Livelli energetici ed orbitali
Gli elettroni orbitano intorno al nucleo secondo schemi particolari. Gli elettroni possono
sistemarsi solo a certe distanze dal nucleo, formando così strati o gusci elettronici, a
ciascuno dei quali compete una particolare energia caratteristica (livello energetico) e
quantizzata in quanto può possedere solo certi valori.
Un elettrone può passare da un livello più interno di energia E i ad uno più esterno libero di
energia Ee, solo se gli viene fornita esattamente la quantità di energia Ee-Ei. Ciò significa che
ogni atomo può assorbire solo certe quantità di energia caratteristiche. Quando un atomo
assorbe energia un elettrone passa da un livello più interno ad uno più esterno (salto
quantico) e l’atomo si viene a trovare in una situazione di instabilità (stato eccitato).
Quando l’elettrone ritorna al livello energetico di partenza viene restituita esattamente la
medesima quantità di energia assorbita e l’atomo riacquista stabilità (stato fondamentale).
Il legame chimico
La vita della cellula dipende dalla sua capacità di immagazzinare ed utilizzare energia.
L'unico modo che la cellula possiede per immagazzinare energia è di fissarla nei
legami chimici che tengono uniti gli atomi nelle molecole. Attraverso una continua
rottura e ricostruzione dei legami e demolizione - riassemblaggio di molecole (metabolismo
cellulare) la cellula fa circolare l'energia al suo interno. Una caratteristica importante di ogni
legame chimico è il suo contenuto energetico (energia di legame).Durante una reazione
chimica vengono rotti alcuni legami dei reagenti mentre si formano nuovi legami necessari per
la sintesi dei prodotti.
Una reazione libera energia (reazione esoergonica) se i legami che si spezzano sono più
energetici dei legami che si formano. I reagenti sono più energetici dei prodotti.
Energia
A + B (reagenti)
E (energia liberata)
C + D (prodotti)
tempo
Se al contrario una reazione avviene con assorbimento di energia (reazione endoergonica)
ed i legami che si spezzano sono meno energetici dei legami che si formano, i reagenti sono
meno energetici dei prodotti.
Energia
C + D (prodotti)
E (energia assorbita)
A + B (reagenti)
tempo
Un esempio di reazione esoergonica è la combustione del metano in presenza di ossigeno.
1
CH4 + 2O2 CO2 + 2H2O + 212,8 kcal
Ogni mole di metano (16 g) che brucia libera 212,8 Kcal.
Un esempio di reazione endoergonica è la scissione dell'acqua in idrogeno ed ossigeno
2H2O + 136,6 Kcal 2H2 + O2
Per spezzare una mole di acqua (18 g) è necessario fornire 68,3 Kcal.
Legame covalente polare
Nella maggior parte dei casi gli atomi che formano legame covalente appartengono ad elementi
diversi che presentano una diversa forza di attrazione sugli elettroni di legame condivisi
(elettronegatività); la nube elettronica che costituisce il legame covalente risulta spostata
verso l'atomo più elettronegativo che acquista una carica parzialmente negativa (-), mentre
l'altro una carica parzialmente positiva (+). La distribuzione asimmetrica della nuvola
elettronica produce due poli aventi carica opposta ed il legame viene definito covalente
polare. Maggiore è la differenza di elettronegatività tra i due elementi, maggiore sarà la
polarità del legame. Quando la differenza di elettronegatività tra i due elementi supera il
valore critico di 1.9, l’elemento più elettronegativo è in grado di strappare l’elettrone all’altro
elemento ed il legame che si forma è ionico. Tutte le sostanze chimiche che presentano dei
dipoli si dicono polari, le altre apolari.
Legami intermolecolari
Le molecole polari sono soggette a deboli legami intermolecolari di reciproca attrazione
elettrostatica tra i poli opposti delle molecole, detti legami dipolo-dipolo. I legami Idrogeno
(ponti a Idrogeno) si presentano quando H si lega con legame covalente polare ad uno dei
tre elementi più elettronegativi della tabella periodica ( F O N). Gli atomi di idrogeno fanno da
ponte tra due atomi del gruppo FON, avendo con uno di questi un legame covalente polare e
con l’altro un legame elettrostatico.
I legami Idrogeno influenzano l'attività dell'acqua.
Specificità chimico-fisiche e importanza biologica dell'acqua
Le molecole d'acqua allo stato liquido tendono a disporsi in modo da presentare il polo negativo
(Ossigeno) di una rivolto verso il polo positivo (Idrogeno) di un'altra. Si formano così legami
intermolecolari di natura elettrostatica (legami idrogeno) che mantengono unite fra loro le
molecole.
2
Caratteristiche termiche dell'acqua e funzione termoregolatrice
L'acqua presenta un elevato calore specifico; ciò significa che, a parità di massa, l'acqua
necessita di una quantità di calore molto più elevata di qualsiasi altra sostanza inorganica per
manifestare lo stesso aumento di temperatura. L'acqua, rispetto ad altre sostanze, può
assorbire molto calore senza scaldarsi eccessivamente. Una conseguenza è che l'acqua si
scalda (e si raffredda) molto più lentamente delle altre sostanze. Per lo stesso motivo assorbe
molto calore durante il processo di evaporazione (elevato calore di evaporazione) perché il
calore fornito deve vincere le forze di attrazione tra le molecole particolarmente intense dei
legami idrogeno per riuscire a farle passare allo stato di vapore, abbandonando la superficie
del liquido. L'elevato calore specifico e di evaporazione fanno dell'acqua un ottimo
refrigerante e termoregolatore, capace cioè di ammortizzare in modo molto efficace
gli sbalzi termici, riducono le escursioni termiche, svolgendo un'azione mitigatrice sul clima. I
viventi (l'acqua è presente dal 50 al 95%) sono in grado di assorbire in modo efficace le
variazioni esterne di temperatura, molti eliminando calore in eccesso facendo evaporare acqua
(sudorazione). Il mantenimento dell'equilibrio termico è vitale per i viventi, in quanto molte
macromolecole sono termolabili e vengono distrutte ad elevate temperature, mentre a basse
temperature la velocità delle reazioni chimiche diminuisce al punto tale da essere incompatibile
con la vita.
Le soluzioni acquose
L'azione più importante è di agire da mezzo disperdente per le molecole consentendo alle
reazioni in soluzione di essere molto più veloci poiché i reagenti sono completamente mescolati
a livello molecolare ed hanno maggior probabilità di scontrarsi e quindi di reagire.
L'acqua scioglie solo le sostanze dotate di carica elettrica (ioniche e polari) per le quali
presenta grande affinità. La forte attrazione che l'acqua esercita su tali particelle (interazioni
idrofile) vince le forze interne di coesione per cui le molecole di soluto vengono staccate una
ad una, rivestite di molecole d'acqua (idratazione o solvatazione) e portate in soluzione.
L'acqua non scioglie invece le sostanze apolari (grassi) che vengono escluse dall'acqua, per cui
tendono a formare grossi aggregati tra di loro (interazioni idrofobiche). Nelle cellule le
interazioni idrofobiche hanno grande importanza soprattutto nel determinare la struttura delle
membrane cellulari costituite da sottili strati lipidici stabilizzati dalle repulsioni idrofobiche
che l'acqua esercita sia dall'interno che dall'esterno della cellula.
Glucidi (saccaridi)
- Monosaccaridi
I più diffusi sono quelli con sei (esosi) e con cinque (pentosi) atomi di carbonio. La presenza
di numerosi ossidrili (-OH) caratterizzati da elevata polarità, rende i monosaccaridi facilmente
solubili in acqua I monosaccaridi rappresentano il combustibile per eccellenza delle cellule.
L'energia è concentrata soprattutto nei legami C-H più energetici dei legami C-O.
- Disaccaridi e Polisaccaridi
I monosaccaridi possono legarsi tra loro facendo reagire 2 gruppi -OH con perdita di una
molecola d’acqua (reazione di condensazione), rimanendo uniti tramite un atomo di O.
3
Quando si uniscono due molecole di glucosio si forma il disaccaride maltosio. Sono disaccaridi
il saccarosio (glucosio+fruttosio) e il lattosio (glucosio+ galattosio).
La condensazione (polimerizzazione) tra molte molecole di monosaccaride origina lunghe
catene polimeriche, variamente ramificate (polisaccaridi). Alcuni polisaccaridi vengono
sintetizzati per immagazzinare riserve di zuccheri, altri hanno funzioni strutturali (costruzione).
Hanno funzione di riserva di energia l'amido (nelle piante) e il glicogeno (negli animali),
entrambi polimeri del glucosio. L'amido si trova abbondante nei semi e nei tuberi delle piante,
il glicogeno nei muscoli e nel fegato. I polisaccaridi di riserva, per essere utilizzati, devono
essere riconvertiti nei monosaccaridi costituenti. La reazione richiede l'introduzione di una
molecola d'acqua che spezza il ponte ossigeno riformando i due -OH. Tale processo è detto
idrolisi.
