltre alle tipiche foglie
con funzione clorofilliana, sono frequenti foglie secondariamente adattate alle più varie funzioni:
accumulo di nutrienti (cotiledoni e catafilli) e di acqua (foglie succulente), difesa (spine), protezione
delle gemme (perule), sostegno (viticci), cattura e digestione degli insetti e assorbimento di azoto
(foglie delle piante insettivore), vessillare (elementi del fiore), produzione di spore (sporofilli), ecc.
L’acquisizione di nuove funzioni ha comportato modificazioni nella forma, nella struttura
anatomica e nel metabolismo delle foglie.
Inserzione fogliare
La disposizione delle foglie sul fusto prende il nome di fillotassi, un carattere utile per
l’identificazione delle specie. Le foglie sono disposte sul fusto in modo da evitare, per quanto
possibile, il reciproco ombreggiamento.
Se ad ogni nodo si inserisce una singola foglia si ha fillotassi alternata , se vi si inseriscono due
foglie la fillotassi è opposta , se vi si inseriscono più di due foglie è verticillata .
Il tipo più comune di fillotassi è quello elicoidale (o a spirale), caratterizzato da una sola foglia per
ogni nodo e con foglie inserite a spirale intorno al fusto ; tale disposizione si trova, ad esempio,
nelle querce.
Morfologia fogliare
Si è detto che le principali funzioni della foglia sono la fotosintesi e la traspirazione. Il processo
fotosintetico sfrutta l’energia luminosa per trasformare l’anidride carbonica e l’acqua in zuccheri e
ossigeno . Gli zuccheri sono utilizzati in vario modo dalla pianta (come fonte energetica e come
precursori per la sintesi di altri composti organici con funzione strutturale o metabolica), l’ossigeno
è in parte impiegato per la respirazione e in parte rilasciato nell’ambiente come prodotto di scarto.
La traspirazione è la perdita dell’acqua, in forma di vapore, dalle parti aeree della pianta, ed avviene
principalmente attraverso la superficie fogliare. La traspirazione è indispensabile per il trasporto di
acqua e nutrienti minerali , ma questo processo deve essere finemente regolato, in modo da
mantenere inalterato il bilancio idrico della pianta; in altri termini, la quantità di acqua persa per
traspirazione non deve essere superiore rispetto a quella assorbita dalle radici, il che porterebbe alla
disidratazione e, oltre certi limiti, alla morte della pianta. La forma fogliare più vantaggiosa per
adempiere a queste funzioni è quella laminare, poiché ha un alto rapporto superficie/volume: quanto
più ampia è la superficie sviluppata dall’organo, tanto più intensi sono gli scambi di sostanze
gassose (vapor d’acqua, anidride carbonica e ossigeno) tra ambiente interno ed esterno, inoltre, in
una foglia sottile gran parte dei tessuti possono essere raggiunti dalla luce.
La parte espansa della foglia è detta lamina o lembo fogliare. In essa è possibile distinguere una
faccia superiore o ventrale e una faccia inferiore o dorsale.
In gran parte delle angiosperme dicotiledoni le foglie sono dotate di un peduncolo, il picciolo, che
connette la lamina fogliare al fusto. Le foglie con picciolo sono dette picciolate , quelle che ne sono
prive, caratteristiche della maggior parte delle angiosperme monocotiledoni, sono definite sessili .
Si ipotizza che il principale ruolo del picciolo sia quello di favorire la mobilità della foglia,
consentendole di assumere l’orientamento più favorevole rispetto alla direzione dei raggi solari (che
varia nel corso del dì e nelle diverse stagioni). Le foglie picciolate sono anche maggiormente
predisposte ad essere agitate dal vento; questo favorisce il ricambio di aria nell’ambiente prossimo
alla superficie fogliare, dove si potrebbe generare un’eccessiva concentrazione di ossigeno e vapor
d’acqua (rilasciati nell’ambiente aereo dalla foglia) e un’insufficiente concentrazione di anidride
carbonica (assorbita dall’ambiente aereo dalla foglia), che avrebbero effetti negativi sul traffico di
queste sostanze tra l’ambiente interno e quello esterno (che avviene per diffusione, dunque secondo
gradiente). I movimenti della foglia, provocati dal vento, contribuiscono anche ad impedire agli
insetti fitofagi, ai microrganismi patogeni e alle polveri atmosferiche, di posarsi sulla lamina
fogliare.
