B
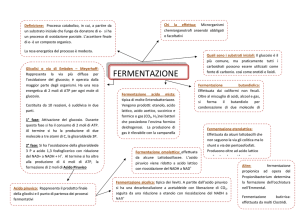
La fermentazione
Nella fermentazione non si ha la completa demolizione del composto di partenza,
che viene solo parzialmente fermentato in uno o più prodotti finali per lo più ancora
organici, che mantengono ancora parte dell’energia del composto iniziale. Pertanto
la resa energetica della fermentazione non è paragonabile a quella dei processi
respiratori e l’ATP viene generato soltanto per trasferimento di un gruppo fosfato
legato ad un intermedio del processo con un legame ad alto contenuto energetico
all’ADP (fosforilazione a livello di substrato)
Rese teoriche in ATP per molecola di glucosio
RESPIRAZIONE: 36-38 molecole teoriche di ATP (2 da glicolisi, 2 dal ciclo di Krebs
(GTP) e 32-34 dalla catena di trasporto degli elettroni)
FERMENTAZIONE ALCOLICA e LATTICA: 2 ATP
FERMENTAZIONE dei BIFIDOBATTERI: 2,5 ATP
Il metabolismo respiratorio è circa 15 volte più efficiente del metabolismo fermentativo
Il metabolismo fermentativo, però, è molto rapido!
1
B
La fermentazione
La glicolisi è la principale via che precede numerose fermentazioni. Per la maggior parte dei
processi fermentativi la stessa glicolisi è anche la via che frutta alle cellule l’energia, scopo
del processo metabolico (2 moli di ATP per mole di glucosio fermentato) attraverso le già citate
reazioni di fosforilazione a livello di substrato. Le tappe successive della fermentazione hanno
invece lo scopo di consentire la riossidazione delle due moli di NADH, prodotte nell’ossidazione
della 3-fosfo gliceraldeide ad acido 1,3-difosfoglicerico, in quanto nei microrganismi anaerobi la
catena di trasporto degli elettroni è incompleta. Pertanto l’equilibrio ossido-riduttivo si ristabilisce
mediante riossidazione del NADH a spese di un intermedio del processo a valle dell’acido piruvico.
In qualche caso, in questo tratto della fermentazione si ha ulteriore produzione di ATP, che porta la
resa energetica della fermentazione a valori comunque di ben poco superiori a 2
Quasi sempre le varie vie fermentative sono caratteristiche di ben definiti gruppi microbici,
i quali dimostrano un’elevata specificità di processo
2
La fermentazione omolattica
B1
È operata dai batteri omolattici
od omofermentanti, anaerobi
obbligati ossigeno-tolleranti
appartenenti ai generi Gram
positivi Streptococcus,
Pediococcus, Lactobacillus…
In questa fermentazione è lo
stesso acido piruvico di origine
glicolitica a fungere da
accettore di elettroni per la
riossidazione del NADH,
riducendosi così ad acido
lattico
lattato deidrogenasi
da ricordare!
3
La fermentazione alcolica
B2
È caratteristicaFERMENTAZIONE
dei lieviti, che sono
eumiceti unicellulari anaerobi facoltativi di cui è
ALCOOLICA
ben noto il genere Saccharomyces. L’enzima-chiave di questa fermentazione è la
piruvato decarbossilasi
glucosio
2ATP
2 gliceraldeide-3- P
2 etanolo
NAD+
NADH
2 acetaldeide
2 CO2
da ricordare!