Nelle piante alcuni polisaccaridi presentano importanti funzioni strutturali. Il più importante è
la cellulosa, polimero del glucosio utilizzato dalle cellule vegetali per formare la parete
cellulare, rivestimento protettivo esterno alla membrana cellulare.
Lipidi
Caratteristica principale è l’insolubilità in acqua. Gran parte dei legami presenti sono legami
C-H che sono praticamente apolari e quindi idrofobici per cui un lipide in acqua forma
una fase separata in virtù delle interazioni idrofobiche. Per queste caratteristiche vengono
utilizzati come materiale isolante ed impermeabilizzante. Inoltre, poiché i legami C-H sono
particolarmente energetici, i lipidi costituiscono un'ottima riserva di energia, facilmente
accumulabile per la loro insolubilità. A parità di peso un lipide fornisce più del doppio di
energia rispetto ad un carboidrato (circa 9 kcal/g contro 3,75 kcal/g dei monosaccaridi e 4
kcal/g dei polisaccaridi o delle proteine).
Il contenuto di energia di un composto può essere descritto in base al suo grado di
ossidazione o di riduzione. Nella reazione di ossidazione un composto perde elettroni ed
energia mentre nella riduzione un composto acquista elettroni ed energia.
La maggior parte delle ossidazioni avviene per sostituzione di un legame C-H con un
legame C-O; possiamo giudicare il grado di ossidazione / riduzione di un composto organico
dal numero di tali legami. Il composto sarà tanto più ossidato quanto più numerosi sono i
legami C-O e tanto più ridotto quanto più numerosi sono i legami C-H.
Così un atomo di carbonio legato a 4 atomi di idrogeno a formare una molecola di metano
(CH4) è un composto molto energetico con un elevato grado di riduzione. Quando lo bruciamo
per estrarne l'energia effettuiamo un'ossidazione, sostituendo gli atomi di idrogeno con gli
atomi di ossigeno e ottenendo un composto estremamente ossidato e povero di energia,
l'anidride carbonica (CO2). La differenza di energia tra i due composti è quella che sfruttiamo.
Tra metano e CO2 esistono composti che presentano gradi di ossidazione intermedi:
metanolo (CH3OH), l'aldeide formica (metanale H-CHO), l'acido formico (H-COOH).
Ossidando l'acido formico si ottiene infine l'anidride carbonica.
Man mano che procede il processo ossidativo gli atomi di idrogeno vengono sostituiti da atomi
di ossigeno.
Gli acidi grassi sono costituiti da lunghe catene di C (fino 22) saturate da atomi di idrogeno.
Lipidi semplici: I trigliceridi sono caratterizzati dalla presenza dell'alcol glicerolo (glicerina);
I trigliceridi sono fondamentali costituendo i principali lipidi di riserva e di deposito.
4
Lipidi composti: Sono formati da lipidi semplici in associazione con altre molecole: lipoproteine, glicolipidi, in
particolare i fosfolipidi legati ad una molecola di acido fosforico (H3PO4). I fosfolipidi sono i costituenti
fondamentali delle membrane biologiche. La testa di gruppo fosfato è polare è idrofila e le
due code costituite dai residui degli acidi grassi sono apolari idrofobe.
L'eteropolarità permette ai fosfolipidi di organizzarsi spontaneamente in acqua formando
strutture ordinate. Le teste tendono a disporsi a contatto con l'acqua, mentre le code, respinte
dalle interazione idrofobiche, tendono a restare rivolte le une verso le altre. Si possono in tal
modo formare micelle sferoidali e lamine bistratificate.
Protidi
Le proteine strutturali hanno forma fibrosa, lunghe catene disposte in fasci a costituire le fibre
insolubili in acqua. Le proteine con funzione di regolazione hanno forma globulare con
catene strettamente compatte. Sono solubili in acqua.
Sono ottenute attraverso la combinazione di 20 amminoacidi che si legano tramite un legame
di condensazione (legame peptidico) facendo reagire il gruppo amminico di un amminoacido
con il gruppo carbossilico di un altro, con perdita di una molecola di acqua.
successione degli aa è diversa da proteina a proteina e definisce la specifica struttura
primaria di una proteine.
I filamenti proteici, rispondendo alle sollecitazioni prodotte dalle loro polarità interne si
ripiegano su se stessi formando strutture a diverso grado di complessità. La struttura
secondaria definita dai tipi di avvolgimenti (stabilizzati da ponti idrogeno) che interessano
tratti del filamento proteico (Eliche, Foglietti). La struttura terziaria è definita dal modo in cui il
filamento proteico si ripiega su se stesso in una conformazione compatta di tipo globulare
contenente le strutture secondarie. La struttura quaternaria si genera quando diverse molecole
proteiche in struttura terziaria globulare si saldano tra loro, assemblandosi.
La
Struttura secondaria
Non appena un filamento proteico viene sintetizzato passa in struttura secondaria attraverso
rotazioni permettono al filamento di avvolgersi su se stesso secondo schemi diversi,
raggiungendo una struttura finale stabile. L'alfa-elica è la conformazione secondaria più
5
"naturale" e più comune nelle proteine. Il filamento si avvolge a formare una spirale resa
stabile dai ponti idrogeno che si formano tra spira e spira.
Le proteine filamentose o fibrose
Le proteine fibrose, insolubili, hanno spesso un ruolo protettivo, connettivo o strutturale nei
viventi: cheratina (capelli, peli, unghie, corna), miosina (fibre muscolari), collagene che
per le sue eccezionali doti di resistenza alla trazione va a costituire gran parte del tessuto
connettivo (tendini, cartilagini, derma).
Struttura terziaria: le proteine globulari
La maggior parte delle proteine, dopo aver raggiunto la struttura secondaria subisce un
ulteriore processo di torsione avvolgendosi ulteriormente su se stesse formano una matassa
globulare compatta, caratteristica della struttura terziaria.
Gli enzimi sono catalizzatori biologici che aumentano la velocità con cui avviene la reazione.
Gli enzimi non si consumano durante la reazione, ma si ritrovano inalterati al termine di
ciascuna reazione, riconoscono in modo specifico il substrato (specificità).
Attraverso gli enzimi la cellula controlla quali reazioni devono essere eseguite e quali no.
Le proteine di trasporto (carriers) sono in grado di agganciare e trasportare specifiche
sostanze chimiche nel sangue e trasportano sostanze, altre sono immerse nella membrana
cellulare e permettono alla cellula di scambiare sostanze con l’esterno.
Struttura quaternaria
Alcune proteine nella loro forma attiva sono costituite dall'associazione di due o più unità in
struttura terziaria (subunità). Si parla in tal caso di struttura quaternaria.
L' emoglobina è un esempio di proteina in struttura quaternaria. La proteina è un tetramero
costituito dall'associazione di due catene (141 amminoacidi) e due catene β (146
amminoacidi). L’emoglobina è una proteina di trasporto in grado di trasferire i gas respiratori
dai polmoni ai tessuti e viceversa.
emoglobina
Esistono anche proteine in struttura quaternaria in cui le subunità si aggregano a formare
strutture filamentose che svolgono in genere funzioni strutturali. Ne sono tipici esempi l'actina
costituita da numerosi monomeri globulari associati a formare lunghi filamenti avvolti a spirale
fra loro.
6
Acidi nucleici: DNA ed RNA
La loro funzione è contenere l’informazione genetica e renderla disponibile per guidare il
metabolismo cellulare. Il “progetto genetico” scritto nel DNA è reso operativo tramite L’RNA.
Gli acidi nucleici si formano a partire da 4 tipi di nucleotidi.
Un nucleotide è formato dall’unione di uno zucchero pentoso che si lega ad una molecola di
acido fosforico H3PO4 (gruppo fosfato) e ad una base azotata con legami di condensazione.
Il DNA è costituito da 4 tipi di nucleotidi diversi da quelli che formano l’RNA
Nei 4 nucleotidi del DNA il pentoso è il desossiribosio, una molecola di ribosio con un atomo
di ossigeno in meno, mentre nell’RNA il pentoso è il Ribosio.
I 4 nucleotidi si differenziano tra di loro per le basi azotate che nel DNA sono:
Adenina (A)
Timina (T)
Citosina (C)
Guanina (G)
Le basi azotate dell’RNA sono:
Adenina (A)
Uracile (U)
Citosina (C)
Guanina (G)
I nucleotidi dell’RNA si differenziano da quelli del DNA solo per lo zucchero (ribosio) e per la
base Uracile che sostituisce la Timina.
I nucleotidi si legano tra loro in successione tramite legami tra il gruppo fosfato di un
nucleotide che si salda allo zucchero del nucleotide successivo in posizione 3. In questo modo
si forma una catena in cui si succedono zucchero e fosfato, mentre le basi azotate
rimangono sporgenti.