Alla base del picciolo possono trovarsi delle espansioni squamiformi, dette stipole, caratteristiche di
alcune dicotiledoni, come quelle appartenenti alla famiglia delle Rosaceae . In molte
monocotiledoni e in talune dicotiledoni, la base della foglia, detta guaina, può circondare il fusto e,
come avviene nelle graminacee, estendersi per l’intera lunghezza dell’internodo (fig. 10.5).
Le foglie sono dette semplici, se la lamina è intera , e composte se è suddivisa in foglioline , ognuna
dotata di un proprio peduncolo . Esistono due tipi di foglie composte: pennate e palmate. Nelle
foglie composte pennate, le foglioline si dipartono separatamente da un asse detto rachide , mentre
nelle foglie composte palmate si inseriscono al termine del picciolo . In alcune specie, come quelle
appartenenti alla mimosa, ciascuna fogliolina è ulteriormente suddivisa in più foglioline; in questi
casi la foglia è definita bipennata.
In talune specie la morfologia fogliare può variare in funzione dell’età. Ad esempio, nell’eucalipto
(Eucaliptus globulus), la giovane pianta possiede foglie ovali, sessili ed opposte, mentre la pianta
matura presenta foglie picciolate, alternate, verticali e di forma falcata . È stato ipotizzato che le
differenze nella morfologia fogliare rappresentino adattamenti ai diversi microhabitat in cui le
foglie della pianta giovane e di quella matura vengono a trovarsi.
La lamina fogliare è dotata di nervature, che costituiscono nel loro insieme il sistema conduttore
della foglia; nella maggior parte dei casi sulla faccia ventrale le nervature di maggior calibro
appaiono in rilievo. Nelle angiosperme dicotiledoni si può riconoscere una nervatura principale, che
attraversa la regione mediana della foglia lungo il suo asse maggiore, dalla quale si originano
nervature secondarie e da queste nervature più piccole; questo modello è indicato come nervazione
reticolata. La gran parte delle angiosperme monocotiledoni presentano, invece, nervazione
parallela, ossia nervature di calibro simile, parallele tra loro e con l’asse maggiore dell’organo,
connesse da nervature di calibro inferiore .
Vista l’importanza della foglia, come uno dei caratteri per l’identificazione delle specie, i botanici
hanno elaborato una ricca terminologia per definirne puntualmente la morfologia; a titolo di
esempio, alcuni dei termini più comunemente utilizzati sono riportati in (fig. 10.10).
Anatomia della foglia
Come il fusto e la radice, anche la foglia possiede una epidermide fogliare), un parenchima
clorofilliano) e un sistema conduttore (fasci conduttori fogliari). Con il termine mesofillo, ci si
riferisce all’insieme dei tessuti compresi tra l’epidermide della faccia superiore e quella della faccia
inferiore. Il mesofillo è in gran parte costituito da cellule parenchimatiche con funzione fotosintetica
(parenchima clorofilliano).
La principale funzione dell’epidermide fogliare è quella di regolare il traffico di sostanze gassose
tra ambiente interno ed esterno. Per la fotosintesi è necessario che la pianta assorba anidride
carbonica, utilizzata per la sintesi di carboidrati, e che si liberi dell’ossigeno in eccesso (la quantità
prodotta con la fotosintesi è di gran lunga superiore rispetto a quella impiegata per la
respirazione).La traspirazione, ossia la perdita di vapor d’acqua che avviene principalmente sulla
superficie fogliare, è indispensabile per il trasporto di acqua e nutrienti minerali, ma la quantità di
acqua rilasciata nell’atmosfera non deve eccedere la quantità assorbita dalle radici. L’epidermide
deve anche consentire alla luce di raggiungere i sottostanti parenchimi clorofilliani, dove viene
utilizzata come fonte energetica nel processo fotosintetico. Essa deve, infine, costituire un’efficiente
barriera contro l’attacco da parte di fitofagi e patogeni, essendo la foglia una delle parti della pianta
maggiormente “appetibili” per questi organismi. L’epidermide fogliare è costituita da cellule
appiattite incastrate l’una con l’altra come i pezzi di un puzzle, in modo da non lasciare spazi
intercellulari . Nella maggior parte delle piante l’epidermide fogliare è costituita da un singolo
strato di cellule , tuttavia, in specie adattate ad ambienti aridi , come l’oleandro (Nerium oleander) o
il ficus (Ficus elastica) può essere pluristratificata . In queste piante lo strato più esterno esplica la
funzione tegumentale mentre gli strati sottostanti assumono la funzione di tessuto acquifero (cioè
capace di immagazzinare acqua).
