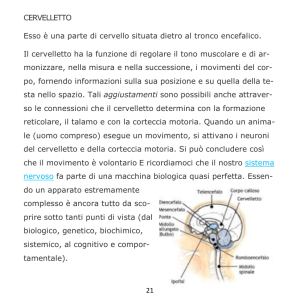
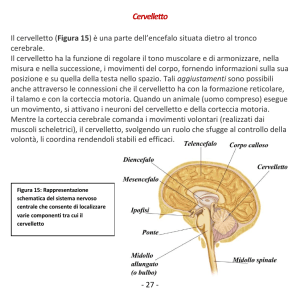
Il cervelletto
È l’organo integrativo per:
– Raffinamento e coordinazione dei movimenti
– Regolazione del tono muscolare e mantenimento
dell’equilibrio
– Apprendimento dei movimenti
Corteccia cerebellare e nuclei profondi
Il cervelletto è’ posto sulla faccia posteriore del tronco cerebrale al quale è
collegato da tre paia di peduncoli.
Possiede una corteccia grigia superficiale che ricopre la sostanza bianca dentro
cui sono dispersi nuclei di sostanza grigia.
Sia anatomicamente che funzionalmente si distinguono porzioni più antiche che
esistono in tutti i vertebrati (archicerebello e paleocerebello) ed altre che si
sono invece sviluppate solo nei vertebrati superiori (neocerebello)
Anatomia del cervelletto
È diviso orizzontalmente in tre lobi.
1. Lobo anteriore.
2. Lobo posteriore.
2. Lobo flocculonodulare.
Anatomia del cervelletto
È diviso verticalmente in tre regioni.
1. Verme.
2. C. intermedio.
2. C. laterale.
La superficie del verme e dei lobi è percorsa da solchi trasversali concentrici che
delimitano la massa cerebellare in lamelle. Le lamelle del cervelletto sono
disposte orizzontalmente e sono più piccole di quelle della corteccia cerebrale.
Corteccia cerebellare
Lamelle
Solco
Strato molecolare
Strato delle cellula del Purkinje
Strato granulare
Sostanza bianca
Nuclei profondi del cervelletto
n. globoso e
emboliforme
n. del tetto
(fastigio)
n. dentato
NUCLEO DEL FASTIGIO (TETTO) - riceve dal
verme, dai nuclei vestibolari e dall’oliva inferiore.
Proietta ai nuclei vestibolari e ad altri nuclei bulbari
NUCLEO GLOBOSO - riceve dal verme. Proietta a
nuclei bulbari
NUCLEO EMBOLIFORME - riceve dalla parte
intermedia degli emisferi. Proietta al talamo
NUCLEO DENTATO - riceve dal neocerebello
(emisferi cerebellari). Proietta al nucleo rosso ed al
talamo
Gran parte delle fibre afferenti ed efferenti hanno come
stazione di relè i nuclei profondi del cervelletto
La corteccia cerebellare è organizzata
in tre strati
fibre parallele
SM
cellula
stellata
SP
cellula a
canestro
strato
molecolare
SG
strato cell.
Purkinje
cellula
del Golgi
cellula del
Purkinje
strato
granulare
fibra
rampicante
cellula
granulare
fibra
muscoide
CORTECCIA CEREBELLARE
STRATO MOLECOLARE
• CELLULE STELLATE
STRATO
DIAPURKINJE
• CELLULE
CANESTRO
•• SOMA
STRATO
DEI GRANULI
CELLULE DI PURKINJE (unica via efferente dalla
FIBRE DELLE
PARALLELE
•• CELLULE
corteccia cerebellare,
DEI
GRANULI
portano impulsi
inibitori)
DENDRITI
DELLE
CELLULE
DI PURKINJE
•• ASSONI
FIBRE
RAMPICANTI
E SOMA
CELLULE
DEL GOLGI
DENDRITI
DELLEDELLE
CELLULE
DEL GOLGI
DI II° TIPO
• FIBRE MUSCOIDI
• ASSONI DELLE CELLULE DI PURKINJE
Fibre rampicanti
• Originano dai neuroni olivo-cerebellari di cui
rappresentano le fibre terminali
• Salgono attraverso lo strato granulare
• Finiscono nello strato molecolare con un ciuffo di
arborizzazioni terminali
• Si connettono ai dendriti delle cellule del Purkinje
(rapporto 1:1) ma ciascuna può formare numerosi
contatti sinaptici con l’albero dendritico della c. del P.