7
Il DNA presenta una struttura secondaria a doppia elica frutto dell’associazione di due
filamenti. L’accoppiamento delle due catene si realizza attraverso la formazione di ponti a
idrogeno tra le rispettive basi azotate che vengono a situarsi nella porzione centrale della
struttura. L’abbinamento fra i nucleotidi può realizzarsi soltanto tra basi complementari, cioè
fra adenina e timina, mediante due ponti a idrogeno (A=T), o tra citosina e guanina, mediante
tre ponti a idrogeno (C≡G).
Le basi presentano una struttura planare e sono disposte in maniera parallela tra loro come i
gradini di una scala a chiocciola.
La complementarietà dei due filamenti ha una conseguenza fondamentale. Stabilita l’esatta
successione di basi di un filamento, si può ricavare, semplicemente con la complementarietà,
la serie di nucleotidi del filamento corrispondente. Ciascuna coppia di basi dista dalle contigue
0,34 nm. Un giro completo dell’elica (passo) misura 3,4 nm, per cui sono presenti 10 coppie
di basi per un giro completo. Il diametro dell’elica è di 2nm. L’avvitamento è destrorso, ossia,
immaginando di guardare lungo l’asse, i due filamenti si avvolgono in senso orario.
8
Funzioni del DNA
Il DNA contiene il “progetto costruttivo” ed i “programmi di gestione e manutenzione” di un
organismo, codificati nella sequenza basi. Le sue funzioni sono:
1) Rendere disponibili le informazioni genetiche per la costruzione ed il corretto
funzionamento dell’organismo. Tale funzione viene mediata da molecole di RNA che
copiano le informazioni genetiche (trascrizione) e le trasformano (traduzione) in
proteine (sintesi proteica). Le diverse sequenze di nucleotidi, caratteristiche del DNA
di un organismo, definiscono quali amminoacidi debbano succedersi nella costituzione
delle proteine.
2) Generare copie delle informazioni genetiche (duplicazione del DNA) per rendere
disponibile l’intero progetto genetico per le nuove cellule. Ad ogni divisione cellulare
(mitosi) devono essere predisposte due copie del DNA in modo che ogni nuova cellula
possieda per intero tutte le informazioni per funzionare correttamente.
Duplicazione del DNA
Nel processo di duplicazione il doppio filamento viene attaccato e tagliato in un punto
dall’enzima topoisomerasi. La doppia elica viene aperta come una cerniera-lampo dall’enzima
elicasi che, rompendo i ponti ad idrogeno che tengono unite le basi azotate complementari,
separa i due filamenti e forma una struttura ad Y (forcella di replicazione) che viene
stabilizzata dalle proteine SSB. L’enzima DNA-polimerasi provvede ad agganciare su ogni
filamento esposto i nucleotidi complementari. In questo modo su ciascun filamento viene
ricostruito il filamento mancante e si generano due copie della doppia elica originaria. Poiché in
ciascuna di queste due copie rimane metà della molecola originaria, tale processo è anche noto
come sintesi semiconservativa.
La costruzione dei nuovi filamenti avviene solo in direzione 5'→3'. Ed avendo il DNA due
filamenti antiparalleli, la sintesi dei due nuovi filamenti avviene necessariamente in direzioni
9
opposte. Un filamento, detto filamento veloce (leading strand) viene sintetizzato in modo
continuo, poiché la DNA-Polimerasi avanza nella stessa direzione dell’elicasi.
L'altro filamento, detto filamento lento (lagging strand) viene sintetizzato in modo
discontinuo, con la formazione di segmenti di DNA detti frammenti di Okazaki, che vengono
successivamente saldati dall’enzima DNA-ligasi.
La DNA-polimerasi necessita di un innesco, una breve sequenza di RNA (RNA primer) da cui
partire per sintetizzare il nuovo filamento, dal momento che è in grado solo di aggiungere
nucleotidi a una catena preesistente. Inoltre la DNA-polimerasi possiede un secondo sito in
grado di controllare che l’attività di appaiamento delle basi sia avvenuta correttamente e di
correggere eventuali errori. Nel caso vengano individuati errori nel processo di duplicazione la
DNA-polimerasi torna indietro a correggerli (proofreading activity o correzione di bozze).
La duplicazione del DNA avviene contemporaneamente in più punti della molecola con le
forcelle di replicazione che si allontanano in direzioni opposte formando bolle di replicazione,
destinate ad estendersi ed a fondersi.
Mentre la bolle di replicazione crescono i filamenti di DNA non ancora aperti tendono a
manifestare una tensione torsionale in quanto aumenta il numero di spire per unità di
lunghezza. Le topoisomerasi risolvono il problema della tensione causato dall'avanzamento
dell’elicasi, tagliando periodicamente uno dei due filamenti e permettendo all’elica di girare,
per svolgere gli avvolgimenti in eccesso. Quando il DNA si è rilassato, la topoisomerasi
riconnette il filamento, ripristinando il DNA a doppia elica. I frammenti di Okazaki vengono
successivamente saldati dalla DNA ligasi, mentre i primers di RNA vengono rimossi e sostituiti
da analoghi filamenti di DNA da un enzima della famiglia delle DNA-polimerasi.
La struttura secondaria dell’RNA
L’RNA è costituito da una catena di nucleotidi in cui lo zucchero è il ribosio e la Timina è
sostituita dall’Uracile. La complementarietà delle basi è in questo caso Adenina-Uracile e
Citosina-Guanina. Mentre il DNA è una molecola di grandi dimensioni e notevolmente stabile,
gli RNA sono più piccoli, hanno durata limitata e sono caratterizzati da una struttura a
singolo filamento. Le molecole di RNA vengono sintetizzate utilizzando un filamento di DNA
stampo sul quale vengono appaiati i singoli nucleotidi complementari dell’RNA (trascrizione).
Enzimi specifici rimuovono alcune zone non codificanti della molecola (introni) e saldano
quelle codificanti (esoni) attraverso un processo di taglia-incolla detto splicing.
Funzioni biologiche dell’RNA
L’RNA permette al DNA di trasformare le informazioni in esso contenute in proteine. Esistono 3
tipi di RNA, tutti coinvolti nella sintesi proteica.
RNA ribosomiale (rRNA) che va a formare i ribosomi, organuli cellulari che
“traducono” le sequenze nucleotidiche in sequenze proteiche.
10
RNA messaggero (mRNA) che copia le informazioni contenute nel DNA
(trascrizione) e le trasferisce ai ribosomi per la traduzione.
RNA di trasporto o transfer (tRNA) che porta gli amminoacidi ai ribosomi affinché li
saldino in catene proteiche (sintesi proteica).
Sintesi proteica
Il DNA contiene le informazioni necessarie per posizionare nella giusta successione gli
amminoacidi di una proteina. Il tratto che “codifica” per una specifica proteina si definisce
“gene”. Nella sintesi proteica esistono due linguaggi: quello del DNA con un alfabeto di 4
lettere (basi azotate) e quello delle proteine con un alfabeto di 20 lettere ( amminoacidi).
Le informazioni devono pertanto essere “tradotte” da un linguaggio ad un altro. Per
effettuare la traduzione e “decifrare” un “messaggio codificato” è necessario possedere il
“codice” della corrispondenza tra i simboli dei due linguaggi.
Il “codice genetico” definisce il modo in cui la successione delle basi azotate del DNA deve
essere tradotta nella corretta successione di amminoacidi di una proteina.
Ciascun amminoacido viene “codificato” da una particolare tripletta di basi (codone).
Esistono più codoni che codificano per il medesimo amminoacido (ridondanza del codice), ma
ciascun codone non può codificare per amminoacidi diversi. Esistono anche codoni (notsense) che non codificano per alcun amminoacido; la cellula li utilizza per segnalare la fine
(stop) della sintesi della proteina.
Il processo di sintesi proteica si articola in due fasi: trascrizione e traduzione. Nella fase di
trascrizione l’informazione viene trasferita dal DNA all’RNAm, mentre nella fase di traduzione
l’informazione passa dall’RNAm alle proteine.
Trascrizione – sintesi dell’RNAm
L’informazione genetica viene copiata (trascritta) dal DNA in una molecola di mRNA. Il
processo avviene grazie all’enzima RNA-polimerasi. L’RNA-polimerasi I, che risiede nel
nucleolo, è responsabile della trascrizione dei geni per la produzione dell’rRNA). L’RNApolimerasi II è responsabile della sintesi del mRNA. L’RNA-polimerasi III sintetizza iltRNA.
Nella fase di inizio l’RNA-polimerasi si lega alla doppia catena del DNA, aprendola in
corrispondenza di una particolare sequenza (promotore) di nucleotidi situata all’inizio del
gene. Successivamente l’RNA-polimerasi scorre lungo il DNA rompendo i ponti Idrogeno tra le
basi azotate complementari ed aprendo la doppia elica. In questo modo una delle due catene
fa da stampo per la sintesi di una molecola di mRNA ad essa complementare. Mentre l’RNApolimerasi scorre sul filamento-stampo del DNA vengono agganciati ad esso i ribonucleotidi
complementari. Particolari sequenze di basi alla fine del gene (terminatore) segnaleranno il
termine della trascrizione. Il filamento di RNAm si stacca ed il DNA si richiude e si riavvolge. La
direzione di lettura del DNA è 3'→5' mentre quella di trascrizione è 5'→3'.