La parete esterna delle cellule epidermiche è impermeabilizzata, tuttavia gli scambi gassosi possono
avvenire attraverso le aperture stomatiche . Per evitare un’eccessiva traspirazione, gli stomi sono
generalmente molto più numerosi sulla pagina inferiore della foglia, in quanto meno interessata
dall’irradiazione solare e dalle correnti d’aria. In alcune piante di ambienti moderatamente umidi gli
stomi si trovano indifferentemente sulla faccia superiore e inferiore, mentre in molte altre sono del
tutto assenti sulla pagina superiore. Sulla superficie fogliare, da alcune cellule epidermiche si
differenziano spesso dei peli (o tricomi), che possono svolgere diverse funzioni: difesa chimica e
meccanica, prevenzione dagli effetti di un’eccessiva traspirazione e irradiazione solare, ecc.
Nella generalità delle dicotiledoni, nel mesofillo è possibile distinguere due tipologie di parenchima
clorofilliano: il parenchima a palizzata e il parenchima spugnoso; il primo localizzato sul lato
superioredella foglia e il secondo sul lato inferiore
Il parenchima a palizzata, il principale tessuto fotosintetico, è costituito da cellule con morfologia
colonnare, con asse principale ortogonale alla superficie dell’organo. Tra una cellula e l’altra si
trovano spazi intercellulari di ridotta ampiezza, nei quali circolano le sostanze gassose.
Generalmente il parenchima a palizzata è costituito da un singolo strato cellulare, ma in piante
adattate ad ambienti caratterizzati da elevata radiazione solare può essere pluristratificato.
Il parenchima spugnoso (o lacunoso) è costituito da cellule con forma irregolare, tra le quali si
trovano ampi spazi intercellulari. Come già accennato, nella maggior parte delle piante gli stomi
sono molto più numerosi sulla pagina inferiore della foglia. L’anidride carbonica diffonde con
facilità dall’atmosfera alle camere sottostomatiche, ampie lacune localizzate in corrispondenza
degli stomi, e da qui negli spazi intercellulari dello spugnoso fino a raggiungere le cellule del
palizzata (che sono quelle in cui il composto è maggiormente consumato). Le cellule del
parenchima spugnoso, come quelle del palizzata, possiedono cloroplasti, ma in numero
relativamente inferiore (essendo anch’esse, seppure in minor misura, fotosinteticamente attive).
Le foglie il cui parenchima clorofilliano è come sopra descritto, sono dette dorsoventrali. La
maggior parte delle dicotiledoni presenta foglie dorsoventrali.
Nel mesofillo delle foglie delle monocotiledoni, non sono distinguibili il parenchima a palizzata e
spugnoso. In varie specie del genere Allium, come l’erba cipollina (A. schoenoprasum) e l’aglio (A.
sativum), il lembo fogliare si incurva, i margini si saldano, e la foglia assume una forma cilindrica
con epidermide inferiore all’esterno e quella superiore all’interno . Un altro caso particolare è dato
dall’Iris, la cui foglia si ripiega e le due metà della faccia superiore si saldano, cosicché
l’epidermide esterna corrisponde, su entrambi i lati, all’epidermide inferiore . Le foglie sopra
descritte, in cui una delle due facce (generalmente la superiore) è ridotta o assente, sono dette
isolaterali
Il sistema conduttore della foglia è costituito dall’insieme delle nervature che sono, a loro volta,
formate dai fasci conduttori. Questi ultimi sono la prosecuzione nella foglia dei fasci conduttori del
fusto che, in corrispondenza dei nodi, divergono nel picciolo.