NAD +
NADH
2 piruvato
4ATP
Avviene anche nel già
citato batterio Zymomonas
Equazione generale:
glucosio
Resa energetica = 2 ATP
2 etanolo + 2 CO2
4
La fermentazione acido-mista
B3
glucosio
2NADH
NADH
succinato
ossaloacetato
2ATP
CO2
fosfoenolpiruvato (PEP)
2ATP
NADH
piruvato
lattato
CoA
NADH
acetaldeide
NADH
acetil-CoA
formiato
etanolo
acetil- P
CO2
ATP
H2
formico
idrogenoliasi
acetato
Gli enterobatteri (cioè i membri della famiglia
Escherichia,
Enterobacteriaceae)
come
Salmonella, Shigella e i generi Proteus, Vibrio e
Photobacterium sono microrganismi Gramnegativi anaerobi facoltativi che in anaerobiosi
fermentano il glucosio secondo questa via. I
diversi prodotti finali caratteristici di questo
provesso sono acidi organici a basso peso
molecolare quali acido formico, acido acetico,
acido lattico e acido succinico e piccole quantità
di etanolo (in E. coli e Proteus l’acido formico è
scisso in CO2 e H2 dalla formico idrogenoliasi,
un enzima la cui presenza assume una
rilevante
importanza
tassonomica
e
diagnostica)
I rapporti percentuali di questi prodotti variano
nei diversi microrganismi e a seconda delle
condizioni colturali e ambientali, sempre nel
rispetto dell’equilibrio ossido-riduttivo del
processo
5
FERMENTAZIONE
2.3-BUTILENGLICOL
B4
La fermentazione 2,3-butilenglicolica
glucosio
2 NADH
NADH
lattato
2 ATP
piruvato
NADH
acetaldeide
NADH
formiato
acetil-CoA
4
etanolo
CO2
CO2
α-acetolattato
CO2
acetoina
acetoino
NADH
2,32,3-butilenglicol
butilenglicole
o butandiolo
H2
Tipica di altre Enterobacteriaceae aventi come
habitat il suolo o le acque (Enterobacter
aerogenes e i generi Erwinia, e Serratia, patogeni
delle piante). Il prodotto principale è il 2,3butilenglicol che si forma dalla condensazione di
due molecole di piruvato con la liberazione di due
molecole di CO2. In minore quantità si producono
anche gli acidi della precedente fermentazione
accanto a notevoli quantità di etanolo per il
mantenimento dell’equilibrio ossido-riduttivo.
L’uso di questa via causa il decremento della
formazione di acidi (il butilenglicole è neutro) e la
formazione dell’intermedio ACETOINO. Coloro
che si occupano di analisi microbiologica delle
acque possono distinguere i coliformi fecali
(fermentazione acidomista) dai non fecali
(fermentazione butileglicolica tipica dei generi
come Klebsiella) andando a svelare la presenza
di acetoino ed un pH più alto
6
La fermentazione propionica
B5
acido assalacetico
acido
malico
acido lattico
Il glucosio viene fermentato ad
acido acetico, CO2 ed acido
propionico
dal
genere
Propionibacterium attraverso una
via
complessa
detta
del
metilmalonilCoA. Veillonella, il cui
habitat è il rumine dei bovini,
compie la stessa fermentazione
partendo dall’acido lattico. Altri
microrganismi producono acido
propionico
da
acido
lattico
attraverso una via diversa
N.B.: La resa energetica, tra le
più elevate, include anche una
mole di ATP prodotto
per
fosforilazione
ossidativa
(cioè
attraverso la ATP sintasi) tramite
una breve catena di trasporto degli
elettroni
nella
riduzione
del
fumarato a succinato (respirazione
del fumarato: è un tipo di
respirazione anaerobica)
acido propionico
Resa energetica = 5 ATP / 1,5 glucosio o 2 ATP / 3 ac. lattico
7
FERMENTAZIONE BUTIRRICA
La fermentazione butirrica
B6
CH3 - CH2 - CH2 - COOH
butirrato
Si ritrova in Clostridium, sporigeno
anaerobio
obbligato
sensibile
all’ossigeno, in grado di fermentare il
glucosio. Caratteristica di questa via è la
produzione di CO2 e H2 nella reazione
del coenzima A (CoASH) con il piruvato.