Il prodotto della trascrizione (trascritto primario) negli eucarioti subirà il processo di
maturazione. Le sequenze non codificanti (introni) durante il processo di splicing vengono
rimosse e avviene il ricongiungimento delle sequenze codificanti (esoni). Lo splicing avviene
grazie a un apparato enzimatico complesso (spliceosoma) in grado di riconoscere sequenze
11
specifiche presenti nelle zone di giunzione esone-introne, di rimuovere gli introni e di
ricongiungere correttamente tra loro i vari esoni. Una volta maturati gli mRNA passano nel
citoplasma per svolgere la loro funzione nella sintesi proteica.
Ogni mRNA è caratterizzato dal codone d’inizio AUG. I tre codoni UAA, UGA e UAG
rappresentano il segnale di terminazione della sintesi della catena polipeptidica.
L’RNA di trasporto (tRNA) trasferisce ai ribosomi i vari amminoacidi che, uniti tra loro con
legame peptidico, formano le proteine. I tRNA sono molecole piccole costituite da 75-80
nucleotidi che si appaiano tra loro in zone specifiche con ponti idrogeno tra basi
complementari. Tale situazione determina una caratteristica conformazione a “trifoglio”. Si
distinguono venti tRNA, ciascuno specifico per un determinato amminoacido.
La parte più caratteristica della molecola del tRNA è l’ansa terminale (anticodone) che porta
tre basi complementari ai codoni degli mRNA.
Gli RNA ribosomiali (rRNA) assemblati insieme a più di 50 diverse proteine, formano i
ribosomi che utilizzano le informazioni dell’RNAm e gli amminoacidi portati dagli RNAt per
assemblare le proteine. Sono costituiti da due subunità. Negli eucarioti i geni che codificano
per gli rRNA sono localizzati nel nucleolo.
12
Traduzione
Nella traduzione, l’informazione contenuta nell’RNAm come sequenza di codoni, viene tradotta
dai ribosomi e trasformata nella corrispondente sequenza di amminoacidi grazie al codice
genetico.
- Il filamento di mRNA si inserisce tra le due subunità del ribosoma che si posiziona sui primi
due codoni.
- Due molecole di tRNA, aventi gli anticodoni complementari si agganciano ai codoni,
posizionando in tal modo gli amminoacidi trasportati uno accanto all’altro. Gli amminoacidi
vengono uniti con legame peptidico ed il primo tRNA esce dal ribosoma lasciando il suo
amminoacido legato nella catena proteica in via di formazione
-Il ribosoma slitta sul filamento di mRNA, scalando di un codone e posizionandosi sul secondo e
terzo codone. Una terza molecola di tRNA si aggancia al terzo codone permettendo l’aggancio
del suo amminoacido ai due precedenti. Il secondo tRNA si sgancia lasciando il suo
amminoacido sulla catena proteica in via di formazione.
- La traduzione procede con il medesimo meccanismo, con il ribosoma che avanza di un codone
per volta ed un tRNA che entra carico del suo amminoacido ed uno che esce dopo aver
scaricato il suo amminoacido.
- Il processo termina quando il ribosoma trova un codone not-sense. Il filamento proteico si
stacca e viene liberato nel citoplasma dove assume la conformazione definitiva.
Un filamento di mRNA può essere tradotto più volte in modo da ottenere più copie della
medesima proteina. La traduzione può essere fatta contemporaneamente da più ribosomi che
si agganciano al medesimo filamento. La struttura che ne deriva prende il nome di
poliribosoma; ciascun ribosoma si trova in una fase diversa della sintesi della medesima
proteina
Derivati dei nucleotidi: ATP e NAD
ATP e NAD sono due nucleotidi che la cellula utilizza per il metabolismo energetico.
13
- L’ATP (Adenosin Trifosfato) viene utilizzato come contenitore e trasportatore di energia. Si
forma dalla condensazione di una molecola di ADP con una molecola di acido fosforico. La
reazione è fortemente endoergonica e richiede circa 7,3 kcal/mol (30,5 kJ/mol)
ADP + Pi + 7,3 kcal → ATP
L’ATP risulta così una molecola altamente energetica. L’energia è fissata nel legame tra il
secondo ed il terzo gruppo fosfato. La cellula utilizza l’energia estratta dai carboidrati e lipidi
per sintetizzare ATP. In questo modo parte dell’energia viene fissata come energia di legame in
molecole rapidamente utilizzabili per fornire energia.
La combustione di una molecola di glucosio permette alla cellula di caricare 36 ATP
C6H12O6 + 6O2 → 6CO2 + 6H2O + Energia (36 ATP)
Quando la cellula ha bisogno di energia può ottenerla rapidamente idrolizzando ATP
ATP → ADP + Pi + 7,3 kcal
Durante il metabolismo la molecola oscilla continuamente tra forma carica ATP e scarica ADP.
- Il NAD (Nicotinammide Adenin Dinucleotide) è un coenzima che catalizza reazioni di
ossidoriduzione,essendo in grado di trasferire elettroni (ed H) da una molecola che si ossida ad
una molecola che si riduce. Quando il NAD+ acquista elettroni da una molecola A che si
ossida, passa alla forma ridotta NADH, mentre quando cede elettroni ad una molecola B che
si riduce passa nella forma ossidata NAD+.
La riduzione richiede 2 elettroni, uno ione H+ e 52,4 kcal/mol (220 kj/mol)
NAD+ + 2e + H+ + 52,4 kcal → NADH
Anche il NAD oscilla tra la forma scarica NAD+ e la forma carica NADH.
Altre importanti ossidoreduttasi sono il FAD (Flavin Adenin Dinucleotide) e il NADP (NAD
fosfato) che esistono nella forma ossidata FAD ridotta FADH2. NADP+ ossidato e NADPH
ridotto.
Il FAD è in grado di assorbire un po’ meno energia del NAD
FAD + 2e + 2H+ + 46,0 kcal → FADH2
Citologia
La cellula procariote forma gli organismi unicellulari del regno monere. E’ relativamente
piccola (1 µm) ed ha struttura alquanto semplice. Gli unici organuli presenti sono i ribosomi. Il
DNA si trova libero nel citoplasma senza essere racchiuso da una membrana nucleare.
La cellula eucariote costituisce tutti gli altri organismi unicellulari e pluricellulari. La sua
dimensione è dell’ordine di 10 µm. Il volume è quindi mediamente 1000 volte più grande di
una cellula procariote. Il DNA è racchiuso da una membrana, formando così un particolare
organulo (nucleo). Possiede organuli deputati a svolgere una particolare funzione, che la
rendono enormemente più complessa.
Le dimensioni delle cellule si mantengono piccole per la necessità di mantenere elevata la loro
efficienza attraverso un ottimale rapporto con l’ambiente da cui la cellula trae le sostanze
nutritizie ed al quale cede le sostanze di rifiuto Quando la cellula si accresce, il volume
aumenta più rapidamente della superficie. Ciò determina uno squilibrio nutrizionale sempre
maggiore perché la superficie non riesce più a garantire scambi con l’ambiente, adeguati alla
massa cellulare.
Inoltre, in una cellula troppo grande le sostanze nutritive richiederebbero tempi troppo lunghi
per diffondere dalla superficie all’interno e altrettanto inefficace risulterebbe l’allontanamento
delle sostanze di rifiuto. La stessa attività di controllo metabolico da parte del nucleo
risulterebbe inefficiente oltre una certa distanza. Le cellule con intensa attività metabolica che
presentano rapidi scambi con l’ambiente, sono più piccole rispetto a cellule meno attive.
Esistono eccezioni: le cellule muscolari presentano una lunghezza di alcuni centimetri, ma
conservano l’efficienza mantenendo un buon rapporto superficie/volume grazie alla loro forma
14
molto allungata ed inoltre sono cellule polinucleate (ogni nucleo gestisce la regione di
citoplasma circostante).
La cellula eucariote animale
Membrana plasmatica
Racchiude il contenuto e costituisce una barriera fra l'ambiente intracellulare e quello
extracellulare. È costituita da un doppio strato continuo di fosfolipidi dello spessore di 8-10 nm,
attraversata da numerose proteine. È presente una piccola frazione di colesterolo che la
stabilizza. I fosfolipidi sono formati da una "testa" di glicerolo legato ad un gruppo fosfato e a
due "code" di acidi grassi. Come conseguenza della presenza di acqua sia all’interno che
all’esterno della cellula, i fosfolipidi rivolgono le teste polari verso le due superfici della
membrana, mentre le code apolari sono rivolte verso la parte interna del doppio strato
Sia le molecole lipidiche che quelle proteiche in essa immerse, sono in grado di muoversi
liberamente (mosaico fluido) consentendo alla membrana di autoripararsi, cioè richiudersi
rapidamente qualora si produca un foro, nonchè di fondersi e scindersi, senza che si abbiano
perdite di materiale cellulare, nel corso di processi di divisione, esocitosi, endocitosi.