I fasci conduttori della foglia, come quelli del fusto, sono costituiti da xilema e da floema. Essi
presentano esclusivamente accrescimento primario , salvo rare eccezioni.
Nella foglia si possono distinguere nervature maggiori e nervature minori, le prime di calibro
superiore rispetto alle seconde. Le nervature maggiori possono essere costituite da più fasci
conduttori, mentre le minori sono composte da un singolo fascio. Le nervature maggiori, che
attraversano l’intero spessore del mesofillo, assolvono alla funzione di trasporto di acqua e nutrienti
minerali dal fusto alla foglia (attraverso lo xilema) e di fotosintati dai parenchimi clorofilliani della
foglia ai siti di utilizzo (attraverso il floema). La funzione delle nervature minori, generalmente
localizzate nel parenchima lacunoso, è quella di scambiare queste sostanze con le cellule
parenchimatiche.
Le nervature, specialmente quelle maggiori, sono costituite, oltre che da tessuti conduttori, anche da
tessuti meccanici (collenchima e sclerenchima) e parenchimatici (guaina del fascio, parenchima del
floema e dello xilema). Man mano che le nervature diminuiscono in spessore si semplifica anche la
loro struttura e la variabilità di tipi cellulari che le compongono. Nelle porzioni terminali delle
nervature di molte dicotiledoni, i fasci conduttori sono costituiti esclusivamente da xilema o floema
oppure, come avviene in molte specie, da solo xilema.
Altri tipi di foglie
I cotiledoni
I cotiledoni sono le prime foglie della plantula e si formano allo stadio embrionale. L’embrione
delle dicotiledoni possiede generalmente due cotiledoni, mentre quello delle monocotiledoni ne
possiede generalmente uno. In alcune specie con semi privi di endosperma i cotiledoni sono carnosi
e assumono la funzione di organi di riserva. In altre specie con abbondante endosperma nei semi, i
cotiledoni sono membranosi e assolvono alla funzione di assorbimento dei nutrienti
dell’endosperma, che vengono poi trasferiti alla plantula in via di sviluppo.
I cotiledoni hanno, generalmente, morfologia e struttura molto diverse rispetto a quella delle vere
foglie ed hanno una vita breve, anche nei casi, come quello del fagiolo, in cui inverdiscono e
possono, in certa misura, svolgere attività fotosintetica.
Foglie delle conifere
Nei pini (Pinus) le foglie hanno morfologia aghiforme; piccole foglie squamiformi si trovano in
altre conifere come ginepro (Juniperus) e tuia (Tuja); nelle conifere dell’emisfero boreale (Aghatis,
Araucaria e Podocarpus) le foglie sono anch’esse squamiformi, ma di maggiori dimensioni.
A titolo di esempio sarà brevemente descritta la struttura della foglia del pino . Frequentemente
l’epidermide contiene sostanze con funzione di difesa chimica. Le pareti esterne delle cellule
epidermiche sono molto spesse, lignificate e ricoperte da una spessa cuticola. Gli stomi sono
localizzati al di sotto della superficie fogliare. Al di sotto dell’epidermide si trova un ulteriore strato
tegumentale, privo di spazi intercellulari, detto ipoderma. Le cellule del tessuto fondamentale
presentano ispessimenti parietali che si proiettano all’interno della cellula, e sono incastrate le une
con le altre in modo da non lasciare spazi intercellulari. Due o più canali resiniferi, paralleli all’asse
principale della foglia, decorrono nel mesofillo. Questo è attraversato da uno o due fasci conduttori,
circondati da un tessuto costituito da cellule parenchimatiche e tracheidi, coinvolto nel trasporto di
materiali tra mesofillo e fasci conduttori, detto tessuto di trasfusione. Quest’ultimo è circondato da
uno strato tegumentale, l’endoderma.
Le foglie della gran parte delle conifere sono persistenti: quelle del pino sono mantenute
mediamente per 2-5 anni. Un caso estremo è rappresentato da Pinus longeva (la pianta vivente più
longeva) in cui le foglie rimangono sulla pianta per 45 anni. Alcune conifere come la metasequoia
(Metasequoia) e il larice (Larix) sono decidue e perdono le foglie in autunno.
