Conduzione dei potenziali;
sinapsi chimiche
CONDUZIONE DEL POTENZIALE D'AZIONE
Il P.d'A. può insorgere in un punto qualunque
di una cellula o fibra nervosa
All'esterno della membrana, quel punto
diventa elettronegativo rispetto ai punti
contigui; all'interno diventa positivo
Poiché la resistenza elettrica lungo la
membrana non è infinita, si instaurano
correnti elettriche dallo stesso lato della
membrana: le
CORRENTI ELETTROTONICHE
Correnti elettrotoniche
B
A
+ + +
- -mv
-90
- - + + +
- - + + +
+ +
- - +
-60 mv
- - +
+ +
- - + +
B
+ + +
- - -
+ +
- - -
A
B
+ + +
--90
- -mv
- - + + +
+ +
+- -60 mv
+- + +
B
Le CORRENTI ELETTROTONICHE
all'esterno spostano cariche positive
verso il punto eccitato, e all'interno
verso i punti contigui (essendo un
ambiente ionico in cui sono in gioco
cationi, sono questi, che hanno carica
positiva, che si spostano).
Le correnti elettrotoniche non
attraversano la membrana e non
richiedono variazioni di
permeabilità, ma le provocano nei
punti contigui, propagando lo stato
di eccitamento, cioè lo spostamento
del potenziale verso il valore soglia.
Le correnti elettrotoniche sono
graduate (non tutto-o-nulla), si
propagano con decremento, ed hanno
costante di tempo e di spazio. Il
potenziale d'azione viene generato nei
punti contigui quando viene superato il
valore soglia.
VELOCITA' DI PROPAGAZIONE
Le correnti elettrotoniche si propagano con la
velocità della corrente elettrica (300.000
m/s), che è virtualmente infinita per le
distanze da percorrere
il P. d'A. si propaga con velocità finita, perché
è una risposta biologica ad un fenomeno fisico.
La velocità di propagazione dipende dalla
resistenza longitudinale ed è tanto più elevata
quanto più grande è la cellula o più grossa la
fibra.
VELOCITA' DI PROPAGAZIONE NELLE
FIBRE MIELINICHE
la presenza della guaina mielinica in molte
fibre nervose aumenta molto la velocità di
conduzione, perchè questa avviene a salti
(conduzione saltatoria), fra le interruzioni
della guaina (nodi di Ranvier).
La PROPAGAZIONE FRA CELLULE
CONTIGUE è possibile solo quando
esistono punti a bassa resistenza
(giunzioni strette) fra le cellule (sinapsi
elettriche).
Conduzione saltatoria
PRINCIPALI CELLULE ECCITABILI: le
caratteristiche del P. d'A. sono parzialmente
diverse per ogni tipo di cellula e verranno
indicate di volta in volta.
NEURONI (centrali e periferici)
MUSCOLO STRIATO (scheletrico)
MUSCOLO CARDIACO
MUSCOLO LISCIO (visceri e vasi)
Trasmissione sinaptica
Premessa
Il potenziale d’azione si propaga per tutta la
lunghezza dell’assone di qualunque neurone
(nel caso del sistema nervoso periferico,
questo può essere anche assai lungo)
Il potenziale d’azione non si propaga fra
assoni contigui o fra diverse cellule se non in
corrispondenza di strutture specializzate,
genericamente chiamate sinapsi
Trasmissione sinaptica
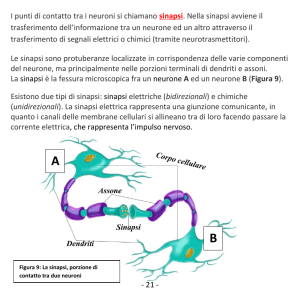
Si chiamano SINAPSI i punti di contatto
fra cellule, in particolare fra le
terminazioni assoniche e le cellule che le
ricevono
Esistono sinapsi elettriche,
prevalentemente nel SNC e nel muscolo
liscio e cardiaco, ma quelle con particolare
struttura e funzione sono le SINAPSI
CHIMICHE.
