INTRODUZIONE AL METABOLISMO
Uno degli aspetti più importanti del biochimico è
capire come la cellula sia in grado di regolare la miriade
di sequenze di reazioni che avvengono in essa
VIA METABOLICA = sequenza di reazioni
CATABOLISMO = degradazione sostanze complesse
ANABOLISMO
complesse
=
sintesi
di
sostanze
organiche
STADIO
1
STADIO
2
STADIO
3
POLIMERI:
Proteine,
degli acidi nucleici,
Polisaccaridi, lipidi
Monomeri:
Amminoacidi, nucleotidi,
zuccheri, acidi grassi, glicerolo
Intermedi metabolici:
Piruvato, acetil-CoA,
intermedi del ciclo
dell’acido citrico
CONSUMO
PRODUZIONE
NETTA DI
ENERGIA
PICCOLE MOLECOLE
SEMPLICI
NETTO DI
ENERGIA
Ogni via metabolica è caratterizzata da tre stadi di
complessità:
STADIO 1 = interconversione dei polimeri e dei lipidi
complessi negli intermedi monomerici;
STADIO 2 = interconversione di zuccheri monomerici,
amminoacidi e lipidi in composti organici ancora più
semplici;
STADIO 3 = degradazione finale a composti organici
semplici tra cui CO2 H2O e NH3, oppure la sintesi a
partire da tali composti
METABOLISMO INTERMEDIO = reazioni che riguardano
immagazzinamento e produzione di energia metabolica
e impiego di questa energia per le sintesi di altri
composti a basso PM e per sostanze di riserva. Sono
reazioni che non richiedono una molecola di acido
nucleico come stampo
METABOLISMO ENERGETICO = parte del metabolismo
intermedio che consiste delle vie che immagazzinano e
generano energia metabolica
VIE METABOLICHE CENTRALI = vie sostanzialmente
identiche nei vari organismi e responsabili della
manipolazione di grandi quantità di energia e materia.
Rappresentano le vie metaboliche principali
VIE CENTRALI NEL METABOLISMO
Una delle vie più importanti è la GLICOLISI (stadio 2)
che catabolizza i carboidrati sia nelle cellule aerobie
che anaerobie.
Il substrato è rappresentato dal GLUCOSIO, che deriva
dai polisaccaridi di riserva o è introdotto tramite la
dieta.
La via porta al PIRUVATO + CHETOACIDI a tre atomi di
carbonio
Negli ANAEROBI il piruvato viene ridotto ad altri
composti tra cui ETANOLO e CO2 (FERMENTAZIONE).
Nel metabolismo ossidativo (AEROBI) il principale
destino del piruvato è la sua ossidazione ad un
composto metabolicamente attivato, l’ACETIL-CoA.
Gli atomi di carbonio di questo composto possono
subire
una
ulteriore
ossidazione
nel
CICLO
DELL’ACIDO CITRICO (stadio 3). Tramite questa via si
arriva all’ossidazione completa del glucosio sino a CO2
ed acqua.
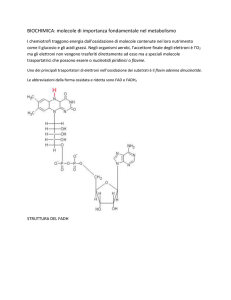
Le reazioni ossidative di questo ciclo producono
trasportatori ridotti di elettroni (NADH e FADH2), la cui
riossidazione promuove la sintesi di ATP
Anche altre vie ossidative portano “carburante” al ciclo
dell’acido citrico. L’acetil-CoA deriva infatti anche dalla
ossidazione degli acidi grassi mediante un processo
noto come - OSSIDAZIONE e da alcune vie ossidative
degli amminoacidi.
Se l’acetil-CoA non viene completamente ossidato può
rappresentare il substrato di partenza di vie
metaboliche di sintesi, per esempio degli acidi grassi.
Questa ed altre vie biosintetiche riduttive utilizzano
come trasportatore di elettroni il coenzima ridotto,
NADPH.
Durante il corso ci occuperemo anche di alcuni
processi coinvolti nella biosintesi dei carboidrati, come
la GLUCONEOGENESI, importante negli organismi
eteretrofi e la FOTOSINTESI, processo chiave
attraverso il quale le piante verdi “catturano” l’energia
luminosa per la produzione di potere riducente
(NADPH) ed energia (ATP) necessari per la sintesi dei
carboidrati.
