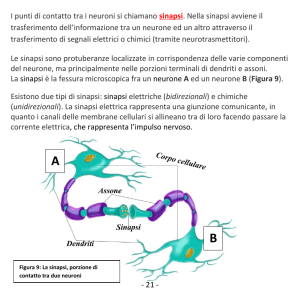
La plasticità neurale al livello della sinapsi fibra parallelacellula di Purkinje (PF-PC)
Le cellule di Purkinje costituiscono il cosiddetto
strato piriforme della corteccia del cervelletto. I loro
assoni decorrono inferiormente nello strato granulare
mentre i loro dendriti, che costituiscono
l’arborizzazione più estesa di tutto il SNC, dipartono a
ventaglio nello strato molecolare sovrastante.
Qui, le cellule di Purkinje, prendono contatto con le
Fibre Parallele, biforcazioni a T dei numerosi assoni
delle cellule dei Granuli, le quali una volta ascese
nello strato molecolare dalle Fibre Muscoidi,
divergono longitudinalmente in due direzioni opposte.
L’esteso albero dendritico è invece contattato da una
singola fibra rampicante proveniente dal nucleo
olivare inferiore. Sia le fibre parallele che le fibre
rampicanti forniscono input eccitatori alle cellule di
Purkinje, le quali inviano proiezioni inibitorie ai
nuclei cerebellari profondi. L’insieme di queste
strutture consente l’espressione di quella complessa
attività elettrofisiologiche GABAergica delle cellule
di Purkinje alla base della regolazione dei movimenti
complessi e coordinati.
La sinapsi fibra parallela-cellula di
Purkinje (PF-PC) rappresenta il punto
anatomico principale di questa attività
definibile
come
neuromodulazione
cerebellare. E’ stato stimato come, nel
cervelletto di ratto, ogni fibra parallela
prenda contatto con almeno 300 cellule
di Purkinje diverse, durante il suo
decorso
trasversale
nello
strato
molecolare. Per questo, un singolo
segnale
proveniente
dalla
fibra
muscoide, prima attraverso le cellule dei
granuli e poi attraverso le fibre parallele,
può essere distribuito in 150.000 cellule
di Purkinje, le quali, oltretutto, fanno
sinapsi con più di una fibra. I contatti
sinaptici PF-PC stimati nel cervelletto di
ratto sono 60 x 109, nell’uomo 1014.
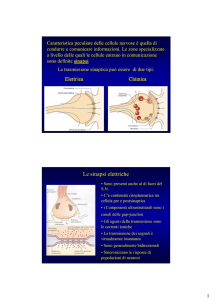
La sinapsi PF-PC è sede dei meccanismi di plasticità funzionale come la depressione a lungo
termine (LTD) e il potenziamento a lungo termine pre-sinaptico e post-sinaptico (LTP). Si definisce
plasticità sinaptica, la capacità del sistema nervoso di modificare l'intensità delle relazioni
interneuronali agendo sulla struttura e la funzionalità delle stesse sinapsi.
L’LTD è il risultato della
contemporanea stimolazione della
cellula di Purkinje da parte della
fibra parallela e della fibra
rampicante.
Il
rilascio
di
glutammato nella sinapsi PF-PC e
l’attivazione dei canali Ca2+
voltaggio/dipendenti indotta dalla
CF, agiscono simultaneamente
determinando un notevole aumento
di calcio nella cellula di Purkinje.
Una
tale
concentrazione
intracellulare dello ione comporta
l’attivazione di una protein-chinasi
(PKC), che, fosforilando i recettori
AMPA,
ne
determina
l’internalizzazione e la successiva
eliminazione dalla membrana. Il
risultato
è
una
riduzione
dell’intensità della trasmissione
sinaptica tra la fibra parallela e la
cellula
di
Purkinje
e
una
conseguente diminuzione della
capacità eccitatoria della cellula.
L’LTP coinvolge invece la sola sinapsi PF-PC ed è il risultato di cambiamenti nel rilascio o nella
capacità della cellula di Purkinje di rispondere al neurotrasmettitore.
La modulazione presinaptica, presuppone la stimolazione tetanica ad alta frequenza nella fibra
parallela che determina l’apertura di canali specifici per il calcio nel terminale assonico.
L’accumulo dello ione consente l’attivazione di proteine specifiche che incrementano il rilascio di
vescicole contenenti Glutammato nello spazio sinaptico. L’intensità di rilascio del
neurotrasmettitore è associata ad una maggiore eccitabilità della cellula di Purkinje.