Attraverso la membrana avvengono e vengono regolati tutti gli scambi di materiali ed
informazioni. Essa ha funzione di trasporto, interazione ed integrazione con l’ambiente.
Trasporto:
La membrana è una barriera selettivamente permeabile. Il doppio strato fosfolipidico
permette il libero passaggio solo dell'acqua, O2 - CO2 e piccole molecole liposolubili, mentre è
impermeabile per ioni e molecole idrosolubili; per questi materiali il trasporto viene effettuato
utilizzando particolari proteine trasportatrici transmembraniche (carriers) e/o attraverso
fenomeni di motilità della membrana (esocitosi ed endocitosi).
Le proteine trasportatrici sono in grado di riconoscere la sostanza da trasportare. Esistono
tre modalità di trasporto. Uniporto: una sola molecola in un unico verso; Simporto: due
molecole in un stesso verso. Antiporto: due molecole in versi opposti.
Le proteine trasportatrici si dividono in pompe e canali
1) Le pompe realizzano il trasporto attivo di molecole; il passaggio avviene con consumo di
energia (idrolisi ATP) contro il gradiente di concentrazione.
15
2) I canali operano il trasporto passivo: il passaggio avviene per diffusione senza consumo
di energia, secondo il gradiente di concentrazione, La diffusione è fenomeno fisico per il quale
un fluido (gas o liquido), come conseguenza del moto disordinato delle molecole che lo
costituiscono, tende a “sparpagliarsi” occupando tutto lo spazio disponibile. Ne consegue la
tendenza spontanea a muoversi (diffondere) da zone a maggior concentrazione verso zone a
minor concentrazione.
Le particelle si muovono in tutte le direzioni; alcune passeranno dalla regione A alla regione B
e viceversa. Ma poiché vi sono più particelle in A che in B vi sarà un maggior flusso da A verso
B, piuttosto che da B verso A. Finché le particelle non saranno distribuite omogeneamente vi
saranno più particelle che passano da A verso B che particelle che passano da B verso A. Ciò
genera diffusione che porta ad eliminare progressivamente la distribuzione non omogenea. I
canali funzionano solo se tra interno ed esterno della cellula esiste una differenza di
concentrazione.
La cellula è in grado di aprire o chiudere i canali a seconda delle necessità. Avvalendosi di
questi meccanismi di trasporto la cellula può mantenere la concentrazione interna delle
sostanze su valori diversi da quelli che caratterizzano l'ambiente esterno.
L’osmosi si manifesta quando una membrana selettivamente permeabile separa due soluzioni
a diversa concentrazione e solo le molecole dell’acqua possono diffondere liberamente, mentre
la diffusione del soluto è impedita. In queste condizioni l’acqua tende a diffondere dalla
soluzione più diluita (dove l’acqua è presente in quantità maggiore) alla soluzione più
concentrata (dove l’acqua è presente in quantità minore). Le cellule animali poste in
ambiente ipertonico (concentrazione maggiore) possono facilmente disidratarsi. Poste in
ambiente ipotonico (concentrazione minore) possono assorbire acqua fino alla lisi. I globuli
rossi devono essere immersi in un mezzo isotonico (plasma) per funzionare. Le cellule
vegetali rivestite di una robusta parete di cellulosa, mantengono il citosol ipertonico rispetto
all’ambiente, richiamando acqua per osmosi che determina una pressione (turgore cellulare)
che le mantiene gonfie. Ciò è fondamentale per mantenere distesi i tessuti vegetali che non
presentano strutture di sostegno efficaci (foglie).
Il processo di importazione di sostanze è detto endocitosi ed avviene grazie alla capacità della
cellula di avvolgere il materiale da introdurre con una piccola porzione di membrana che si
stacca (vescicola di endocitosi) all’interno del citosol. Si parla di endocitosi mediata da
recettori quando la sostanza che deve essere inglobate viene riconosciuta e legata da specifici
recettori situati sulla superficie della membrana. Se il materiale da introdurre è solido si parla
di fagocitosi, se è liquido di pinocitosi. La fagocitosi avviene grazie alla crescita di due
estroflessioni (pseudopodi) attorno al materiale da inglobare, che crescono fino ad avvolgere
16
il materiale e a fondersi alle sue spalle inglobandolo in una vescicola di fagocitosi. In questo
modo i macrofagi del nostro organismo inglobano e distruggono batteri ed eliminano cellule
morte e strutture estranee.
La pinocitosi si produce grazie alla formazione di minuscole introflessioni della membrana,
all’interno delle quali viene risucchiato il liquido da introdurre. La membrana successivamente
si richiude liberando nel citosol la vescicola di pinocitosi
Il processo di esportazione è detto esocitosi ed avviene grazie alla capacità della cellula di
avvolgere il materiale da espellere con una piccola porzione di membrana (vescicola di
esocitosi) che si stacca dai sistemi interni di membrana (apparato del Golgi) e si va a fondere
con la membrana plasmatica. In tal modo il contenuto delle vescicole di esocitosi viene versato
all’esterno.
La cellula utilizza l’esocitosi sia per eliminare sostanze di rifiuto (escrezione) ma anche per
effettuare la secrezione di sostanze utili (neurotrasmettitori, ormoni).
La membrana è il mezzo con cui la cellula si "fa riconoscere" dalle altre cellule; tramite
molecole (marcatori) riconosce segnali chimici, tramite (recettori) è “sensibile” agli stimoli.
Organuli della cellula eucariote
- Sistemi di membrane: complessa rete di membrane e vescicole costituita da reticolo
endoplasmatico, apparato di Golgi, lisosomi, perossisomi.
17
Reticolo endoplasmatico (RE)
Sistema di membrane costituito da una fitta rete di cisterne e tubuli tra loro interconnessi. Si
distingue un Reticolo endoplasmatico liscio (REL), privo di ribosomi ed un Reticolo
endoplasmatico rugoso (RER) caratterizzato da membrane cosparse di ribosomi.
I ribosomi sono responsabili della sintesi delle proteine destinate ad essere secrete dopo
essere state rielaborate (formazione di strutture quaternarie, glicosilate, glicoproteine). Le
molecole prodotte vengono espulse del RER attraverso vescicole, raggiungendo l’apparato di
Golgi. Il RER è particolarmente abbondante nelle cellule con attività secretrice (pancreas).
Il Reticolo endoplasmatico liscio ha il compito di detossificare sostanze dannose per
l'organismo (etanolo). E’ molto diffuso nelle cellule epatiche; inoltre è sede del metabolismo di
fosfolipidi, acidi grassi, steroidi.
Apparato di Golgi
E’ costituito da una pila di cisterne schiacciate, ciascuna indipendente dalle altre.
Le proteine sintetizzate e modificate nel Reticolo Endoplasmatico, e rilasciate in vescicole,
entrano nel Golgi dove subiscono ulteriori modifiche. Completata la trasformazione le sostanze
vengono rilasciate attraverso vescicole che si formano per gemmazione e possono essere
utilizzate all’interno della cellula, per processi digestivi (lisosomi) o essere secrete
esternamente (vescicole di esocitosi).
Lisosomi
Vescicole contenente enzimi idrolitici digestivi in grado di degradare molecole e strutture
interne alla cellula nei loro costituenti elementari che vengono poi riutilizzati o espulsi.
Attraverso questo processo i globuli bianchi digeriscono i microrganismi o cellule morte.
18
Perossisomi
Vescicolari presenti nelle cellule eucarioti, la cui funzione è l’attività ossidativa (respiratoria) di
acidi grassi, etanolo; consuma ossigeno (O2) e forma perossido di idrogeno (H2O2), molecola
altamente reattiva che può facilmente produrre radicali liberi tossici; i perossisomi lo
rimuovono attivamente usando l’enzima catalasi attraverso la reazione
2H2O2 → O2 + 2H2O
Preservando così le strutture cellulari dall’azione nociva del perossido di idrogeno.
Sistema citoscheletrico
Citoscheletro
Sistema di filamenti proteici: microtubuli, microfilamenti e filamenti intermedi. Nelle
cellule animali contribuisce a mantenere la struttura e la forma della cellula fornendo
un'impalcatura per l'organizzazione interna e un punto di ancoraggio per gli organuli.
19
Ribosomi
Presenti nel citosol oppure sul RER, sono composti da RNA ribosomiale e proteine e
sintetizzano le proteine. Se la cellula secerne le proteine prodotte, possiede solo ribosomi
attaccati al RER. Se immagazzina le proteine che sintetizza, possiede anche ribosomi liberi nel
citoplasma. Le due subunità del ribosoma sono sintetizzate nel nucleolo.