• Liberano il neurotrasmettitore glutamato
dall’oliva
bulbare
Fibre muscoidi
• Input: da neuroni del ponte e del midollo spinale
• Costituiscono tutte le rimanenti afferenze cerebellari
• Si ramificano e terminano nello strato granulare
• Hanno terminazioni dilatate (rosette)
• Formano sinapsi con:
– i dendriti delle cellule dei granuli
– gli assoni dei neuroni del Golgi
da ponte
e midollo
spinale
Cellule dei granuli
• sono neuroni molto piccoli (7mm), sterttamente
impachettati
• Piccolo albero dendritico nello strato granulare
• Un assone amielinico
– che si dirige verso lo strato molecolare (centrifugo)
– e si divide a T per formare la fibra parallela
– Le fibre parallele decorrono longitudinalmente
lungo le lamelle
– Incrociano i dendriti di molte cellule del Purkinje
• Liberano glutamato come neurotransmettitore
Cellule di Golgi
sono localizzate nello strato granulare; il
loro assone, insieme alle fibre muscoidi e
ai dendriti delle cellule dei granuli
costituiscono una struttura ciamata
glomerulo.
Glomerulo cerebellare
E’un complesso di sinapsi tra la terminazione di una fibra muscoide al centro, e:
a) dendriti di cellule granulari, b) assoni di neuroni del Golgi
Assone di una
cellula del Golgi
Dendrite di un
granulo
Fibra muscoide
Terminazione di
una fibra muscoide
Assone di una
cellula del Golgi
Dendrite di un
granulo
Fibra muscoide
Nel glomerulo cerebellare il terminale assonico di una fibra muscoide (in blu) invia
messaggi di tipo sensoriale a ~50 cellule granulari (in rosso) (divergenza).
Cellule di Purkinje
Consistono di:
• Un ampio albero dendritico nello strato
molecolare, con spine dendritiche a livello
delle sinapsi
• Un grosso soma
• Un assone che costituisce la via efferente
dal cervelletto, e manda collaterali nello
strato granulare.
• GABA è il neurotrasmettitore liberato
• ogni cellula di Purkinje riceve una fibra rampicante (rapporto 1:1), che può
formare però numerosi contatti sinaptici con il suo albero dendritico
• ogni cellula di Purkinje riceve inoltre circa 200,000 contatti sinaptici dalle fibre
parallele (convergenza)
• ogni fibra parallela prende contatto con numerose cellule di Purkinje (divergenza)
• gli assoni delle cellule di Purkinje costituiscono la principale via efferente del
cervelletto, attraverso i nuclei cerebellari. Ciascun assone forma circa 1000
contatti con le cellule dei nuclei profondi. Il neurotrasmettitore liberato è il
GABA.
Le cellule stellate e a canestro sono situate trasversalmente nello strato molecolare
Cellule stellate
– Nella parte superficiale dello strato molecolare
– Formano sinapsi con i dendriti delle cellule del
Purkinje
– Azione inibitoria
Cellule a canestro
– Nella parte profonda dello strato molecolare
– Gli assoni formano un canestro attorno
all’encoder delle cellule del Purkinje
– Azione inibitoria
Circuiti cerebellari di base
Il circuito di base è lo stesso in tutte le parti del cervelletto. Esso è costituito da tre parti
1. Via diretta : l’input afferente proietta
direttamente ai sistemi motori attraverso i
nuclei profondi senza passare per la
corteccia.
cellula
2. Circuito laterale indiretto: input delle granulare
fibre muscoidi alle cellule granulari, da
queste mediante le fibre parallele alle
cellule del Purkinje, e da queste ritorno ai
fibra
fibra
rampicante
cellula
del
nuclei profondi del cervelletto. Questo
muscoide
Purkinje
circuito è usato per correggere le risposte
riflesse.
