30/04/15
L’ATP è sintetizzato in
tre principali modi:
mediante fosforilazione a
livello del substrato,
mediante fosforilazione
ossidativa,
mediante fotofosforilazione
1
30/04/15
Prima via metabolica ad essere compresa.
Universalmente diffusa in tutte le cellule.
Avviene nel citosol di tutte la cellule
Una molecola di
glucosio viene
degradata in una
serie di reazioni
catalizzate da
enzimi per produrre
due molecole di
piruvato
Importante ruolo metabolico centrale
La glicolisi avviene in 10 tappe
Fase
preparatoria di
investimento
energetico
- 2 ATP
Fase di
recupero
energetico
+ 4 ATP
+ 2 equivalenti di
riduzione NADH
Negli organismi aerobici la glicolisi è il primo passaggio per la
combustione completa del glucosio a CO2 e H2O
Negli organismi anaerobici è l’unica via per la produzione di ATP
La conversione del glucosio a piruvato nelle cellule
in grado di respirare è detta glicolisi aerobica
2
30/04/15
L’ATP è sintetizzato
in tre principali modi:
mediante
fosforilazione
fosforilazioneaalivello
del
substrato,
livello
del substrato,
fosforilazione
ossidativa,
fotofosforilazione
3
30/04/15
Investimento
energetico per
aumentare
l’energia libera
degli intermedi
della via
metabolica.
Gli zuccheri
fosforilati non sono
più capaci di
attraversare la
membrana cellulare
Le catene
metaboliche di tutti
gli esosi sono
convertite in due
molecole di
gliceraldeide-3fosfato
La resa netta
della glicolisi
sono
2 molecole
di ATP per
molecola di
glucosio.
Altra energia
è conservata
n e l l a
formazione di
2 molecole
di NADH
4
30/04/15
Nelle cellule
eucariotiche la glicolisi
avviene nel CITOSOL,
mentre la successiva
ossidazione del
piruvato avviene nei
mitocondri.
In alcuni tripanosomi
avviene in un organulo
citoplasmatico detto
glicosoma
5
30/04/15
1°
Esochinasi fosforila il glucosio intrappolandolo nella cellula infatti il
glucosio 6-fosfato non può uscire dalla cellula.
ISOENZIMA
Negli epatociti :
glucochinasi che
differisce
dall’esochinasi per
diverse proprietà
cinetiche e
regolatorie
QUESTE FORME ISOENZIMATICHE
Consentono al cervello e al muscolo di usare per
primi il glucosio quando la concentrazione nel
sangue è bassa
L’esochinasi nei diversi organismi
è generalmente caratterizzata da:
1. bassa specificità per gli zuccheri
2. bassa Km (0,1 mM)
3. Inibizione retroattiva da
parte del suo prodotto,
glucosio-6-fosfato.
Concentrazione di glucosio
nel sangue = 5mM
La glucochinasi nel fegato è
invece caratterizzata da :
1.Specificità per il glucosio
2. Più alta Km (10 mM)
3. Non presenta inibizione
retroattiva da prodotto
Regolazione al livello del substrato.
6
30/04/15
Effetto della
concentrazione del
glucosio del sangue
sulla velocità di
fosforilazione
catalizzata dalla
glucochinasi (nel
fegato, rimuove
glucosio dal sangue
dopo i pasti)
e dalla
esochinasi (nei
tessuti diversi dal
fegato è in saturazione
di substrato e funziona
a ritmo costante per
fornire glucosio 6fosfato alle cellule)
1
7
30/04/15
Prima reazione dove si ha conservazione di energia
8
30/04/15
La formazione
di ATP mediante
il trasferimento
di gruppi
fosforici da un
substrato come
1,3-bisfosfoglicerato viene detta
FOSFORILAZIONE AL LIVELLO DEL SUBSTRATO
per distinguerla dalla fosforilazione legata alla respirazione.