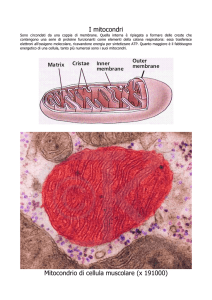
Mitocondri
Organuli addetti alla respirazione cellulare. Sono costituiti da due membrane, interna ed
esterna separate da uno spazio intermembrana. Lo spazio delimitato dalla membrana
interna è detto matrice mitocondriale. La membrana interna si estende nella matrice
formando delle pieghe (creste mitocondriali) dove si concentrano gli enzimi respiratori. La
membrana esterna è permeabile grazie alla presenza di canali proteici (porine), quella interna
presenta trasportatori altamente selettivi che la rendono poco permeabile.
La funzione più importante consiste nell'estrarre energia dai substrati organici (respirazione
cellulare) per produrre un gradiente protonico (H+) sfruttato per sintetizzare ATP.
Respirazione aerobica
Sistema di processi esoergonici di ossidoriduzione che implicano consumo di O2 e
formazione di CO2. Il substrato energetico (glucosio) viene ossidato ad anidride carbonica
(CO2) cedendo i propri elettroni all’ossigeno (O2) che si riduce ad acqua (H2O)
C6H12O6 + 6 O2 → 6 CO2 + 6H2O + Energia
La reazione globale è l’ossidazione completa del glucosio come in una normale combustione,
ma solo una parte dell'energia liberata viene trasformata in energia chimica sotto forma di
ATP. In totale si ottengono 38 ATP per ogni molecola di glucosio corrispondenti a (38 x 7.3)
277 Kcal/mol. L’efficienza del processo è di circa il 40% in quanto la reazione di combustione
libera 686 Kcal/mol (277/686 ≈ 0.40).
Viene riassunta in tre stadi: glicolisi, ciclo di Krebs e fosforilazione ossidativa. Il primo
avviene nel citoplasma, gli ultimi due nei mitocondri, il ciclo di Krebs nella matrice, la
fosforilazione ossidativa nella membrana interna (creste).
- Glicolisi
Via metabolica universale, è costituita da una sequenza di 10 reazioni che avvengono nel
citoplasma e degradano il glucosio a due molecole di acido piruvico (piruvato) senza consumo
20
di ossigeno, producendo 2 ATP e 2 NADH. La glicolisi carica 4 ATP, ma deve essere
inizialmente attivata consumando 2 ATP per cui il guadagno netto è 2 ATP.
L’acido piruvico viene poi ossidato con formazione di una molecola di CO2 e un gruppo acetile
(CH3CO-). L’energia e gli elettroni che si liberano dal processo caricano 1 NADH (poichè il
glucosio ha prodotto 2 piruvati, vengono caricati 2 NADH). Il gruppo acetile viene agganciato
da un coenzima A (CoA) con formazione dell’acetil-CoA che trasporta il gruppo acetile
all’interno del ciclo di Krebs per completare il processo ossidativo.
- Ciclo di krebs (ciclo dell’acido citrico)
Serie di reazioni, che avvengono nella matrice, che ossidano completamente i due C del gruppo
acetile, formando due molecole di CO2. Durante il processo si libera energia, elettroni e ioni H+
che vanno a caricare 3 NADH, 1 FADH2 e 1 ATP. Poiché una molecola di glucosio fornisce al
ciclo 2 gruppi acetile, il guadagno ottenuto per molecola di glucosio sarà di 6 NADH, 2 FADH2
e 2 ATP. Se sommiamo a questo le molecole caricate in precedenza si ottengono:
10 NADH
2 FADH2
4 ATP
NADH e FADH2 rappresentano accettori temporanei di elettroni che devono scaricarsi e
cedere energia ed elettroni ad un accettore finale permanente.
- Fosforilazione ossidativa
NADH e FADH2 si scaricano cedendo elettroni e ioni H+ all’ossigeno che, riducendosi ad acqua,
costituisce l’accettore finale e permanente di elettroni. Il trasferimento di elettroni non avviene
direttamente, ma attraverso una serie intermedi (citocromi) che costituiscono una di catena
di trasporto degli elettroni (catena respiratoria). In questo modo l’energia viene liberata
in piccole quantità ad ogni passaggio e può essere utilizzata per caricare ATP.
Ogni NADH che si scarica nella catena respiratoria è in grado di caricare 3 ATP
Ogni FADH2 che si scarica nella catena respiratoria è in grado di caricare 2 ATP
I 10 NADH caricheranno quindi 30 ATP, i 2 FADH2 caricheranno altri 4 ATP, per un totale di 34
ATP che, sommati ai 4 ATP già ottenuti portano il guadagno totale della respirazione
aerobica a 38 ATP. In alcune cellule il trasporto all’interno dei mitocondri dei 2 NADH formati
durante la glicolisi richiede un consumo di 2 ATP. In tal caso la resa finale sarà di 36 ATP. La
catena respiratoria è costituita da quattro complessi redox; gli elettroni vengono trasferiti da
un gruppo al successivo e durante il passaggio sono in grado di generare un flusso di ioni H +
dalla matrice verso lo spazio intermembrana (pompe protoniche). In questo modo si genera
ai due lati della membrana delle creste una differenza di concentrazione in ioni H+ (gradiente
elettrochimico) sotto la cui spinta gli ioni H+ rientrano nella matrice utilizzando l’ATPsintetasi, un enzima transmembranico in grado di convertire il flusso di ioni H + in energia
chimica per la sintesi di ATP (teoria chemiosmotica).
21
L’intero processo può essere così schematizzato:
Respirazione aerobica - schema
22
Vie metaboliche anaerobiche
Sono le vie di demolizione dei substrati energetici in cui l’accettore finale di elettroni non è
l’ossigeno; molto meno efficienti, dal punto di vista energetico, ma sufficienti a garantire la
sopravvivenza di molti microrganismi. Utilizzano la glicolisi come tappa iniziale.
- Fermentazione lattica
I 2 NADH che si formano dalla glicolisi si scaricano sul piruvato, riducendolo ad acido lattico
(CH3-CHOH-COOH). Il guadagno energetico rimane 2 ATP formati durante la glicolisi. La
fermentazione lattica è alla base della trasformazione del latte in yogurt grazie all’attività di
particolari batteri. Può avvenire anche nei nostri muscoli, quando il lavoro è particolarmente
intenso e l’apporto di ossigeno non è sufficiente a soddisfare le richieste energetiche. In queste
condizioni la glicolisi può essere accelerata (non richiedendo ossigeno), ma l’eccesso di
piruvato che si forma e che non può essere assorbito dal ciclo di Krebs, viene
temporaneamente convertito in acido lattico. L’acidità che si genera nelle cellule muscolari è
responsabile dell’indolenzimento muscolare e può provocare crampi.
- Fermentazione alcolica
I 2 NADH che si formano dalla glicolisi si scaricano sul piruvato che viene però anche
decarbossilato, liberando una molecola di CO2 e formando alcol etilico. Il guadagno energetico
è sempre di 2 ATP. La fermentazione alcolica è alla base della produzione di bevande
alcoliche fermentate come vino, birra, sidro ad opera dei lieviti. La liberazione di CO2 permette
di ottenere fermentati gasati (birra, spumante). La fermentazione alcolica viene utilizzata
anche nei processi di lievitazione durante i quali i carboidrati contenuti nella farina vengono
trasformati in etanolo e CO2 che si libera nell’impasto gonfiandolo e rendendolo spugnoso.
Nucleo
Contiene il DNA, trasmette i caratteri ereditari (duplicazione DNA) e controlla il metabolismo
cellulare mediante la trascrizione dei geni ed il controllo della sintesi proteica. La membrana
nucleare è interrotta da pori attraverso i quali avviene il trasporto di grosse molecole. In
materiale per la forte affinità con i coloranti basici è chiamato cromatina. Sono inoltre
presenti corpi fortemente basofili a causa dell’abbondanza di RNA (nucleoli), regioni deputate
alla trascrizione dell’RNA ribosomiale e al successivo assemblaggio dei ribosomi.
23
La cromatina è costituita da DNA e proteine nucleari (istoni) e non-istoniche. Può
manifestarsi in una forma diversamente condensata in conseguenza del diverso grado di
compattazione del DNA attorno agli istoni. Gli istoni sono piccole proteine la cui funzione è di
organizzare il DNA compattandolo in modo da conservarlo in un volume ridotto. Nel primo
livello di compattazione il DNA si avvolge attorno ad un ‘rocchetto’ di 8 istoni (nucleosoma).
Tra un nucleosoma e l'altro, è presente un filamento di DNA clinker che dà alla cromatina
l’aspetto di una “collana di perle”.
In questo stato la cromatina decondensata (despiralizzata) prende il nome di eucromatina
che è trascrizionalmente attiva cioè il DNA è accessibile per la trascrizione dell’informazione
genetica e la sintesi di mRNA. Ogni nucleosoma è associato all’istone H1 che compatta i
nucleosomi. Nel secondo livello di condensazione i nucleosomi formano una fibra da 30 nm di
diametro. Questi livelli di condensazione sono trascrizionalmente inattivi (il DNA non può
essere trascritto) e sono noti con il termine eterocromatina.