Nucleo cerebellare
3. Le fibre rampicanti mandano un input
profondo
alle cellule del Purkinje. Questo è l’input
di rilevamento dell’errore. Questo circuito
Input
Output
è usato per l’apprendimento.
(dagli organi di senso)
(ai sistemi motori)
Circuiti cerebellari di base
Le fibre parallele decorrono parallelamente ai solchi.
Esse intersecano l’arborizzazione delle cellule del Purkinje ad angolo retto.
Gli input delle fibre parallele e rampicanti hanno effetti molto diversi
sulle cellule del Purkinje
Molte fibre parallele (~200,000) prendono contatto
con una singola cellula del Purkinje (convergenza).
Ciascuna sinapsi genera un debole PPSE e sono
richiesti molti inputs (sommazione) per generare un
potenziale d’azione. La scarica generata è duratura
e ritardata rispetto a quella generata dalle fibre
rampicanti (spikes semplici: azione tonica lenta)
Un’unica fibra rampicante contatta ciascuna
cellula del Purkinje formando molte sinapsi.
Ciascun singolo input genera una rapida
scarica di potenziali d’azione di latenza e
durata brevi (spikes complessi: azione
fasica rapida).
Si ritiene che l’attivazione delle fibre rampicanti possa rendere silente per alcune
centinaia di ms la scarica evocata nelle cellule del Purkinje dalle fibre parallele
Spikes semplici e complessi generati dalle cellule del Purkinje
Le cellule di Purkinje normalmente generano potenziali d’azione ad elevata
frequenza anche in assenza di inputs sinaptici. La frequenza media durante la veglia
è di circa 40 Hz (40 spikes per sec).
I treni mostrano una mistura di cosiddetti spikes semplici e complessi.
Spikes semplici
S. complessi
Gli spikes complessi sono spesso
seguiti da una pausa di alcune
centinaia di msec durante la
quale l’attività degli spikes
semplici è soppressa.
50 ms
Uno spike semplice è un singolo
potenziale d’azione seguito da un
periodo di refrattarietà di circa 10
msec.
Gli spikes semplici sono prodotti da
una combinazione di attività basale
e di inputs dalle fibre parallele.
Uno spike complesso è una sequenza
stereotipata di potenziali d’azione con
intervalli molto brevi tra uno spike e l’altro
e ampiezze decrescenti.
Gli spikes complessi, (che si susseguono ad
una velocità basale di circa 1 Hz, massimo
10 Hz) sono costantemente associati
all’attivazione delle fibre rampicanti.
L’azione delle cellule del Purkinje sui nuclei cerebellari
profondi è inibitoria
L’unico output dal cervelletto è rappresentato
dalle cellule del Purkinje.
I neuriti delle cellule del Purkinje inibiscono i
nuclei profondi liberando GABA.
I segnali inibitori da essi inviati ai neuroni dei
nuclei profondi ne frenano continuamente
l’attività.
Il ritardo, l’intensità e la durata dell’inibizione
prodotta sono il risultato della complessa
elaborazione attuata nella corteccia cerebellare.
GABA
Quindi, ai neuroni dei nuclei profondi, che sono la stazione d’uscita dei segnali dal
cervelletto, giungono immediatamente e direttamente i segnali eccitatori portati
dalle collaterali di tutte le fibre afferenti al cervelletto e, successivamente, i segnali
inibitori provenienti dalle cellule del Purkinje.
Circuiti cerebellari di base
Spikes semplici
S. complessi
Formano sinapsi inibitorie con le
cellule del Purkinje.
fibre
parallele
Afferenze eccitatorie: fibre parallele.
cellula a
canestro
cellula
stellata
cellula del
Purkinje
cellula
del Golgi
cellula
granulare
Funzione: accentuano il contrasto tra i
gruppi di cellule del Purkinje attivi.
fibra
rampicante
Cellule del Golgi (strato granulare).