Il principale prodotto finale è l’acido
butirrico, la cui formazione consente il
mantenimento dell’equilibrio ossidoriduttivo
ATP
CH3 - CH2 - CH2 - CO - O - P
butirril fosfato
glucosio
Ferredossina
CoA
Pi
2NADH
CH3 - CH2 - CH2 - CO - CoA
butirril-CoA
2H2
2CH3 - CO - COOH
2FdOX
NADH
CH3 - CH CH - CO - CoA
crotonil-CoA
Ferredossina
idrogenasi
2FdRED
2CoA
2CH3 - CO - CoA
CoA
CH3 - CH - CH2 - CO - CoA
CH3 - CO - CH2 - CO - CoA
acetoacetil-CoA
OH
L (+) -β-idrossibutirril-CoA
NADH
Resa: 3 ATP
2CO2
2H+
H2O
Equazione generale: glucosio
2ATP
butirrato + 2CO2 + 2H2
Alcuni Clostridi producono in particolari
condizioni butanolo e acetone, che sono
di interesse industriale
N.B.: la ferredossina è una ferro-zolfo proteina
che media il trasferimento di elettroni in diverse
reazioni metaboliche
8
B7
La fermentazione omoacetica
In alcuni clostridi, il glucosio (1 mole) viene stechiometricamente fermentato a 3 moli
di acetato, una delle quali deriva dalla riduzione delle 2 moli di CO2, che provengono
dalla scissione del piruvato ad acetil-CoA
Non va confusa con la produzione di acido acetico da parte di Acetobacter
e Gluconobaceter, i quali, per produrre acido acetico, non operano una
fermentazione, ma ossidano l’etanolo ad acido acetico per recuperare
potere riducente (NADH + H+)
4
B8
9
Il metabolismo fermentativo dei bifidobatteri
Ha scarsa attività
nei bifidobatteri
È assente nei
bifidobatteri
Aldolasi
Il fruttosio 1,6 bisfosfato
funge da induttore della
lattato deidrogenasi
10
4
Fruttosio-6-P
Pi
B8
H2O
F6PPK (xfp)
D-eritrosio-4-P
Acetil-P
ADP
1
Diidrossiaceton
transferasi
ATP
Fruttosio-6-P
Acetato
Via fermetativa di Bifidobacterium
o
via della Fruttosio-6-P Fosfochetolasi
D-sedoeptulosio-7-P
D-gliceraldeide-3-P
D-xilulosio-5P
D-ribosio-5-P
D-ribulosio-5-P
D-xilulosio-5-P
Pi
2
H2O
F6PPK (xfp)
2 Acetil-P
ADP
2
ATP
Acetate
2 D-gliceraldeide-3-P
ACETATO:LATTATO = 3:2
Formiato
Etanolo
11
Legenda per la descrizione tassonomica
p__ = phylum, cioè divisione
c__ = classe
o__ = ordine
f__ = famiglia
g__ = genere
Per esempio, per descrivere Escherichia coli, scriveremo:
p__Proteobacteria;c__Gammaproteobacteria;o__Enterobacteriales;
f__Enterobacteriaceae;g__Escherichia
…oppure, per descrivere Bacillus licheniformis:
p__Firmicutes;c__Bacilli;o__Bacillales;f__Bacillaceae;g__Bacillus
12
I BATTERI LATTICI
Sono batteri Gram POSITIVI, immobili, chemioeterotrofi a forma di bastoncino o cocco
Il loro nome deriva dal fatto che formano ACIDO LATTICO come principale (e talvolta unico)
prodotto finale del loro metabolismo energetico
Hanno un metabolismo fermentativo; sono tutti anaerobi con variabile capacità di
tollerare l’ossigeno (con rare eccezioni, legate alla presenza di EME esogeno)
Sono incapaci di produrre ATP attraverso un metabolismo aerobio, a causa dell’incapacità di
sintetizzare citocromi o altri enzimi contenenti il gruppo eme. A causa dell’impossibilità di
produrre proteine eminiche, sono CATALASI NEGATIVI e quindi non possono operare la
decomposizione dell’H2O2 in H2O e O2
Una caratteristica propria dei batteri lattici è la loro elevata ACIDO-RESISTENZA, la quale
consente loro di crescere fino a quando il pH raggiunge valori inferiori a 5. Questa
caratteristica fisiologica è di grande importanza ecologica, perché permette loro di vincere la
competizione di altri batteri in ambienti ricchi di materia organica. Come risultato della loro
specializzazione fisiologica, i batteri lattici sono confinati in pochi e caratteristici ambienti
naturali: alcuni vivono in associazione con le piante e crescono a spese delle sostanze nutritive
liberate in seguito alla morte e alla decomposizione dei tessuti vegetali; si trovano quindi in
alimenti e bevande preparate con materiale vegetale come sottaceti, crauti, foraggi insilati, vino
e birra. Altri fanno parte del normale microbiota degli animali (bocca, intestino, mucosa
vaginale). Infine, alcuni sono spesso associati al latte (per es. Streptococcus thermophilus).
