MECCANISMI DI COMUNICAZIONE CELLULARE
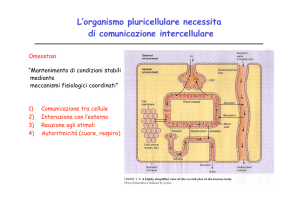
La COMUNICAZIONE CELLULARE è cruciale negli organismi multicellulari
- serve a coordinare/organizzare le attività lo sviluppo dei vari
organi/tessuti
-Lo scambio di informazioni avviene mediante rilascio di segnali
chimici e/o elettrici
Le molecole segnale (ormoni, fattori locali e neurotrasmettitori) sono di
varia natura:
-proteine (insulina, glucagone, interferone, fattori di crescita e
differenziamento, fattori di morte,…)
-aminoacidi e derivati (Glu, GABA, glicina, noradrenalina ed adrenalina,
serotonina, tiroxina),
-lipidi o steroidi (testosterone, estradiolo, prostaglandine, acido
arachidonico)
-Gas (CO e NO=ossido di azotoà contrazione muscolatura vasi sanguigni)
Struttura chimica di alcuni ormoni endocrini animali.
Le molecole segnale agiscono con un diverso raggio d’azione
Il recettore è una proteina capace
di riconoscere e legare
specificamente e con alta affinità la
molecola segnale (ligando)
(affinità molto alta per ormoni endocrini
che agiscono a conc bassissime)
Interazione recettore-ligando
funziona come interruttore
molecolare che accende una certa
via metabolicaà le funzioni
cellulari si modificano in funzione
del messaggio ricevuto
-di membrana
recettore
(molecole segnale idrofiliche)
-intracellulare
(molecole idrofobiche)
I recettori intracellulari
sono fattori trascrizionali
in seguito all’associazione
con la molecola segnale si
attivano
àe vanno a legare
sequenze di DNA
regolatore
à attivando/reprimendo
espressione genica
recettori di membrana
In seguito all’associazione con le molecole
segnale, i recettori innescano meccanismi
di trasduzione del segnale dall’esterno
all’interno della cellula
Si scatenano così eventi biochimici che
modificano varie funzioni cellulari (es
espressione genica, attività metabolica,
proliferazione, ecc)
Ligando-recettore
Trasduzione del segnale
(modificazione di enzimi e proteine,
variazione di concentrazione di ioni)
Risposta cellulare
Fasi del passaggio della informazione durante la trasduzione del segnale
Fase di trasduzione del segnale da
parte di recettori di membrana
a) Recettoreàeffettoreà modifica
[secondo messaggero]
b) Recettoreàmodifica stato di proteina
interna
(spesso a e b insieme)
Comunque sia, viene attivata una via di
segnalazione interna basata su
catena enzimi/proteine che operano in
successione: ogni enzima modifica le
proprietà dell’enzima /proteina a valle
nella sequenza à fino a convergere
su proteina bersaglio: coinvolta
direttamente in specifico processo
cellulare (es organizz. citoshceletro,
trascrizione genica, cambio
permeabilità di membrana, etc)
I meccanismi di trasduzione si basano su modificazioni delle
attività di alcuni enzimi/proteine
L’attività enzimatica può essere modulata
mediante modificazione chimica
dell’enzimaà cambi conformazionali
Es fosforilazione/defosforilazione
La maggior parte delle vie di trasduzione si basa
su attività di chinasi e fosfatasi
La fosforilazione : aggiunta di gruppi
fosfato può modificare la
conformazione di una proteina
influenzando sua attività, e/o
localizzazione
I meccanismi di trasduzione si basano anche su modificazioni delle
concentrazioni intracellulari di specifici ioni e/o piccole molecole
Es variazione [Ca 2+ ]
Il Ca funziona come
secondo messaggero in
varie vie di trasduzione
del segnale
Il Ca lega proteine Ca
dipendenti modulandone
attività
A sua volta il Ca++ lega proteine calmoduline che regolano
l’attività di vari enzimi/proteine bersaglio
Es: la calmodulina troponina
C modula contrattilità dei
filamenti nella fibra
muscolare
Variazioni di Ca controllano processo della fecondazione
Variazioni di Ca in una cellula nervosa
Ca è importante secondo
messaggero anche nelle cellule
vegetali:
es apertura /chiusura degli stomi
regolata da stato di turgore delle
cellule guardia dipendente da conc