Afferenze eccitatorie: fibre parallele.
Contribuiscono alla costituzione del
glomerulo cerebellare, formando
sinapsi inibitorie con i dendriti delle
cellule dei granuli e sopprimendone
l’eccitamento in risposta
all’attivazione delle fibre muscoidi.
Funzione: scansione temporale dei
processi eccitatori.
fibra
muscoide
neuroni
profondi del
cervelletto
fibre
ascendenti
dagli organi
di senso
oliva inferiore
fibre disc.
ai sistemi
motori
Circuito eccitatorio principale
Cellule stellate e a canestro (strato
molecolare).
Circuito inibitorio collaterale
Circuiti inibitori collaterali
La circuiteria del cervelletto
(+)
G
(+)
(+)
(+)
St
(-)
(+)
P
(-)
(-)
GABA (-)
Gr
GABA (-)
GABA (-)
fibra muscoide
(+)
da bulbo e midollo spinale
(+)
C
P
fibra rampicante
fibra parallela
P ≡ c. del Purkinje
C ≡ c. a canestro
G ≡ c. del Golgi
St ≡ c. stellata
Gr ≡ c. granulare
dall’oliva inferiore
nuclei
profondi
dendriti
eccitatorie
inibitorie
Le cellule stellate e a canestro causano un’inibizione laterale delle
cellule del Purkinje situate ai lati del fascio di fibre parallele attivate
cellula del Purkinje
Le cellula del P. che non ricevono
afferenze dalle fibre parallele del
fascio vengono inibite dalle cellule
stellate e a canestro
Fascio di fibre parallele eccitate.
Le cellule del P. che ricevono
afferenze dalle fibre parallele del
fascio vengono eccitate
cellula stellate e a
canestro
Anche le cellule del Golgi ricevono un input dalle fibre parallele, ma distribuiscono i
loro assoni indietro alle cellule granulari. È un esempio di inibizione a feedback che
taglia gli input dopo un breve ritardo, tendendo ad accorciare la durata dei treni di
potenziali d’azione nelle fibre parallele.
ORGANIZZAZIONE MORFOFUNZIONALE
DEL CERVELLETTO
Paleocerebellum o
spinocerebello
Neocerebellum o
cerebrocerebello
Verme
Archicerebellum o
vestibolocerebello
(lobo flocculo-nodulare)
Le principali funzioni di ciascuna suddivisione
2. 3.
Paleocerebellum
Neocerebellumoo
1. cerebrocerebello
Archicerebellum o
Spinocerebello
vestibolocerebello
Input:
daldalla
midollo
spinale (vie
Input:
corteccia
Input:
dagli organisensibilità
vestibolari.
spino-cerebellari),
cerebrale.
propriocettiva
ecorteccia
cutanea.
Output:
ai alla
muscoli
di gambe,
Output:
tronco
e occhi.
Output:
al midollo
cerebrale
motoria spinale.
e premotoria.
controllo
dell’equilibrio
Funzione:
regolazione
del tono
Funzione:
pianificazione
e
e dei
movimenti
oculari
muscolare,
della
postura
e
dei
avvio dei movimenti volontari.
movimenti involontari.
Funzione generale del cervelletto:
Agisce sul controllo motorio
Quindi, dal cervelletto non dipende tanto la capacità di compiere i
movimenti, quanto la capacità di compierli in modo corretto.
Ad es., sollevare una scatola. Il sollevamento iniziale viene regolato in
base al peso supposto della scatola.
Quando avviene l’intervento regolatore?
Prima, durante e alla fine di ogni movimento in modo da regolarne:
• durata,
• ampiezza
• gradualità.