Seconda
fosforilazione
al livello del
substrato
Glucosio + 2ATP + 2NAD+ + 4ADP + 2 Pi
2 piruvato +2ADP + 2NADH + 2H+ + 4ATP + 2H2O
Cancellando i termini comuni
Glucosio + 2NAD+ + 2ADP + 2Pi
2 piruvato + 2NADH + 2H+ + 2ATP + 2H2O
Devono venire riossidate
9
30/04/15
Affinché la via glicolitica
possa operare, il NADH deve
essere riossidato a NAD+
mediante il trasferimento
dei suoi elettroni ad un
accettore in modo da
mantenere uno stato
stazionario:
all’ossigeno (fosforilazione
ossidativa)
O ad un’altra
molecola
ossidante
Fermentazione lattica
Fermentazione alcolica
10
30/04/15
Lo scopo della riduzione del piruvato a lattato od
ad etanolo è quello di rigenerare NAD+ in modo
che la glicolisi possa procedere regolarmente
NADH
piruvato
Fegato
rapporto
NADH/NAD+
lattato BASSO
NAD+ + H+
ISOENZIMI
della LATTATO
DEIDROGENASI
MEMBRANA
CELLULARE
permeabile
Muscolo :
rapporto
NADH/NAD+
ALTO
Diverse forme ISOZIMATICHE. La
lattato deidrogenasi (LDH) è una proteina
tetramerica costituito da due diverse
subunità: M ed H
M4, HM3, H2M2, H3M, H4
FORME DIVERSE PER DIVERSE FUNZIONI TESSUTALI
Piruvato
decarbossilasi
ALCOOL
DEIDROGENASI
Anche i tessuti
animali contengono
alcol deidrogenasi.
Alcune delle
principali
conseguenze
metaboliche
dell’intossicazione
da alcool derivano
dall’ossidazione
dell’alcool, nel
fegato,da parte di
questo enzima.
Per prima cosa si ha una massiccia riduzione di NAD+ a
NADH, che riduce il flusso della glicolisi; inoltre l’acetaldeide
è piuttosto tossica e molti degli effetti spiacevoli dei postumi
dell’ubriacatura dipendono dall’azione dell’acetaldeide e dei
suoi metaboliti
11
30/04/15
La rigenerazione del NAD+
che si verifica nella
riduzione del piruvato a
lattato o etanolo sostiene la
continuità del processo
glicolitico in condizioni
anaerobiche
La fermentazione lattica negli organismi superiori
avviene quando la quantità di ossigeno è limitata
rispetto alla richiesta di energia della cellula
Nei vertebrati durante una breve
ma intensa attività muscolare
muscolo
lattato
glucosio
fegato
L’eccesso di ossigeno consumato
durante il periodo di riposo
rappresenta il DEBITO DI
OSSIGENO, quantità di ossigeno
necessaria a fornire ATP per la
gluconeogenesi ( rigenerazione di
glucosio da parte degli epatociti)
12
30/04/15
1) Nella maggior parte dei
tessuti degli animali è
convertito ad acetil-CoA per
essere demolito a CO2 nel ciclo
dell’acido citrico
2) Certi tessuti e tipi di cellule ,
retina ed eritrociti,convertono il
glucosio in lattato anche in
condizioni aerobiche
3) In alcuni tessuti di piante in
alcuni invertebrati, protisti,
microrganismi come il lievito di
birra.
4) Il piruvato può avere
destini anche anabolici
13
30/04/15
Glucosio + 2NAD+ + 2ADP + 2Pi
2 piruvato + 2NADH + 2H+ + 2ATP + 2H2O
Conversione esoergonica del glucosio in piruvato
Glucosio + 2NAD+
2 piruvato + 2NADH + 2H+
ΔG1’o= - 146 KJ/mole
Formazione endoergonica di ATP
2ADP + 2Pi
2ATP + 2H2O
Processo irreversibile portato
a completamento da questa
grande diminuzione di
energia libera
ΔG2’o= 2 (30,5 KJ/mole) = 61 KJ/mole
ΔGs’o= - 85 KJ/mole
Nella glicolisi viene ricavato solo il 5,2%
dell’energia totale del glucosio (2840 KJ/mole)
glicolisi
strettamente
coordinata ad
altre importanti
vie metaboliche
SINTESI DEL GLICOGENO
DEGRADAZIONE DEL GLICOGENO
VIA DEL PENTOSO FOSFATO
GLUCONEOGENESI
CICLO DELL’ACIDO CITRICO
SINTESI DEGLI ACIDI GRASSI
DIFFICILE CONSIDERARE LA GLICOLISI
INDIPENDENTE DALLE ALTRE VIE METABOLICHE
NELLA GLICOLISI DUE SONO GLI ENZIMI CHE HANNO
FUNZIONI REGOLATRICI SUL CATABOLISMO DEL
GLUCOSIO
La FOSFOFRUTTOCHINASI-1 e la PIRUVATO CHINASI
14
30/04/15
enzima
allosterico
Enzima allosterico che ha
come ATTIVATORI : AMP e ADP
nel FEGATO anche il
FRUTTOSIO-2,6-BISFOSFATO
1
6
P O—CH2
5
CH2OH
O
2
HO
4
O P
3
OH
INIBITORI:
ATP
(si lega
ad un sito
distinto
dell’enzima con
affinità inferiore
a quella del sito
attivo).
L’inibizione consiste
in una forte inibizione
dell’affinità
apparente per il
Il CITRATO, intermedio chiave per
fruttosio-6-fosfato.
l’ossidazione aerobica del piruvato.