di Ca intracellulare (Ca accumulato
nel vacuolo)
Cellula stimolata chimicamente
risponde con oscillazioni nella
conc. intracell di Ca
Il sistema di trasduzione del segnale dipende dal tipo di recettore di
membrana: 3 principali classi di recettore
1) Canale ionico
2) Associato a proteine G
3) Associato ad enzimi
Recettori associati a proteine G
Legano ligandi sul lato extracell. e interag. con proteine G trimeriche su lato
interno à la subunità Gα si attiva legando GTP e a sua volta attiva altri bersagli
•Recettore ha struttura tipica
con 7 α-eliche trans-membrana
La subunità α attivata (legante GTP) si dissocia dal trimero e
interagisce con le proteine bersaglio (inizio della via segnalazione)
lo stato attivato cessa con l’idrolisi del GTPà riassociazione nel trimero
àspegnimento automatico e attività temporalizzata
Stato attivo
Stato inattivo
proteine G
Variazione conformazionale tra stato attivo e inattivo
ciclo di attività con spegnimento automatico
Recettori associati a proteine G sono coinvolti nelle percezioni sensoriali: vista
(rodopsina e latri recettori fotosensibili presenti nella retina), olfatto e gusto (recettori
attivati da sostanza chimica specifica)
Molte vie di trasduzione attivano enzimi kinasi (PKA e PKC) che a
loro volta, fosforilando enzimi target (Ser e Treo), modulano
metabolismi vari
Es attivazione di PKA (= proteina kinasi A)
L’enzima è attivato dal legame del cAMP sulle subunità regolative
Il cAMP è un nucleotide ciclico prodotto dall’enzima adenilato ciclasi
a partire da ATP
L’attività dell’adenilato ciclasi è stimolata dalla Gα attivata
Es di via di trasduzione del segnale
innescata da recettori associati a proteine
G e che attiva PKA: Segnalazione da parte
di glucagone su epatociti
Glucagone segnala bassa glicemia
Interazione col recettore attiva Gα
àattiva adenilato ciclasi
à↑ cAMP
àattivazione di PKA che fosforila enzimi
chiave del metabolismo del glucosio
modulandone attività in modo da favorire
glicogenolisi e quindi rilascio di Glu nel
sangue
L’omeostasi del Glu dipende da complesse comunicazioni intercellulari
L’attivazione
di
adenilato
ciclasi e PKA produce risposte
cellulari diverse a seconda del
tipo di tessuto
Ad es in alcune cellule PKA fosforila
il
fattore
trascrizionale
CREB
innescando una risposta cellulare a
livello di espressione genica
L’aumento di cAMP ha varie
conseguenze a livello cellulare
Principio generale:
Cellule diverse reagiscono
diversamente allo stesso
stimolo chimico (primo e/o
secondo messaggero)
Es stesso ormone (adrenalina);
stesso recettore; cAMPà PKAà
varie risposte
Ormoni ed effetti mediati da cAMP
La PKC è attivata da recettori associati a proteine G che inducono
l’attività della fosfolipasi-C e l’aumento degli ioni Ca
La Gα attiva l’enzima di membrana fosfolipasi C che scinde un lipide di
membrana in DAG + IP3à DAG attiva PKC
IP3 stimola apertura canali Caà Ca attiva PKC
Vari secondi messaggeri derivano da fosfolipidi di membrana tagliati da varie
classi fosfolipasi (es fosfolipasi C genera IP3 e DAG dal lipide fosfatidil-inositolo (PI)
Risposte mediate da IP3 e quindi variazioni di Ca citosolico
La produzione di secondi messaggeri (Ca++, cAMP, IP3, DAG…)
permette l’amplificazione del segnale originario con un meccanismo a
cascata à risposta cellulare efficace
L’amplificazione può avvenire a diversi livelli della via trasduzionale
Amplificazione è maggiore per modulare metabolismi
minore per attivazione/repressione genica
L’ossido nitrico (NO)
È prodotto nell’endotelio in seguito a
stimolazione di recettori associati a
proteine G sulle cellule endoteliali
diffonde verso muscoloà
rilassamento della muscolatura
liscia dei vasi sanguigni
NO: molecola rilasciata dai neuroni del pene induce vasodilatazione → erezione.
Sildenafil (VIAGRA) : inibitore della fosfodiesterasi di GMP ciclico (cGMP) che
così rimane ad ↑ concentraz. per più tempo.