Consideriamo ad esempio il controllo eseguito dal cervelletto sui movimenti volontari
Inputs al cervelletto:
- dall’apparato propriocettivo muscolare (fascio spino cerebellare) una completa
informazione sulla posizione che le diverse parti del corpo assumono istante per istante;
- dalla corteccia cerebrale (fascio cortico-ponto cerebellare) una completa informazione
sugli “ordini” motori elaborati dalla corteccia cerebrale.
Azioni del cervelletto:
- confronto tra quadro delle afferenze propriocettive e delle efferenze motorie;
- rilevazione degli “errori” negli ordini;
- correzione in tempo reale con segnali inviati alla corteccia tramite la via ascendente
cerebello-talamo-corticale e al midollo spinale tramite la vie discendenti (cerebellorubro-, cerebello-reticolo- e cerebello-vestibolo-spinali).
Inizio del movimento
Corteccia premotoria
e motoria
Neocerebellum
Esecuzione motoria
e correzione
movimento
Spinocerebellum
feedback
sensoriale
Il cervelletto nell’apprendimento motorio
Il cervelletto potrebbe avere un ruolo nell’apprendimento di nuove capacità motorie.
I segnali convogliati dalle fibre rampicanti modificherebbero, in periodi di tempo
lunghi, le risposte dei neuroni di Purkinje rispetto ai segnali che arrivano attraverso
le fibre muscoidi.
Dati sperimentali dimostrano che i circuiti cerebellari vengono modificati con
l’esperienza e che queste modifiche sono importanti nell’apprendimento motorio.
Il cervelletto nell’apprendimento motorio
Esempio 1. Aggiustamento del riflesso vestibulo-oculare
Cos’è il riflesso vestibulo-oculare
(VOR)?
• La funzione del VOR è di stabilizzare
l’immagine della retina durante le
rotazioni della testa.
• quando la testa ruota ad una certa
velocità e in una certa direzione, l’occhio
ruota alla stessa velocità ma in direzione
opposta.
• Senza il VOR, l’occhio vedrebbe
un’immagine offuscata ogni volta che si
muove la testa.
Ciò accade perché l’occhio è come una
macchina fotografica con velocità di
otturazione lenta.
Il cervelletto nell’apprendimento motorio
Esempio 1. Aggiustamento del riflesso vestibulo-oculare
Quindi, lo scopo del VOR è di ruotare l’occhio in direzione opposta alla testa,
mantenendo ferma l’immagine sulla retina. Il VOR è realizzato tramite due vie.
2) Una via indiretta
attraverso le fibre muscoidi,
le fibre parallele e le cellule
del Purkinje del cervelletto,
che inibisce il nucleo
vestibolare e permette le
opportune correzioni.
cellula
granulare
fibra
muscoide
cellula del
Purkinje
canale
semicircolare
occhio
n. vestibolare
1) Una via diretta dai canali semicircolari (e nucleo vestibolare) agli appropriati
muscoli dell’occhio causandone la contrazione (vedi sist. vestibolare)
Supponiamo che a causa di un trauma o dell’età alcuni neuroni coinvolti nel
riflesso funzionino male o muoiano e di conseguenza i muscoli dell’occhio non si
contraggano più adeguatamente. Occorre qualcosa che ponga riparo a ciò.
3) Quando il VOR non lavora adeguatamente
(p.es., l’occhio non ruota abbastanza) uno
scorrimento dell’immagine è captato dall’occhio e
inviato al cervelletto tramite le fibre rampicanti.
L’imput dalle fibre rampicanti altera in maniera
semi-permanente le sinapsi delle fibre parallele
attivate simultaneamente.
fibra
rampicante Quando il VOR è riparato (cioè nessuno
scorrimento dell’immagine sulla retina) l’attività
delle fibre rampicanti cessa.
oliva
Quindi le fibre rampicanti istruiscono l’azione
inferiore
delle fibre muscoidi (apprendimento motorio o
adattamento).
Il cervelletto nell’apprendimento motorio
Esempio 2.