13
Alcuni generi di batteri lattici
Streptococcus
Oenococcus
Lactococcus
Enterococcus
Carnobacterium
Pediococcus
Lactobacillus
Leuconostoc
Immagini al SEM
p__Firmicutes;c__Bacilli;o__Lactobacillales;f__Lactobacillaceae;g__Lactobacillus
p__Firmicutes;c__Bacilli;o__Lactobacillales;f__Lactobacillaceae;g__Pediococcus
p__Firmicutes;c__Bacilli;o__Lactobacillales;f__Streptococcaceae;g__Lactococcus
p__Firmicutes;c__Bacilli;o__Lactobacillales;f__Streptococcaceae;g__Streptococcus
p__Firmicutes;c__Bacilli;o__Lactobacillales;f__Leuconostocaceae;g__Oenococcus
p__Firmicutes;c__Bacilli;o__Lactobacillales;f__Leuconostocaceae;g__Leuconostoc
p__Firmicutes;c__Bacilli;o__Lactobacillales;f__ Enterococcaceae;g__Enterococcus
p__Firmicutes;c__Bacilli;o__Lactobacillales;f__Carnobacteriaceae;g__Carnobacterium
14
I BATTERI LATTICI
Sulla base dei loro prodotti di fermentazione, sono divisi in tre sottogruppi metabolici
1 – OMOFERMENTANTI: degradano il glucosio via glicolisi e non sono in grado di
utilizzare i pentosi. Il piruvato viene poi ridotto ad acido lattico, unico prodotto della
loro fermentazione
2 – ETEROFERMENTANTI OBBLIGATI: non possono degradare il glucosio via
glicolisi in quanto non possiedono l’enzima FRUTTOSIO 1,6 DIFOSFATO
ALDOLASI che catalizza la scissione del fruttosio 1,6 difosfato in gliceraldeide-3fosfato e diidrossi-acetone-fosfato. Per tale ragione essi fermentano il glucosio
attraverso la via dei pentoso fosfati, ottenendo tre prodotti finali in rapporto
equimolare: ACIDO LATTICO, ETANOLO e CO2. Possono utilizzare anche i pentosi,
nel qual caso senza produzione di CO2. Possono produrre anche piccole quantità di
acido acetico
3 – ETEROFERMENTANTI FACOLTATIVI: in presenza di ESOSI essi li fermentano
attraverso la glicolisi con produzione di solo ACIDO LATTICO, comportandosi perciò
come omofermentanti. In presenza di PENTOSI, effettuano l’ossidazione di tali
composti attraverso la via dei pentoso fosfati, senza produzione di CO2
L’acido lattico prodotto può essere in configurazione L, D oppure in miscela
racemica DL
15
Oenococcus
16
Omofermentanti
Zuccheri
Zuccheri pentosi
esosi
X
Esosi
Eterofermentanti
facoltativi
Eterofermentanti
obbligati
esosi
Pentosi
in quantità ridotte
(
)
17
TERRENI COLTURALI PER BATTERI LATTICI
Terreno MRS a pH 6,2-7: non selettivo, adatto per il mantenimento dei
lattobacilli
Terreno SL: selettivo per l’isolamento dei lattobacilli (tripticase,
estratto di lievito, citrato, glucosio, arabinosio, saccarosio, acetato, pH
finale 5,4)
Terreno ATB: adatto per l’isolamento di Oenococcus oeni (peptone,
estratto di lievito, succo di pomodoro 25 %, pH finale 4,8)
Poiché i batteri lattici sono anaerobi (sebbene ossigeno tolleranti), i migliori
risultati di crescita si ottengono incubando le piastre di isolamento in giare di
anaerobiosi (Gas-Pak)
18
I BATTERI ACETICI
Appartengono alla famiglia delle Acetobacteraceae
p__Proteobacteria;c__Alphaproteobacteria;o__Rhodobacterales;f__Acetobacteraceae
Sono bastoncini Gram NEGATIVI, aerobi chemio-eterotrofi, catalasi positivi,
termolabili e psicrotrofi
Sono marcatamente acidofili (crescono fino a pH 4, a concentrazioni di ac. acetico fino a 0,4 M)
Il pH ottimale è compreso tra 5,4 e 6,3. Acetobacter pasteurianus cresce anche a pH 3 se
l’etanolo è inferiore all’8%
Si localizzano soprattutto sulla superficie di piante, in particolare fiori e frutti
Sono microrganismi mesofili che crescono con temperatura ottimale intorno a 30 °C; alcuni
ceppi si riescono a sviluppare anche a 5 °C, altri sono blandamente termotolleranti (37-40 °C)
Tollerano basse concentrazioni di acido acetico; tollerano bene l’etanolo e molti ceppi
crescono in un range compreso tra 8 e 15 %. La concentrazione tollerata dipende da
pH, temperatura e concertazione di O2
I tre generi più noti:
Acetobacter: A. xylinus, A. aceti, A. pasteurianus, A. cerevisiae, A. pomorum
Gluconoacetobacter: G. xylinus, G. diazotrophicus, G. intermedius, G. liquefaciens
Gluconobacter: G. oxydans, G. frateurii
Altri generi e specie: Acidomonas methanolica, Asaia spp., Kozakia baliensis,
Saccharibacter spp., Swaminathania spp., Frateuria spp.