1 Viticci
Alcune specie rampicanti possiedono foglie modificate, dette viticci (o cirri), per svolgere la
funzione di sostegno. Un esempio è dato dalla foglia del pisello (Pisum sativum), il cui apice è,
appunto, trasformato in un viticcio. Altri esempi sono rappresentati da varie specie della famiglia
delle Cucurbitaceae (cetriolo, zucca, ecc.).
Differentemente dalla foglie che presentano accrescimento limitato (a sviluppo ultimato arrestano
la loro crescita), i viticci presentano accrescimento indefinito (posseggono meristemi ad attività
illimitata). Un’altra sostanziale differenza è che i movimenti delle foglie sono stimolati dalla luce
(fototropismo), mentre quelli dei viticci dal contatto : i viticci si avvolgono attorno a qualunque
oggetto solido con il quale stabiliscono un contatto. Questo può avvenire grazie all’accrescimento
delle cellule che si trovano sul lato opposto a quello che ha stabilito il contatto; il processo è molto
rapido: in un’ora un viticcio può avvolgersi una o più volte.
Va, tuttavia, sottolineato che il termine viticcio non si riferisce esclusivamente a foglie modificate:
in specie quali la vite da uva (Vitis vinifera) e la vite americana (Parthenocissus quinquefolia) i
viticci non si sono originati dalle foglie, ma bensì dai rami.
Spine
Molte specie di ambienti aridi, come quelle appartenenti alla famiglia delle Cactaceae,
Euphorbiaceae e Asclepiadaceae, posseggono foglie non fotosintetizzanti, con funzione di difesa
meccanica, dette spine; la funzione fotosintetica è affidata al fusto, che si presenta, per questo, di
colore verde. Il fusto assolve, in queste specie, anche alla funzione di organo per la conservazione
dell’acqua; le piante che hanno tessuti specializzati per questa funzione sono definite succulente.
In assenza di un’adeguata difesa meccanica, nell’ambiente in cui esse vivono, dove acqua e cibo
sono scarsissimi, queste piante sarebbero divorate dagli erbivori. A questo va aggiunto che si tratta
di specie a lentissimo accrescimento, dunque, quand’anche riuscissero a sopravvivere all’attacco di
un erbivoro, avrebbero grandi difficoltà a reintegrare la biomassa asportata prima dell’attacco
successivo.
Le spine mancano di tessuti parenchimatici e conduttori essendo quasi esclusivamente formate da
fibre strettamente impacchettate, con pareti lignificate che muoiono una vota raggiunto il pieno
sviluppo.
Strutture simili alle spine, ma con diversa origine evolutiva, sono gli aculei e le emergenze: i primi
sono rami modificati mentre le seconde sono protuberanze della corteccia .
Perule
La dormienza delle gemme è di primaria importanza per la sopravvivenza delle piante perenni di
climi caratterizzati da una stagione sfavorevole (rappresentata, nei climi temperati, dall’inverno). La
gemma dormiente è un germoglio allo stato embrionale costituito da un meristema apicale, una
serie di nodi e internodi non ancora allungati, varie foglie rudimentali, primordi fogliari e delle
foglie modificate, dette perule (o squame delle gemme) che avvolgono il tutto. La perule fungono
da isolanti termici e contribuiscono a prevenire l’essiccamento, nonché a ridurre la penetrazione
dell’acqua e la circolazione di ossigeno nella gemma.
Poiché la principale funzione delle perule è quella di protezione, esse non sono fotosinteticamente
attive, hanno un picciolo breve o del tutto assente, sono molto rigide con consistenza
frequentemente coriacea e ricche, soprattutto sulla faccia esposta all’atmosfera, in sostanze idrofobe
(suberina, resine e cere).
Foglie succulente
In molte piante succulente, come l’agave (genere Agave) la funzione di conservazione dell’acqua è
affidata alle foglie .