La modulazione postsinaptica è invece frutto di cambiamenti morfologici nella membrana
plasmatica della stessa cellula di Purkinje e determinata dalla bassa frequenza di scarica da parte
della fibra parallela. L’aumento dell’eccitabilità della cellula è la conseguenza dell’incremento nel
numero di recettori AMPA in membrana, favorito dalla scarsa concentrazione di calcio
intracellulare che segue la lieve stimolazione.
Gli studi hanno rivelato come la plasticità sinaptica che coinvolge la sinapsi PF-PC sia la proprietà
neurobiologica che non entra in gioco unicamente in caso di necessità, come nella riparazione della
connessione neuronale in seguito a danni del SNC, ma si ritiene sia alla base degli eventi di
apprendimento motorio e del fenomeno della memoria.
Per apprendimento motorio si intende l’acquisizione di adeguamenti involontari come l’adattamento
al riflesso vestibulo-oculare (VOR), il battito ciliare condizionato da uno stimolo uditivo o visivo e
l’apprendimento di nuovi riflessi posturali in seguito alla perdita improvvisa di equilibrio.
E’ stato osservato, infatti, come i cambiamenti associati all’LTD avvengano proprio durante la
continua correzione operata dal SNC nel fissare nella retina l’immagine di un oggetto che si muove
in direzione opposta al capo durante il VOR. Pertanto, i segnali convogliati dalle fibre rampicanti
modificherebbero, in periodi di tempo lunghi, le risposte dei neuroni di Purkinje rispetto ai segnali
che arrivano attraverso le fibre parallele.
Inoltre, studi su cellule di Purkinje di coniglio addestrato ad associare il riflesso ciliare ad uno
stimolo particolare, mostrano come l’eccitabilità della membrana cellulare persista anche dopo
trenta giorni dall’esercizio, rivelando l’acquisizione di cambiamenti a lungo termine a favore della
trasmissione sinaptica (LTP). I circuiti cerebellari vengono perciò modificati a lungo temine con
l’esperienza. Anche l’adeguamento ad una nuova postura in seguito alla perdita di equilibrio
sarebbe determinato da cambiamenti fini al livello della sinapsi PF-PC.
Per questi motivi, l’alterata funzionalità della sinapsi PF-PC è stata osservata in disordini
neurologici e motori alla base di alcune malattie umane, come l’atassia spinocerebellare. Inoltre,
negli ultimi anni alcuni studi stanno indagando il suo ruolo in alcuni disordini psichiatrici, come
l’autismo infantile.
Le atassie cerebellari sono malattie
degenerative progressive che si verificano
negli adulti sporadicamente o possono essere
ereditarie. I sintomi includono perdita di
coordinazione di muscoli volontari, comparsa
di dismetria, perdita di equilibrio e problemi
di linguaggio. Tuttavia, entrambi i casi di
atassia cerebellare, sporadici ed ereditari,
presentano caratteristiche fisiopatologiche
comuni come la degenerazione specifica dei
principali neuroni cerebellari, le cellule di
Purkinje.
L'Atassia spinocerebellare di tipo 1 (SCA1) è associata ad una espansione genica della tripletta
codificante per la glutammina che causa l’aggiunta soprannumeraria dell’amminoacido nella
proteina ataxina-1. La mutazione è associata alla deregolazione nell’espressione di molti geni
compreso il recettore per il glutammato (mGlu1), presente nella membrana plasmatica delle cellule
di Purkinje. In particolare il recettore metabotropico subisce una riduzione nella sua espressione
provocando deficit al livello della sinapsi, essendo la cellula di Purkinje non più in grado di
rispondere al neurotrasmettitore. E’ stato dimostrato come il trattamento di topi affetti da atassia
spinocerebellare di tipo 1 con modulatori allosterici del recettore, porti ad un miglioramento a lungo
termine delle performance motorie. L’utilizzo del modulatore positivo PAM migliora l’attività del
recettore compensando la sua limitata quantità sulla membrana plasmatica della cellula di Purkinje.
Al contrario, il trattamento di PC con antagonisti del recettore, riduce drasticamente la performance
motoria del topo transgenico per la malattia. Per questi motivi l’atassia di tipo 1 è stata associata ad
un deficit del recettore del glutammato, la cui funzionalità è necessaria per la trasmissione sinaptica.
Anche nell'Atassia spinocerebellare di tipo 5 (SCA5) si nota una riduzione del recettore nelle
spine dendritiche delle cellule di Purkinje. Tuttavia questa volta la riduzione è derivante dalla
mutazione della proteina spettrina, la cui funzione in queste cellule è quella di stabilizzare il
recettore del glutammato sulla superficie della membrana plasmatica.