19
Le specie più importanti nella produzione industriale dell’aceto sono:
- Acetobacter aceti subsp. orleanensis
- Acetobacter aceti subsp. xylinum (madre dell’aceto)
- Acetobacter pasteurianus subsp. lovaniensis
- Acetobacter pasteurianus subsp. ascendens
- Acetobacter pasteurianus subsp. paradoxus
In particolare, i membri del genere Acetobacter sono particolarmente importati da un punto di
vista industriale (sia positivamente che negativamente) per diversi motivi, tra i quali:
1. sono usati per la produzione dell’aceto (usati per convertire intenzionalmente l’etanolo del
vino o di altri prodotti alcolici in acido acetico)
2. possono deteriorare il vino producendo quantità eccessive di acido acetico o etil-acetato
(entrambi rendono sgradevole il vino). La crescita di Acetobacter nel vino può essere inibita
attraverso la completa eliminazione dell’ossigeno durante la conservazione del vino oppure
con l’aggiunta di moderate quantità di SO2
3. sono usati per acidificare intenzionalmente alcune birre
aventi periodi di maturazione lunghi (per esempio per
la produzione della birra tradizionale Flemish Sour Ale)
20
Peculiarità metaboliche
I batteri acetici sono aerobi obbligati aventi metabolismo aerobio (respirazione
aerobica)
In particolare, i batteri acetici sono caratterizzati da metabolismo aerobio con
ossidazione incompleta dei substrati organici
Utilizzano diverse fonti di carbonio, dagli zuccheri agli aminoacidi. Preferiscono
etanolo, glicerolo, lattato, mannitolo, sorbitolo, fruttosio, glucosio
Come fonte di azoto molti ceppi utilizzano NH4+
Gli zuccheri sono ossidati esclusivamente attraverso la via del pentosio fosfato
L’acetato deriva dalla ossidazione parziale della materia organica e non da un vero
processo di fermentazione
Il PIRUVATO è trasformato attraverso una decarbossilazione non ossidativa ad
acetaldeide (la decarbossilazione ossidativa del piruvato ad Acetil-CoA e CO2,
tipicamente presente nel processo di respirazione, non avviene in quanto il
COMPLESSO DELLA PIRUVATO DEIDROGENASI nei batteri acetici è inattivo)
FONDAMENTALE!!! DA RICORDARE!!!