COESISTENZA DI VIE DEGRADATIVE E
BIOSINTETICHE
Abbiamo detto che l’ossidazione dei grassi porta alla
liberazione di molecole di acetil-CoA
Gli acidi grassi vengono sintetizzati a partire da
molecole di acetil-CoA
Queste due vie metaboliche sono ben distinte l’una
dall’altra anche se possono presentare intermedi o
alcune reazioni enzimatiche comuni
Queste due vie metaboliche
compartimenti cellulari diversi
avvengono
in
due
Occorre inoltre sottolineare che le biosintesi non sono
mai l’esatto contrario delle vie degradative anche se
spesso iniziano e terminano con gli stessi metaboliti
LE VIE DEGRADATIVE E QUELLE
BIOSINTETICHE SONO INOLTRE DISTINTE
PER DUE MOTIVI:
- UNA VIA PUO’ ESSERE ESOERGONICA
IN UNA SOLA DIREZIONE
- LE VIE DEBBONO ESSERE REGOLATE
SEPARATAMENTE PER EVITARE CICLI
FUTILI
CONSIDERAZIONI DI BIOENERGETICA
Ogni reazione esoergonica in grado di liberare
sufficiente energia può essere accoppiata ad una
endoergonica
Negli organismi viventi la maggior parte dell’energia
libera richiesta per promuovere le reazioni biochimiche
deriva dall’ossidazione dei substrati organici
L’ossigeno, un potente ossidante, rappresenta
l’accettore finale di elettroni negli organismi AEROBICI
Per questa tendenza dell’ossigeno e per la sua elevata
presenza nell’atmosfera, gli organismi viventi hanno
evoluto la capacità di ricavare energia dall’ossidazione
dei composti organici
Le ossidazioni biologiche
L’ossidazione biologica dei composti organici è
sostanzialmente uguale alle ossidazioni non biologiche
come ad esempio la combustione del legno
Le ossidazioni biologiche sono però molto più
complesse; quando il legno brucia tutta l’energia viene
rilasciata sotto forma di calore
Le ossidazioni biologiche non determinano un elevato
aumento della temperatura ed una parte dell’energia
viene convertita in energia chimica (soprattutto ATP)
La maggior parte delle ossidazioni biologiche non
comporta un trasferimento diretto di elettroni da un
substrato ridotto all’ossigeno
Più frequentemente si verifica una serie di reazioni
accoppiate di ossidoriduzione nelle quali gli elettroni
vengono trasferiti ad una serie di trasportatori
intermedi
(CATENA
DI
TRASPORTO
DEGLI
ELETTRONI) come il NAD+ e da ultimo sono trasferiti
all’ossigeno (ACCETTORE FINALE DEGLI ELETTRONI)
L’energia delle ossidazione viene rilasciata in piccole
aliquote in modo da controllare meglio l’ossidazione e
catturare l’energia appena viene rilasciata.
Non tutta l’energia comunque deriva dall’ossidazione
dell’ossigeno
Molti microrganismi possono o devono vivere in
condizioni anaerobiche ed in questo caso essi
espletano il processo delle fermentazioni, vie
cataboliche in grado di produrre energia e che
procedono in assenza di ossigeno
Se l’energia deriva essenzialmente dalle reazioni di
ossidazione ne deriva che quanto più uno strato è
ridotto quanto maggiore sarà la sua capacità di
generare energia
Tra le molecole organiche, i grassi sono i composti che
liberano la maggior parte di energia
C6H12O6 + 6O2 6CO2 + 6H2O
G0’ = - 15,64 kJ/g
C16H32O2 + 23O2 16CO2 + 16H2O
G0’ = - 38,90 kJ/g
- i grassi hanno un maggior numero di protoni ed
elettroni in grado di legarsi all’ossigeno rispetto al
glucosio
L’ATP come moneta di scambio energetico
Le cellule viventi catturano l’energia libera rilasciata dal
catabolismo principalmente sotto forma di ATP
L’energia chimica immagazzinata nell’ATP può essere
convertita in altre forme di energia nel processo noto
come TRASDUZIONE DELL’ENERGIA
L’ATP non sarebbe l’unica molecola con legami ad alta
energia;
tuttavia,
molti
enzimi
si
legano
preferenzialmente all’ATP ed utilizzano la sua energia di
idrolisi per rendere possibili reazioni endoergoniche
PRINCIPALI MECCANISMI DI CONTROLLO DEL
METABOLISMO
Tra i principali meccanismi abbiamo certamente il
controllo delle attività enzimatiche, come la
concentrazione del substrato e la regolazione
allosterica
Controllo livelli enzimatici: induzione e repressione
sintesi enzima
Controllo attività enzimatica: interazioni con substrati,
prodotti ed effettori allosterici e modificazione
covalente della proteina
Compartimentazione:
organelli cellulari
Regolazione ormonale
nell’ambito
di
determinati