Un prisma davanti agli occhi altera la posizione apparente degli oggetti
1) Lanciando il dardo con
un prisma
2) Senza
Gradualmente
davanti
il
prismaagli
occhi,l’errore
ilcervelletto
bersaglio
appare
aiuta a
è piccolo
spostato
correggere
di lato el’errore.
quindi
viene mancato.
errore nel
lancio del
dardo
3) Togliendo i prismi, il bersaglio viene di nuovo sbagliato, ma nella direzione
opposta.
4) Il cervelletto, di nuovo, correggerà l’errore.
Gli occhiali hanno un effetto simile a quello dei prismi. Il VOR è ricalibrato
ogni volta che vengono utilizzate nuove lenti.
Il cervelletto nell’apprendimento motorio
Esempio 3. apprendimento di nuovi riflessi posturali
Normalmente, quando si sta
in piedi su una superficie
ferma (pavimento), una
distensione dell’estensore
significa che ci si sta
sporgendo in avanti troppo e
e la risposta corretta
dell’azione riflessa è di
contrarre l’estensore.
EMG del muscolo
estensore
Il cervelletto nell’apprendimento motorio
Esempio 3. apprendimento di nuovi riflessi posturali
Quando ci si trova su una
piattaforma in equilibrio su
una palla, lo stesso riflesso
provoca una caduta.
EMG del muscolo
estensore
Il cervelletto nell’apprendimento motorio
Esempio 3. apprendimento di nuovi riflessi posturali
Bisogna imparare a rilassare
l’estensore, quando la
piattaforma stira l’estensore.
Gradualmente il cervelletto
impara a sopprimere questo
riflesso di contrazione.
Non c’è apprendimento se il
cervelletto è danneggiato.
EMG del muscolo
estensore
Lesioni del neocerebellum provocano atassia
L’atassia consiste in:
1) Un ritardo nell’inizio del movimento.
Vi è anche una maggiore lentezza nel movimento perché l’attività del muscolo
agonista è meno fasica.
2) Dismetria: il movimento supera o non raggiunge il bersaglio.
3) Tremore: Si verifica dopo un movimento o quando c’ò una perturbazione nel
momento in cui si sta tentando di tenere un arto in una posizione particolare.
Il tremore non si manifesta a riposo.
In conclusione
Il cervelletto opera come un’officina per riparazioni.
1) Impara ad adattarsi a nuovi compiti
2) I disordini motori prodotti da lesioni al di fuori del cervelletto
sono velocemente mascherati da quest’officina.
3) Le lesioni cerebellari provocano disordini motori perché se
l’officina è danneggiata i disordini diventano evidenti.
Il deficit persiste se esso eccede le capacità riparatorie
dell’officina – p. es., non c’è compensazione per un muscolo
totalmente paralizzato.
FINE
Vie afferenti
dalle fibre rampicanti
Aree
motoria e
premotoria
1.
movimento
muscolare
Area
sensibilità
somatica
2.
per avere le
sensazioni
tattili
Area visiva
associativa
3.
per vedere
cosa stiamo
facendo
Cervelletto
Oliva
La via cortico-olivo-cerebellare
convoglia inputs dalle aree associative
sensoriale, motoria e visiva al
cervelletto attraverso le fibre
rampicanti.
Vie afferenti
dalle fibre muscoidi
Le fibre muscoidi convogliano informazione richiesta per fare degli aggiustamenti
• Spinocerebellari
– Informazione di feedback da muscoli e articolazioni
• Vestibulo-cerebellari
– Feedback circa la postura e il movimento
• Cortico-ponto-cerebellari
– Dalle aree associative
– Aggiustamenti motori che richiedono giudizio, discriminazione, e decisioni
Vie efferenti
L’output dal cervelletto avviene tramite le cellule del Purkinje e i nuclei profondi
Gli asssoni delle cellule del Purkinje terminano
sui neuroni che formano i nuclei profondi: essi
sono tutti inibitori
Output dai nuclei
profondi a vari centri
n. fastigio
n. vestibolari
formazione reticolare
n. emboliforme
n. globoso
n. rosso
talamo
n. dentato
corteccia cerebrale