La più evidente modificazione morfologica delle foglie succulente è data dall’aumento dello
spessore, che comporta una forte riduzione del rapporto superficie/volume e conseguentemente una
minore perdita di acqua per traspirazione. Questo causa, però, una diminuzione nella quantità di
anidride carbonica assorbita dalla foglia; inoltre, solo gli strati di parenchima più prossimi alla
superficie fogliare sono raggiunti dalla luce e quindi fotosinteticamente attivi, mentre quelli
profondi espletano esclusivamente la funzione di riserva idrica. È, quindi, evidente che il risparmio
idrico avviene a spese dell’attività fotosintetica e questo spiega, seppure solo in parte, la lentezza
del metabolismo delle piante succulente.
Foglie con funzione di riserva di nutrienti
Si è già detto, trattando le modificazioni del germoglio, che nel
bulbo di alcune specie biennali, come la cipolla (Allium cepa), le
foglie carnose assumono la funzione di organi di riserva. Queste
foglie modificate, dette catafilli, accumulano carboidrati, utilizzati
all’atto della ripresa vegetativa alla fine della stagione invernale.
Foglie delle piante insettivore
Le piante insettivore si dividono in circa 600 specie, distribuite su gran parte del pianeta: dalle calde
e umide foreste tropicali all'alta montagna, alta tanto da essere lasciata libera dagli alberi di alto
fusto. Perché carnivore? Perché integrano la loro "dieta" (costituita da acqua, sali minerali e
sostanze organiche assorbite tramite le radici) con animali, catturati e digeriti tramite speciali foglie
modificate.
I luoghi in cui queste piante vivono (paludi, rocce nude, tronchi di altre piante, ambienti sommersi),
sono poveri di sostanze azotate, che le piante carnivore possono procurarsi digerendo le prede. Le
sostanze azotate sono indispensabili per la crescita, perciò la possibilità di procurarsele anche in
ambienti dove esse scarseggiano costituisce un grande vantaggio per le piante carnivore, che
possono sfruttare un ambiente dove altre specie non riescono a vivere.
Le piante carnivore hanno escogitato diversi sistemi per la cattura delle prede, e le differenze
morfologiche dell'organo di cattura (la "trappola") consentono il riconoscimento dei generi e delle
specie. Le trappole possono essere raggruppate nelle categorie seguenti:
•
•
•
•
•
a tagliola
a carta moschicida con ghiandole invisibili
a carta moschicida con ghiandole ben visibili
ad ascidio
altre
Consideriamole in dettaglio.
TRAPPOLA A TAGLIOLA il termine tagliola deriva dal fatto che le foglie sono modificate in
modo da creare un meccanismo che si chiude a scatto quando viene sollecitato da un animaletto. La
foglia si richiude sul malcapitato intrappolandolo e soffocandolo. La preda viene poi digerita grazie
alla secrezione di enzimi digestivi dentro la tagliola stessa. Esiste una sola specie dotata di questo
tipo di trappola: Dionaea muscipula, comunemente "Dionea" o "acchiappamosche". E' diffusa negli
Stati Uniti sudorientali, in terreni acquitrinosi e soleggiati. Sopporta il freddo invernale entrando in
uno stato di dormienza, ma quando è in vegetazione richiede il pieno sole. Essa presenta una
rosetta schiacciata al terreno, che può raggiungere i 30 cm di diametro, con decine di foglie. Le
trappole possono raggiungere i 6 cm. I fiori, bianchi, sono semplici e poco vistosi, riuniti in un
capolino.
TRAPPOLA A CARTA MOSCHICIDA Molte specie hanno adottato questo meccanismo di
cattura, che consiste nella secrezione di una sostanza vischiosa sulla pagina superiore di speciali
foglie modificate. Gli insetti che vi si posano rimangono intrappolati come su una striscia di carta
moschicida. Il vischio è percepibile passando un dito sulle foglie. Di solito, sono visibili a occhio
nudo le ghiandole che secernono la sostanza vischiosa. Esse hanno la forma di un sottile filamento
sporgente dalla pagina superiore della foglia; alla loro estremità si trovano delle goccioline
traslucide contenenti le sostanze che servono ad intrappolare e digerire le prede. Le foglie di questo
tipo sono di norma mobili e nel giro di qualche ora si ripiegano sulla preda, per facilitarne la
digestione. Questo tipo di trappola, a ghiandole visibili, è adottato dai seguenti generi:
Drosera, che annovera numerose specie distribuite su tutto il pianeta ed in molti ambienti. In Italia è
presente con le specie D. rotundifolia, D. anglicae D. intermedia.