Le vie di trasduzione basate su cAMP e su IP3 –DAG
sono bersaglio di tossine batteriche
Tossina colerica, pertosse, tossina di alcuni ceppi di E. coli
Proteina G sempre attiva
Aumenta cAMP
La via è senza controllo così come
tutte le reazioni che scatena
(es t. colerica: inibisce attività GTPasica di Gà attivazione permanenteà permeabilità
di membrana alterata nell’epitelio intestinaleà perdita liquidià disidratazione
Mutazioni
Attivano permanent. InsP3/DAG
Divis.cell. rapida e incontrollata
Recettori associati ad enzimi
Es. recettori tirosina kinasi: transmembrana monopasso con dominio
citoplasmatico dotato di attiv.Tyr kinasica.
In seguito al legame con ligando dimerizzano e si fosforilano a vicenda
(es recettori per vari fattori di crescita EGF, PDGF, NGF, ed insulina)
Dimerizzazione dei recettori tirosina kinasi: mediata da ligando bivalente o da
recettori stessi
Una volta fosforilati i recettori richiamano sui domini citoplasmatici varie
proteine (SH2) che innescano la trasduzione intracellulare del segnale
Ad es il recettore fosforilato può associarsi a proteina che attiva Ras
(proteina G monomerica)
Ras si attiva legando GTP e regola via segnalazione che controlla
proliferazione e differenziamento cellulare
Ras innesca a cascata l’attivazione di varie kinasi MAP à
fosforilazione di fattori trascrizionali e proteine che modulano
espressione genica
(MAP= mitogen activated protein kinase)
Mutazioni di Ras provocano tumori
àRas sempre attiva
àAttivazione permanente dei geni di
proliferazione
Effetto di una mutazione dominante sul recettore Tyr chinasi per FGF
FGF: fattore di crescita dei fibroblasti.
FGFR: ruolo importante nello sviluppo
delle cellule del mesoderma
à muscolo, cartilagine,
precursori colonna vertebrale.
Embrione di rana: normale e
Mutante: con solo testa, senza
corpo.
Il recettore per insulina è un tipo peculiare di recettore associato ad enzima
Non lega direttamente proteine SH2 ma proteine di attracco IRS che poi legano
le proteine SH2
Alto Glu ematicaà stimolazione IR in muscolo e adipociti à via di segnalazione
che porta trasportatori Glut sulla membrana plasmatica mediante esocitosi :
assorbimento di Glu (trasporto facilitato)
Recettori canale
Recettori canale si aprono/chiudono in seguito al legame con molecola-segnale à
alterano polarità di membrana.
Es recettore nicotinico dell’acetilcolina sulle giunzioni neuromuscolari.
Depolarizzazione indotta dall’apertura del canale fa aprire anche canali Naà
ulteriore depolarizzazioneà apertura canali Ca del reticolo sarcoplasmatico
àcontrazione miofibrille
La contrazione del miocardio è innescata dall’ingresso di Ca attraverso canali
a controllo di potenziale
Ingresso di Ca nel citosol
àAumento di Ca citosolicaà apertura
canali membrana SER (recettori
rianodinici)à aumento ulteriore Ca
citoslicaà contrazione
àRitorno a stato inizialae grazie a
pompe attive
Ciclo si ripete ad ogni battito
Le varie vie di segnalazione, iniziate da diversi recettori, possono
convergere e dialogare tra loro
La risposta recettoriale deve essere temporalmente definita à
meccanismi di “spegnimento” della risposta recettoriale
-Molti elementi del sistema di trasduzione si autoinattivano (es G protein) o
sono abbinati a sist. di spegnimento (cAMP fosfodiesterasi, pompe Ca,…)
Esistono anche strategie di
desensibilizzazione ad un certo
stimolo
Es endocitosi del recettore-ligando seguita
da riciclo o degradazione del recettore (a-b)
Intervento di proteina inibitrice sul
recettore o su anello + a valle della via
trasduzione (c-d)
A volte la stessa via trasduzione attiva
inibitore (e)
Esempio di desensibilizzazione:
meccanismo che previene riposta a stimolo
persistente (risposta a variazioni di stato)
Il legame tra ligando e recettore fa
sì che quest’ultimo sia fosforilato
da GRKà legame con arrestinaà
blocco via segnalazione