24
Le fibre da 30 nm possono ripiegarsi su se stesse in anse, stabilizzate da una impalcatura di
proteine non-istoniche, che raggiungono il grado finale di compattazione condensandosi in
cromosomi.
I cromosomi compaiono solo al momento della divisione cellulare (mitosi e meiosi) in quanto
la condensazione della cromatina in cromosomi rende più agevole l’ordinata ripartizione del
materiale genetico. Il cromosoma presenta una forma bastoncellare costituita da due filamenti
identici di DNA (cromatidi fratelli) uniti a livello di una strozzatura detta centromero a
ciascuno dei quali è associata una placca proteica (cinetocore) che serve come punto di
attacco per le fibre del fuso mitotico.
Le estremità di un cromosoma (telomeri) sono strutture marcatempo che conteggiano il
numero di volte in cui una cellula può riprodursi. Le replicazioni cessano dopo 50-70 divisioni.
Il telomero subisce un accorciamento progressivo ogni volta che i cromosomi vengono duplicati
in fase S della divisione cellulare (orologio mitotico). Dopo un numero calcolabile di
25
duplicazioni (50-70 nelle cellule umane) la crescita cellulare si blocca, per passare da una fase
di senescenza ad una fase di crisi, la cellula entra in apoptosi (morte programmata).
Solo il 25% del menoma umano è costituito da DNA genico (30.000 geni) e solo l’1,5% è
DNA codificante. Dunque circa il 98.5% del genoma umano è composto da DNA non
codificante.
Numero e struttura dei cromosomi costituiscono il cariotipo, ben evidenziabile (e
fotografabile) durante la divisione cellulare.
Tutte le cellule di un organismo possiedono lo stesso menoma, tuttavia ogni cellula si
specializza in determinate funzioni utilizzando solo parte delle informazioni contenute nel suo
DNA (esprime solo certe proteine). Per ogni cellula inoltre, tipo e quantità di proteine prodotte
dipendono anche dalle necessità in un determinato istante. Le cellule devono dunque
possedere meccanismi che permettano loro di esprimere certi geni e silenziarne altri. Accanto
ai geni strutturali destinati ad essere trascritti in mRNA e ad esprimere la proteina, è
presente un promotore costituito da una particolare sequenza di DNA che indica dove deve
iniziare la trascrizione; è la zona di riconoscimento e di attacco per l’RNA polimerasi che deve
sintetizzare l’mRNA. La sua azione è sotto il controllo di un gene regolatore in grado di
bloccare o sbloccare la trascrizione del gene in funzione delle esigenze.
Cellula eucariote vegetale
Non presenta alcune strutture della cellula animale: centrioli e corpi basali, ciglia e flagelli.
Possiede però alcune strutture tipiche: parete, vacuolo e plastidi.
- Parete cellulare
Costituisce uno strato rigido e robusto esterno alla membrana. È composta da sottili filamenti
di cellulosa che, unendosi, formano filamenti più spessi. Altro costituente è la lignina che le
conferisce rigidità.
- Vacuolo
Cavità piena di succo vacuolare, occupa gran parte del volume cellulare ed è delimitata da
una membrana (tonoplasto). Costituisce un idroscheletro che, esercitando pressione contro la
superficie della cellula racchiusa dalla parete, produce sostegno (turgore). All'interno del
vacuolo possono essere immagazzinate sostanze di riserva, prodotti intermedi di processi e
scorie, deposito per sostanze nocive o repellenti come difesa nei confronti degli animali.
- Cloroplasto
Organulo verde per la presenza di clorofilla, è sede della fotosintesi clorofilliana. Presenta
dei ripiegamenti della membrana interna a forma di minuscoli sacculi appiattiti (tilacoidi) che
ospitano i pigmenti fotosintetici e si sovrappongono costituendo una pila (grano).
La fotosintesi è un processo di ossidoriduzione nel corso del quale l'energia luminosa viene
trasformata in energia chimica e l'anidride carbonica viene trasformata in glucosio. La reazione
globale è opposta alla reazione di respirazione aerobica.
6 CO2 + 6H2O + Energia → C6H12O6 + O2
Il Carbonio viene ridotto da anidride carbonica a glucosio. Gli elettroni per la riduzione vengono
forniti dall’acqua che si ossida ad ossigeno; l’energia proviene direttamente dalla
radiazione luminosa ed è catturata dai pigmenti fotosintetici.
26
Gli zuccheri semplici possono essere immediatamente utilizzati nella respirazione cellulare
come fonte di energia, o immagazzinati come amido accumulato nelle radici o tuberi per essere
utilizzato successivamente,. Poiché gli organismi fotosintetici sono in grado di sintetizzare le
molecole alimentari sono detti autotrofi mentre gli animali sono eterotrofi in quanto non
sono autonomi dal punto di vista alimentare.
Nello schema generale della fotosintesi si possono distinguere due fasi: fase lucedipendente, fotochimica, localizzata nelle membrane dei tilacoidi, fase non luce
dipendente, chimica, localizzata nello stroma. La fase fotochimica consiste nella cattura da
parte dei pigmenti fotosintetici della radiazione luminosa e sua trasformazione in legami ad
alta energia dell’ATP e la riduzione di NADP+ a NADPH. L’acqua viene ossidata ad ossigeno
per fornire gli elettroni necessari alla riduzione del NADP+ e viene liberato ossigeno gassoso
O2. ATP e NADPH forniscono l'energia e il potere riducente necessari per la riduzione della
CO2 a glucosio vel ciclo di Calvin.
- Fase luce-dipendente
I pigmenti organizzati in due strutture (fotosistema I e fotosistema II) contengono, oltre
alla clorofilla, pigmenti accessori (carotenoidi) in grado di assorbire una particolare e
differente lunghezza d'onda della luce. In questo modo ogni pigmento si comporta come
un’antenna in grado di assorbire particolari radiazioni luminose. La struttura della clorofilla è
tale per cui assorbe le lunghezze d'onda blu e rosso, mentre riflette il verde. Le molecole che
hanno assorbito energia e si trovano in uno stato eccitato, instabile che le porterà a riemettere
tale energia. I fotosistemi sono organizzati in modo da assorbire la radiazione luminosa e
trasferirla ad una molecola di clorofilla (centro di reazione) del fotosistema. L’energia così si
concentra (imbuto energetico) ed è sufficiente a strappare un elettrone al centro di reazione,
provocando la sua ossidazione con cui cede elettroni ad una molecola (accettore primario).
Il fotosistema I (P 700) ha il massimo assorbimento a lunghezza d'onda 700 nm. Il
fotosistema II (P 680) a 680 nm. I due fotosistemi lavorano in modo integrato generando un
flusso di elettroni, alimentato dall’energia luminosa, che li percorre partendo dall’acqua fino a
raggiungere il NADPH. Il fotosistema II lavora ad un livello energetico leggermente inferiore
rispetto a fotosistema I.
1) Gli elettroni del centro di reazione del fotosistema II, spinti dai fotoni assorbiti vengono
captati dall’accettore primario del fotosistema (plastochinone).
27
2) Gli elettroni vengono successivamente ceduti dall’accettore primario attraverso una catena
di trasporto degli elettroni, al centro di reazione del fotosistema I che si trova ad un livello
energetico inferiore. Durante questa discesa si libera energia utilizzata i parte per sintetizzare
ATP (fotofosforilazione).
3) Gli elettroni giunti al centro di razione del fotosistema I, spinti dai fotoni assorbiti, vengono
captati dall’accettore primario del fotosistema I e da qui vengono ceduti al NADP+ che si riduce
a NADPH che rappresente il potere riducente.
4) Il centro di reazione del fotosistema II viene continuamente rifornito di elettroni dalla
reazione di fotolisi dell'acqua che in presenza di luce viene ossidata ad ossigeno molecolare.
Il meccanismo che accoppia la sintesi di ATP con il trasporto degli elettroni viene spiegato con
l’ipotesi chemiosmotica di Mitchell. La produzione di ATP è accoppiata alla formazione di un
gradiente di protoni (H+). Alcune proteine (pompe protoniche) pompano ioni H+ nel lume dei
tilacoidi con formazione di un gradiente elettrochimico. La sintesi di ATP avviene come
conseguenza del ritorno dei protoni nello stroma, in risposta al gradiente generato, attraverso
le ATP-sintetasi situate nelle membrane dei tilacoidi.