Drosophyllum: questo genere comprende solo la specie D. lusitanicum, distribuita in Spagna,
Portogallo e Marocco, molto simile ad una Drosera, con fusto lignificato.
Byblis: genere con poche specie rare, tipico di Australia e Nuova Zelanda.
Roridula: genere quasi estinto, che annovera solo due specie rarissime, presenti in Sud Africa.
Il genere Pinguicula, invece, possiede trappole con ghiandole invisibili. Le foglie sono ricoperte da
moltissime ghiandole microscopiche, la cui secrezione si stende sulla superficie della foglia come
un velo. Questo genere è distribuito in tutto il mondo; in Italia è rappresentato da sei specie, tra cui
P. vulgarise P. alpina, comunemente denominate erba unta.
TRAPPOLA AD ASCIDIO L'ascidio è una foglia modificata, che si ripiega in modo da formare
una specie di bicchiere con coperchio. Al fondo dell'ascidio si accumula un liquido che attrae gli
insetti, i quali finiscono per cadervi dentro: pessima idea, dato che nel liquido sono contenuti enzimi
digestivi che decompongono i corpi dei malcapitati. Alcuni dei generi che hanno adottato questo
sistema sono:
Nepenthes: Nelle specie appartenenti a questo genere, l'ascidio si forma all'estremità della foglia: le
piante sembrano quindi avere foglie normali con "bicchieri" pendenti dalla punta di esse. Vi sono
specie che raggiungono dimensioni di alcuni metri, con ascidi della capacità di 1-2 litri.
Cephalotus:è presente con un'unica specie australiana: C. follicularis. Essa ha due distinti tipi di
foglie: uno laminare, che è una foglia normale, l'altro a bicchiere, e quindi un ascidio, munito di
coperchio.
Sarracenia e Darlingtonia: in questi generi, l'intera foglia è ripiegata in forma di cono, con vertice
rivolto a terra. Gli insetti sono attirati all'interno del cono e cadono sul fondo, dove si trova un
accumulo di liquido digestivo: qui muoiono e sono lentamente assimilati. Alcune sarracenie
superano il metro di altezza e formano colonie con centinaia di ascidi.
Il genere Darlingtoniacomprende solo la specie D. californica, detta anche "pianta cobra" per la
forma particolare dei suoi ascidi, che assomigliano alla testa di un cobra.
Heliamphora: altro genere in cui tutte le foglie sono trasformate in ascidi conici. Le specie di
questo genere vivono in ambienti estremi sugli altopiani venezuelani e brasiliani sferzati da piogge
continue e forte vento, con temperatura notturna sottozero.
ALTRE TRAPPOLE Le specie afferenti al genere Utricularia sono piante di varia forma - alcune
ricordano piccoli cuscinetti di muschi, altre hanno grandi foglie pennate - che colonizzano ambienti
terrestri e acquatici Le utricolarie hanno trappole sotterranee o subacquee costituite da piccole (1-2
mm) vesciche sottovuoto che si aprono brevemente quando un insetto li sfiora, per poi richiudersi
inglobando l'insetto con un'azione di risucchio.
Aldrovanda vesiculosa assomiglia ad una Dionaea, ma è acquatica. Possiede piccolissime trappole
che funzionano a scatto. Un tempo comune in tutta Europa, oggi si ritiene ormai scomparsa dagli
ambienti dove era conosciuta, a causa dell'inquinamento.
Altri generi di piante carnivore, tra cui Genlisea e Ibicella, sono rarissimi e poco conosciuti.
Terminata questa panoramica sulle trappole delle piante carnivore non vi rimane altro che decidere
se prenderne una da coltivare, alcune (Nepenthes) richiedono una certa esperienza, ma non è così
per tutte. L'importante è comunque documentarsi sulle loro "preferenze" che in line adi massima
sono: acqua, torba e sole.