GIUNZIONE NEUROMUSCOLARE
(PLACCA MOTRICE)
E' l'esempio più tipico e più facile da studiare di
trasmissione sinaptica
Si tratta del punto di congiunzione fra il
terminale assonico di un neurone motore e la
corrispondente fibra muscolare
Il neurone motore è una fibra mielinica che si
divide, in prossimità del muscolo, in numerose
diramazioni, ciascuna delle quali forma una
sinapsi con una sola fibra muscolare
L'insieme di un neurone motore e di
tutte le fibre muscolari da esso
innervate costituisce l'unità
motoria, che è l'elemento unitario
per la regolazione della contrazione
muscolare
STRUTTURA (elementi essenziali):
terminazione nervosa, contenente
vescicole sinaptiche
spazio sinaptico
Dalla parte del muscolo: pliche giunzionali
contenenti i recettori specifici
FUNZIONE
il P. d'A. aumenta la permeabilità al Ca++
della terminazione, permettendone
l'ingresso
il Ca++ provoca la fusione della membrana
delle vescicole con quella della cellula e la
liberazione sincronizzata del mediatore
chimico: l’ACETILCOLINA (ACh).
L'ACETILCOLINA (ACh)
È prodotta dal neurone e portata alla
terminazione con un processo di trasporto
assonico; è sintetizzata anche a livello del
terminale
Si lega ai recettori provocando un aumento
della permeabilità del sodio e del potassio
Ne deriva una depolarizzazione localizzata,
chiamata potenziale di placca (end plate
potential=EPP)
DIFFERENZE FRA P. d'A. E EPP
L'EPP è un fenomeno graduabile: anche in assenza di
P.d'A. presinaptico avviene una continua liberazione
di ACh in piccoli pacchetti di vescicole, ciascuna
delle quali rappresenta un quanto (si parla perciò di
liberazione quantica) di ACh, che provoca un piccolo
EPP (potenziale in miniatura), non sufficiente ad
eccitare la fibra muscolare
La placca motrice non contiene canali per il sodio
voltaggio-dipendenti e perciò non può produrre
P. d'A., mentre l'EPP tende ad un valore
intermedio fra ENa ed Ek (vicino a 0 mV).
Destino dell’acetilcolina
I recettori per l’ACh rispondono nel momento in
cui si accoppiano al mediatore
L’ACh deve essere rimossa per permettere une
nuova trasmissione sinaptica (per aprire una
porta bisogna prima chiuderla)
Lo spazio sinaptico è ricco dell’enzima
acetilcolinesterasi, che idrolizza la molecola di
ACh, separandola in colina ed acetato
Destino dell’acetilcolina
L’ACh liberata dalla terminazione nervosa
diffonde nello spazio sinaptico, dove è attratta
dai recettori
L’ACh diffonde anche nel liquido interstiziale; è
attaccata dall’ACh-esterasi; la sua
concentrazione nello spazio sinaptico diminuisce
e la molecola si stacca dai recettori
La colina è in parte ricuperata dal terminale
assonico dove è utilizzata direttamente per la
costruzione di nuove molecole di ACh
Le SINAPSI CHIMICHE sono costituite da uno
spazio sinaptico, che, all'arrivo dell'impulso
(P.d'A.) viene invaso dalle molecole di un
mediatore chimico (neurotrasmettitore),
liberate dalla terminazione presinaptica
l'altra parte dello spazio sinaptico (membrana
postsinaptica) contiene dei recettori specifici
per il mediatore chimico
il legame del mediatore con il recettore permette
la trasmissione del segnale dalla cellula pre- a
quella post- sinaptica.
Le sinapsi chimiche possono essere eccitatorie o
inibitorie.
Proprietà comuni sono:
il ritardo sinaptico, che rallenta la conduzione;
la monodirezionalità della trasmissione
(soltanto dall'elemento pre- a quello postsinaptico)
Le correnti elettrotoniche, e quindi il P. d'A., si
propagano in tutte le direzioni (per es. lungo
una fibra nervosa), ma la propagazione si
arresta a livello della membrana presinaptica.
Proprietà comuni delle sinapsi sono:
la presenza, nel terminale, di vescicole che
contengono il mediatore chimico (sempre uguale
per lo stesso neurone): questo viene liberato
all’arrivo del potenziale d’azione
la presenza di sistemi (enzimatici) per la rimozione
del mediatore chimico
a livello delle sinapsi non c'è continuità anatomica
fra le due cellule.