15
30/04/15
INIBIZIONE DELLA
GLICOLISI DA PARTE
DEL GLUCOSIO
La glicolisi è controllata dall’attività
della fosfofruttochinasi.
Osservazione fatta 1 secolo fa da
Pasteur: Egli vide che quando delle
colture anaerobiche di lievito che
metabolizzavano glucosio venivano
esposte all’aria, la velocità di
reazione diminuiva drasticamente.
Questo effetto ha un preciso
significato metabolico dovuto al
fatto che dall’ossidazione completa
Il flusso metabolico
della fosfofruttochinasi del glucosio si ottiene di gran lunga
viene rallentato in modo più energia che dalla sola glicolisi
specifico dalla presenza
di O2
Il piruvato è punto di
biforcazione nel catabolismo
degli zuccheri e l’enzima che
catalizza la sua produzione è il
secondo enzima che regola la
glicolisi .
Presenta una attivazione a
“feedforward”, inverso della
retroinibizione, da parte del
fruttosio-1,6-bisfosfato.
16
30/04/15
17
30/04/15
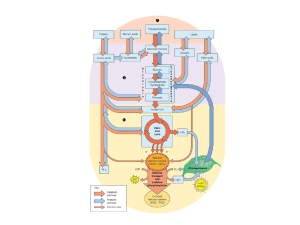
Quadro di insieme
della regolazione
della glicolisi
Per essere riossidato
nella fosforilazione
ossidativa deve
entrare nel
mitocondrio … ma il
NADH non può
attraversare la
membrana
mitocondriale interna
………….. ????????
Sistema
navetta
Diidrossiaceto
ne fosfato/
glicerolo-3fosfato
NADH
Sistemi navetta
per il
trasferimento
degli equivalenti
di riduzione dal
citosol ai
mitocondri
Sistema navetta
malato/aspartato
18
30/04/15
SISTEMI NAVETTA (SHUTTLE)
OSSIDAZIONE DEL NADH CITOSOLICO
Particolarmente
attivo nel fegato
e nel cuore
Particolarmente
attivo nel
cervello e nei
muscoli del volo
degli insetti
19
30/04/15
La forza motrice protonica fornisce energia al trasporto attivo
FAVORITA DAL
GRADIENTE
ELETTRONICO
TRANS-MEMBRANA
FAVORITA DAL
GRADIENTE
PROTONICO
TRANS-MEMBRANA
Molti altri carboidrati
sono disponibili come
fonti di energia
Provengono sia dalla
digestione di alimenti
che da metaboliti
endogeni
20
30/04/15
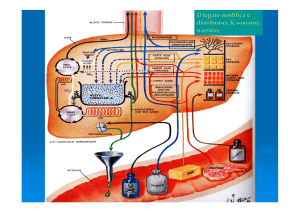
Nell’epatocita
Idrolisi del
saccarosio
nel muscolo e nel
rene dei vertebrati
NEL FEGATO
Esochinasi
ha
un’affinità
20 volte più
alta per il
glucosio
Scavalca la regolazione a livello
della fosfofruttochinasi
spiegando la facilità con cui il
fruttosio proveniente dalla dieta
viene convertito in grasso
21
30/04/15
LA DIGESTIONE DEI GRASSI
NEUTRI (triacilgliceroli) e della
maggior parte dei fosfolipidi
genera GLICEROLO come uno
dei prodotti. Il glicerolo entra
nella glicolisi
22
30/04/15
Digestione dei
polisaccaridi introdotti
con la dieta : amido e SCISSIONE
glicogeno
IDROLISI
Mobilizzazione delle
riserve di glicogeno
proprie dell’animale
FOSFOROLISI
Il glucosio unica fonte energetica usata dal cervello (ad eccezione dei
periodi di digiuno prolungati). Il glicogeno è presente principalmente nel
citosol cellulare del fegato e del muscolo scheletrico, sottoforma di
granuli che contengono proteine regolatorie ed enzimi che ne catalizzano
la sintesi e la degradazione.
Scissione fosforolitica dei legami glicosidici
α(1-4) catalizzata dalla glicogeno fosforilasi
IMPORTANZA DEL
GLUCOSIO-1-FOSFATO
1. In questa fosforolisi parte
dell’energia del legame
glicosidico viene conservata
mediante la formazione del
legame estere fosforico del
glucosio-1-fosfato
Estremità non
riducente
2. Il glucosio-1-fosfato non
può diffondere fuori dalla
cellula il glucosio sì
23
30/04/15
Il glicogeno del muscolo rappresenta una
sorgente rapidamente disponibile per la
cellula. Nel fegato rappresenta una fonte
per l’omeostasi di glucosio del sangue
nell’intervallo tra i pasti. Dopo 12, 18 ore
da un pasto il glicogeno del fegato è
quasi completamente consumato.