I BATTERI ACETICI
Glucosio
tutto il NADH generato in queste reazioni
di ossidazione è destinato alla catena di
trasporto degli elettroni, per creare il
gradiente di H+ che permette la sintesi
dell’ATP
NAD+
ACIDO LATTICO
ETANOLO
via del pentosio fosfato
pentosi-P
interconversione non ossidativa
triosi-P
ultime tappe della glicolisi
NADH+H+
Ac. piruvico
lattato deidrogenasi
NAD+
NADH+H+
alcol deidrogenasi
decarbossilazione non ossidativa
CO2
Acetaldeide
NAD+
NADH+H+
acetil-CoA ligasi
acetil-CoA sintetasi
Gluconobacter
non ha un ciclo di Krebs
funzionale e non può
dunque ossidare l’acetato
21
aldeide deidrogenasi
Ac. acetico
Acetil-CoA
Ciclo dei
TCA
CO2
CO2
In Acetobacter
il ciclo di Krebs è
inibito solo a
concentrazioni
elevate di etanolo
22
I BATTERI ACETICI: la metabolizzazione dell’etanolo
Sono capaci di crescere utilizzando etanolo come unica fonte di carbonio ed
energia producendo acido acetico
L’etanolo, infatti, viene ossidato ad acetato, ed il potere riducente ottenuto si trasforma
in energia per la cellula (cioè in ATP) a livello della catena di trasporto degli elettroni
CH 3CH 2OH
pirrolochinolin-chinone, associato
all’aldeide deidrogenasi
biosintesi
NAD +, PQQ
catena respiratoria
NADH, PQQH 2
ATP
CH 3COOH
KREBS
CH 3COSCoA
acetil-CoA sintetasi
rilasciato extracellularmente
detta anche
acetil-CoA ligasi
X
Il blocco del ciclo di Krebs è causato da:
→ presenza di etanolo in Acetobacter
→ carenze genetiche in Gluconobacter
23
I BATTERI ACETICI:
la metabolizzazione dell’etanolo
La produzione di acido acetico a partire da
etanolo da parte dei batteri acetici NON è
una fermentazione, ma il risultato di una
semplice ossidazione parziale dell’etanolo,
al fine di recuperare potere riducente
Solo i membri del genere
Acetobacter possono ossidare
completamente l’acetato a CO2,
se le concentrazioni di etanolo
non sono troppo alte
24
Come distinguere Acetobacter da Gluconobacter in laboratorio
Si devono preparare piastre contenenti il seguente terreno di coltura agarizzato:
Estratto di lievito
50 g/l
CaCO3
20 g/l
rende il terreno di coltura opaco
Etanolo
7%
è la fonte di carbonio ed energia per il batterio
Agar
20 g/l
Dopo semina e incubazione a 28/30 °C per 2-3 giorni, parte dell’etanolo sarà convertito dai batteri
acetici in acido acetico, che abbasserà il pH solubilizzando il carbonato di calcio: le colonie
appariranno quindi con un alone di chiarificazione intorno a loro. Prolungando l’incubazione la
concentrazione di etanolo diminuirà fino a permettere agli Acetobacter di ossidare completamente
l’acido acetico prodotto (poiché il ciclo di Krebs è sbloccato).
Dopo altri 2-3 di incubazione, quindi, il
Acetobacter
pH sarà risalito, con conseguente riprecipitazione del carbonato di calcio.
Dopo un totale di 5-6 giorni di
incubazione, quindi, le colonie di2-3 giorni di
Gluconobacter avranno un alone diincubazione
chiarificazione, mentre quelle di
Acetobacter no, poiché il carbonato di5-6 giorni di
calcio riprecipitando avrà reso diincubazione
nuovo opaco il terreno (sì vedrà un
alone opaco di precipitazione).
Gluconobacter
colonia batterica
alone di chiarificazione
alone di precipitazione
25
La fermentazione malolattica
(o meglio, conversione malolattica)
NON è una fermentazione nel senso
metabolicamente corretto del termine
26
CENNI DI ANABOLISMO
L'anabolismo comprende tutto
l'insieme dei processi di sintesi
o bioformazione delle molecole
organiche (biomolecole) più
complesse da quelle più
semplici o dalle sostanze
nutritive.
Questi
processi
richiedono energia, al contrario
del catabolismo
Sebbene l'anabolismo e il
catabolismo siano due processi
contrari, funzionano in maniera
coordinata ed interdipendente
Le vie cataboliche forniscono gli
intermedi
biochimici
utili
all’anabolismo per operare la
biosintesi delle macromolecole
27
I PRECURSORI BIOSINTETICI DEGLI AMMINOACIDI
28
SINTESI DELLE STRUTTURE CELLULARI A PARTIRE DA GLUCOSIO
I precursori metabolici formati
dalla glicolisi, dal ciclo dei TCA
e dalle vie correlate entrano
nelle vie biosintetiche che
producono…
…i monomeri o i
mattoni di tutte le
componenti cellulari.
PROTOTROFIA
e
AUXOTROFIA
INCAPACITA’ DI SINTETIZZARE UN ...
amminoacido ... una base azotata ... etc...
Ulteriori reazioni
polimerizzano i monomeri
per formare
macromolecole…
…che interagiscono per
produrre le strutture che
costituiscono la cellula
batterica
CAPACITÀ DI SINTETIZZARE... un
amminoacido... una base azotata... etc…
FABBISOGNO COLTURALE,
NUTRIZIONALE
29
LA BIOSINTESI DELLE BASI AZOTATE
30