SINAPSI INTERNEURONICHE
In generale i neuroni emettono una singola fibra
efferente, detta assone, ma hanno numerose
propaggini, dette dendriti, sulle quali si trovano le
sinapsi afferenti (quelle che il neurone riceve)
I dendriti contraggono sinapsi con un numero anche
molto elevato di terminazioni di altri neuroni (bottoni
sinaptici)
Le sinapsi interneuroniche possono essere eccitatorie,
ma anche inibitorie
L'attivazione di una singola sinapsi non è normalmete
in grado di eccitare il neurone, ma è necessaria la
sincronizzazione di molti elementi presinaptici.
Le sinapsi eccitatorie provocano sulla membrana
postsinaptica una piccola depolarizzazione (EPSP)
meccanismo generale: il legame mediatorerecettore apre canali ionici indifferenti, che
provocano una corrente K+ verso l’esterno e una
Na+ verso l’interno. La membrana si depolarizza
fino ad un valore ≈ (ENa+EK)/2 ≈ 0 mV
Le sinapsi inibitorie provocano sulla membrana
postsinaptica una piccola iperpolarizzazione
(IPSP). Meccanismo generale: il legame
mediatore-recettore apre canali ionici per K+ e Cl-;
l’uscita di K+ e l’ingresso di Cl- iperpolarizzano la
mambrana
A livello delle sinapsi non si generano potenziali
d’azione (ma solo EPSP o IPSP) perché non sono
presenti canali Na+ voltaggio-dipendenti
Le attività sinaptiche danno luogo a correnti
elettrotoniche, che convergono verso un punto
preciso del neurone, il monticolo assonico, dal quale
origina l'assone.
Sul monticolo assonico sono presenti canali Na+
voltaggio-dipendenti, che danno origine a potenziali
d’azione ogni volta che viene superata la soglia. In
questo punto la soglia è più bassa che nel resto del
corpo cellulare
Avviene continuamente una sommazione
(spaziale e/o temporale) fra le diverse
attività eccitatorie ed inibitorie, che fa
fluttuare il potenziale a livello del
monticolo assonico: qui si originerà un
potenziale d'azione, che percorrerà
tutto l'assone fino alle sue
terminazioni, ogni volta che viene
superato il livello soglia.
Soglia
EPSP
IPSP
Sommazione temporale
+
=
Sommazione spaziale
Potenziale di riposo
I
I
E
E
I
E
E
Sommazione spaziale
I
E
E
Sommazione temporale
PROPRIETA' INTEGRATIVE DELLE SINAPSI
CENTRALI
l'attività di ogni neurone è “modulata in
frequenza", mentre il potenziale del corpo
cellulare (in particolare del monticolo assonico)
fluttua in continuazione ed è pertanto "modulato
in ampiezza" . Modulato = codificato
Se prevalgono gli IPSP, la probabilità di scarica
si riduce, fino ad annullarsi; se prevalgono gli
EPSP la cellula scaricherà con una frequenza
tanto più grande quanto maggiore è la corrente
depolarizzante che investe il monticolo assonico.
Corrente depolarizzante piccola: bassa frequenza di scarica
I
E
E
I
E
I
E
Corrente depolarizzante intensa: alta frequenza di scarica
I
E
E
E
E
E
E
INIBIZIONE PRESINAPTICA:
sinapsi inibitorie si possono trovare anche in
prossimità delle terminazioni assonali: quando sono
attivate bloccano la trasmissione del P. d'A. e
impediscono la liberazione del mediatore.
MEDIATORI CHIMICI CENTRALI:
sono numerosi e non tutti identificati; la loro
azione non è costante, ma dipende dal particolare
tipo di relazione mediatore-recettore. Tuttavia,
per lo stesso neurone, l’azione è sempre uguale
MEDIATORI CENTRALI:
Ach, noradrenalina, adrenalina, dopamina (catecolamine),
serotonina
INIBITORE: ac. Gamma-aminobutirrico (GABA)
ECCITATORE: glutammato
NEUROMODULATORI