La glicogeno fosforilasi agisce sulle
estremità non riducenti delle ramificazioni
del glicogeno
GLICOGENO Un residuo ogni 10 contiene una ramificazione:
legame α(1-6) glicosidico
La glicogeno fosforilasi agisce ripetitivamente
sulle estremità non riducenti delle
ramificazioni del glicogeno fino a 4 residui di
glucosio dall’inizio di una ramificazione
L’attività transferasica dell’enzima sposta, da
prima un blocco di tre residui dalla
ramificazione all’estremità non riducente
vicina legandola con legame glicosidico
(α
1-4), poi taglia l’ultimo residuo rimasto sulla
ramificazione con attività glicosidica (α 1-6).
Le due attività, trasferasica e glicosidica, sono
attività separate di un’unica proteina
enzimatica: ENZIMA DERAMIFICANTE
Il glucosio-1-fosfato è poi convertito a
glucosio-6-fosfato per entrare nel flusso
metabolico principale (Es.:GLICOLISI)
24
30/04/15
Glucosio 1-fosfato
fosfoglucomutasi
glucosio 6-fosfato
La MOBILIZZAZIONE DEL GLICOGENO così come la
GLUCONEOGENESI producono forme fosforilate che non
possono uscire dalla cellula epatica. La conversione in
glucosio libero comporta l’azione della Glucosio-6fosfatasi enzima presentenel fegato,nel rene e
nell’intestino. Al contrario il glicogeno muscolare serve
per il CATABOLISMO della cellula per cui il muscolo non
possiede questo enzima.
Nel fegato (e nei reni), ma non nel muscolo
Glucosio-6-P
Glucosio-6-fosfatasi
glucosio + Pi
può uscire dalla cellula nel sangue
La mobilizzazione
del glicogeno è
controllata da
ORMONI attraverso
una cascata
metabolica che è
attivata dalla
formazione di
cAMP e prevede di
FOSFORILAZIONI
SUCCESSIVE di
proteine
enzimatiche
25
30/04/15
Molte sono le vie che confluiscono o
che partono dal glucosio-6-fosfato
GLICOGENO
GLUCOSIO LIBERO
Glucosio-6-fosfato
GLICOLISI
VIA DEL PENTOSO
FOSFATO
GLUCONEOGENESI
26
30/04/15
Produce NADPH e ribosio 5-fosfato
GPD
idrolasi
Formazione di NADPH
per la sintesi di acidi
grassi e degli steroli
Biosintesi dei
nucleotidi e
degli acidi
nucleici
Glucosio 6-fosfato + 2NADP+ + H2O
ribosio 5-fosfato + CO2 + 2 NADPH + 2H+
PGD
isomerasi
Il ruolo di questa via è
principalmente anabolico ( i
prodotti servono per l’anabolismo)
piuttosto che catabolico, sebbene
compor ti l’ossidazione del
glucosio
Nei tessuti che richiedono principalmente, od in un certo
momento NADPH , i pentoso fosfati vengono riciclati a
glucosio 6-fosfato mediante una serie di reazioni
isomerasi
tK
tA
tK
GPD
PGD
epimerasi
Tutte
le
reazi
oni
sono
rever
sibili
idrolasi
27
30/04/15
Formazione
di R5P a
partire da
intermedi
della
glicolisi
VIE
ALTERNATIVE
DEI
PENTOSO FOSFATI
28
30/04/15
NADPH è utilizzato in reazioni che richiedono un
donatore di elettroni con elevato potere riducente
Specie reattive dell’ossigeno
Reazioni antiossidanti
Nei globuli rossi la via dei pentoso fosfati fornisce
NADPH per la riduzione del glutatione ossidato
Le cellule rigenerano glutatione
ridotto attraverso una reazione
catalizzata dalla glutatione
reduttasi che utilizza NADPH
come fonte di elettroni riducenti
I globuli rossi dipendono
esclusivamente dalla via dei
pentosi fosfati per la
produzione di NADPH
29
30/04/15
In individui con
carenza di G6PD
Farmaci ossidativi
Antibiotici
Antimalarici
Antipiretici
FAVISMO
Effetto emolitico
innescato da
ingestione di fave
Il gene per questo enzima si trova sul cromosama x
Il difetto si manifesta con emolisi dei globuli rossi quando individui
sensibili sono sottoposti a stress ossidativo causato da farmaci
antiossidativi o per l’ingestione di fave ( Vicia faba, FAVISMO)
300 mutazioni che spiegano
numerose varianti del gene
G6PD A- e G6PD mediterranea
differiscono dalla condizione
normale per un solo aa
30
30/04/15
31