L’Atassia spinocerebellare di tipo 3 (SCA-3) è associata ad una amplificazione genica della
tripletta codificante per la glutammina nel gene ataxina-3. Le cellule di Purkinje di topi transgenici
per il gene mutato, mostrano una completa assenza del segnale retrogrado di soppressione sinaptica
endocannabinoide-dipendente. Tale risultato rivela dunque come in modelli murini di SCA-3 il
pathway del recettore metabotropico del glutammato risulti compromesso,come nelle altre forme di
atassia.
Studi recenti hanno inoltre dimostrato come l’Atassia spinocerebellare di tipo 27 (SCA27) possa
derivare da anomalie fisiologiche al livello della sinapsi in questione. E’ stato osservato come
modelli murini con questo tipo di atassia, mostrino una cospicua riduzione nel rilascio di
neurotrasmettitore al livello della sinapsi PC-PF imputata ad un deficit nel funzionamento dei canali
Ca2+-dipendenti nel terminale assonico delle fibre parallele. Questo difetto è dovuto ad una
mutazione del gene FGF14, che codifica per un fattore di crescita essenziale per una corretta
trasmissione sinaptica tra le due cellule. Infatti colture cellulari contenenti cellule dei granuli, fibre
parallele e cellule di Purkinje nelle quali viene completamente eliminato il fattore di crescita,
mostrano una riduzione nella loro trasmissione sinaptica imputabile ad una soppressione del rilascio
del neurotrasmettitore da parte delle fibre parallele.
Anche alcuni disturbi dello spettro autistico sembrano attribuibili ad anomalie fisiologiche del
cervelletto. Pazienti autistici mostrano spesso malformazioni dell’organo o perdita di cellule di
Purkinje nello strato piriforme.
Il ruolo delle PC in questo disturbo è stato esaminato in modelli affetti da sclerosi tuberosa (TSC1),
una facomatosi che si manifesta in associazione a ritardo mentale. Sia gli eterozigoti che gli
omozigoti per la mutazione del gene Tsc, mostrano infatti difetti nell’interazione sociale e
comportamenti ripetitivi che possono essere associati a quelli dei pazienti con autismo. Tuttavia,
nonostante gli eterozigoti non mostrino una diminuzione nel numero di cellule di Purkinje, l’albero
dendritico risulta caratterizzato da un anomalo aumento del numero di spine. Né l’efficacia della
trasmissione sinaptica attribuita alla sinapsi PF-PC né quella CF-PC sembra compromessa, ma le
cellule di Purkinje mostrano una ridotta capacità di generare potenziali d’azione in uscita.
Il 10% dei casi di autismo risulta inoltre caratterizzato dalla mutazione del gene pTen, un
oncosoppressore coinvolto nell’inibizione del pathway di Akt. Topi mutati per il gene e
caratterizzati da visibili comportamenti autistici, mostrano una riduzione nel numero di cellule di
Purkinje dopo il sesto mese di vita, un ingrossamento del soma cellulare, un’anomalia funzionale
nella generazione di potenziali d’azione in uscita ma una eccessiva funzionalità della sinapsi PF-PC.
Questo lavoro di ricerca mostra come la sinapsi Fibra Parallela-Cellula di Purkinje (PF-PC) abbia
un ruolo centrale nel network cerebellare. La plasticità sinaptica al livello della connessione
neuronale entra in gioco durante l’apprendimento motorio e il processo mnemonico. Per questo
motivo, la compromessa funzionalità della sinapsi risulta essere implicata sia in disturbi
neurodegenerativi che dello spettro autistico.
Bibliografia:
1. E. Hoxha et al, Modulation, Plasticity and Pathophysiology of the Parallel Fiber-Purkinje
Cell Synapse; Frontiers in Synaptic Neuroscience 2016.
2. Kimpo RR et al, Gating of neural error signals during motor learning. Elife 2014.
3. Notartomaso et al, Pharmacological enhancement of mGlu1 metabotropic glutamate
receptors causes a prolonged symptomatic benefit in a mouse model of spinocerebellar
ataxiatype1.Mol. Brain 2013.
4. Konno A. et al, Mutant ataxin-3 with an abnormally expanded polyglutamine chain
disrupts dendritic development and metabotropic glutamate receptor signaling in mouse
cerebellar Purkinje cells, Cerebellum. 2014.
Candidati:
Eugenia Giuliani, Christian Maracchioni.
02/02/2017