- Ciclo di Calvin-Benson
Si svolge nello stroma dove ATP e NADPH vengono impiegati per ridurre la CO2 a glucosio
tramite una serie di reazioni (ciclo di Calvin-Benson). Ad ogni ciclo una molecola di CO2 si
combina con uno zucchero a 5C, il ribulosio-1,5-difosfato (RuDP), per formare due molecole di
un composto a 3C (PGA). Il PGA viene ridotto dal NADPH e fosforilato dall’ATP trasformandosi
in un composto a 3C (PGAL). Dopo sei cicli, ciascuno dei quali consuma una molecola di CO2,
due di NADPH e tre di ATP, vengono prodotte 12 molecole di PGAL, due delle quali si
combinano a formare una molecola a 6C (glucosio), mentre le rimanenti rigenerano il RuDP. Le
reazioni della fase oscura possono essere così riassunte:
6 CO2 + 12 NADPH + 12H+ + 18 ATP
12 NADP+ + C6H12O6 + 6 H2O
→
28
Divisione cellulare
La mitosi è la divisione cellulare in cui le cellule figlie presentano il medesimo corredo
cromosomico diploide (2n) della cellula madre. Le due cellule sono geneticamente identiche
(clonazione). Gli organismi unicellulari utilizzano questo processo per riprodursi, i
pluricellulari per aumentare il numero delle loro cellule (sviluppo ed accrescimento) e per
sostituire le cellule morte ed invecchiate. Tutte le cellule di un organismo pluricellulare
derivano da un'unica cellula (zigote) che si forma dalla fecondazione di un gamete femminile
(uovo) con un gamete maschile (spermatozoo). Lo zigote genera poi tutte le cellule somatiche
attraverso continue mitosi.
La meiosi è la divisione cellulare in cui le cellule figlie presentano un corredo cromosomico
dimezzato (aploide) rispetto alla cellula madre e sono tra loro geneticamente diverse. Viene
utilizzata dai pluricellulari per creare le cellule riproduttive (gameti) che dovranno fecondarsi
per rigenerare lo zigote diploide. La meiosi ha il compito di creare “variabilità genetica”,
producendo cellule aploidi geneticamente una diversa dall’altra in modo che i nuovi individui
siano geneticamente diversi.
Mitosi e ciclo cellulare
L’intervallo di tempo tra due mitosi è definito interfase ed occupa la maggior parte del ciclo
vitale della cellula. Il succedersi di interfase e fase mitotica definisce il ciclo cellulare.
L’interfase si suddivide nelle fasi G1, S e G2.
La fase G1 (growth=crescita) è molto più lunga delle altre in quanto la cellula cresce
aumentando dimensioni e numero degli organuli. Durante la fase S (S=sintesi) si ha la
replicazione del DNA. La fase G2, relativamente corta, rappresenta il periodo in cui la cellula si
prepara alla divisione mitotica.
La mitosi viene suddivisa in: profase, metafase, anafase e telofase, ciascuna riconoscibile al
microscopio ottico. L’apparato mitotico necessario, comprende un fuso di microtubuli
longitudinali, tessuti tra due centrioli ai due poli della cellula.
- Profase (formazione cromosomi e fuso mitotico, scomparsa membrana nucleare)
La cromatina si condensa a formare i cromosomi, ciascuno costituito da due cromatidi che
diventano visibili. I nucleoli e la membrana nucleare scompaiono. I centrioli migrano ai poli
opposti della cellula e separandosi allungano progressivamente i microtubuli per l’aggiunta di
subunità di tubulina formando il fuso.
- Metafase (formazione piastra equatoriale)
I cromosomi si agganciano alle fibre del fuso mediante il cinetocore in corrispondenza del
centromero, la struttura che mantiene legati insieme i due cromatidi. I cromosomi si
dispongono nella parte mediana del fuso formando la piastra equatoriale o metafasica.
- Anafase (separazione e migrazione cromatidi)
Separazione del centromero, i cromatidi vengono così tirati dai tubuli connessi al cinetocore,
verso le estremità opposte del fuso. Ciò permette un’esatta divisione del materiale genetico. Al
termine dell’anafase, due gruppi di cromosomi identici sono ammassati ai poli opposti.
- Telofase (scomparsa cromosomi e fuso, formazione membrana nucleare, citodieresi)
I cromosomi si despiralizzano in cromatina. Intorno ai due nuovi complessi cromosomici
ricompaiono le membrane nucleari, il fuso mitotico si disassembla. La telofase si conclude con
la citodieresi (citocinesi) con cui la cellula si divide nelle due cellule figlie.
29
La citodieresi avviene con modalità diverse nelle cellule animali e vegetali. Nella cellula
animale la membrana plasmatica si introflette in modo da formare un solco equatoriale che
circonda la cellula strozzandola progressivamente, finché non si divide in due cellule figlie. Un
anello di microfilamenti (actina) è presente sotto il solco di divisione e la citodieresi avviene
per contrazione di questo anello.
Nella cellula vegetale l’apparato di Golgi produce vescicole contenenti polisaccaridi che si
vanno a depositare sulla linea mediana, si fondono e formano una parete (piastra cellulare).
Quando questa lamella è completata ogni cellula costruisce la membrana cellulare, utilizzando
le membrane delle vescicole che si sono fuse.
- Meiosi
Processo mediante il quale una cellula diploide dà origine a quattro cellule apolidi (gameti).
Prima divisione
Determina la formazione di due cellule figlie
aploidi. Durante la profase I i cromosomi
omologhi
si
appaiono
e
si
uniscono
longitudinalmente (sinapsi). I cromatidi
iniziano a separarsi rimanendo incrociati in
punti (chiasmi) formando strutture a 4
cromatidi (tetradi). In questo modo gli
omologhi
si
scambiano tratti di DNA
(crossing-over). Quando si separeranno,
alcuni geni del cromosoma paterno si saranno
spostati su quello materno e viceversa, con
una ricombinazione del patrimonio genetico. Il crossing-over crea variabilità e consente ai
gameti aploidi di essere geneticamente diversi uno dall’altro. Durante l’anafase I gli omologhi
si separano e migrano verso i poli opposti del fuso (i cromatidi rimangono uniti). La
30
separazione avviene in modo casuale e ciascuna coppia si separa in modo indipendente dalle
altre (assortimento indipendente). In questo modo le cellule aploidi umane che si creano
alla fine della prima divisione meiotica possono presentare 2 23 diverse combinazioni di
cromosomi materni e paterni.
Seconda divisione
Ciascuna cellula aploide generata dalla meiosi I si divide formando due cellule aploidi. Questo
processo è molto simile alla mitosi in quanto durante l’anafase II ciascun cromosoma si separa
nei due cromatidi che migrano poi ai poli opposti della cellula.
Genetica classica
Alla base della genetica c'è il concetto di gene, un tratto di DNA che controlla la
manifestazione di un carattere codificando per una particolare proteina. Un cromosoma è
un’ordinata successione di geni. Un gene può presentarsi con forme varianti (alleli). Nelle
cellule diploidi che contengono due serie di cromosomi omologhi (2n), ciascun gene è portato
sia dall’omologo materno che dall’omologo paterno. In un individuo diploide il carattere si
presenta sempre con due alleli che occupano la medesima posizione (locus). I due alleli che
controllano un particolare carattere potranno essere uguali o diversi. Un individuo i cui due
alleli per la determinazione di un carattere sono uguali si dice omozigote; se i due alleli sono
diversi si dice eterozigote. La particolare combinazione di alleli che controlla un carattere si
definisce genotipo. La manifestazione osservabile di un carattere si definisce fenotipo. Nel
caso di omozigoti il fenotipo riflette l’unica informazione esistente nel genotipo. Nel caso di
eterozigoti il genotipo può manifestarsi con fenotipi differenti a seconda del tipo di interazione
che si produce tra i due alleli diversi. In particolare si possono presentare tre tipi di relazione:
dominanza completa, dominanza incompleta e codominanza.
- dominanza completa: uno dei due alleli prevale (dominante) ed è l’unico a manifestarsi
determinando il fenotipo. L’allele che rimane latente è detto recessivo. Per convenzione l’allele
dominante viene indicato con una lettera maiuscola (A) e l’allele recessivo con la
corrispondente minuscola (a). L'allele dominante si esprime sempre. Ciò comporta che gli
eterozigoti siano indistinguibili dagli omozigoti dominanti, avendo lo stesso fenotipo. La
comparsa del fenotipo recessivo, invece, identifica senza ambiguità un omozigote recessivo.
- dominanza incompleta: il fenotipo dell'eterozigote è una mescolanza tra il fenotipo
dell'omozigote dominante e quello dell'omozigote recessivo. Dal fenotipo si può sempre risalire
al genotipo. Il colore del fiore della bocca di leone può essere rosso (omozigote dominante RR),
rosa (eterozigote Rr) o bianco (omozigote recessivo rr).
- codominanza: l’eterozigote esprime il fenotipo di entrambi gli alleli, non mescolati, ma
separati. Sono esempi il colore pezzato del mantello di molti mammiferi ed i sistemi di gruppo
sanguigno AB0.
Mendel: le leggi dell’ereditarietà
Mendel si occupò di caratteri che si presentavano nel fenotipo ben distinti e non ammettevano
forme intermedie (caratteri discreti). La pianta di pisello (Pisum sativum) risultò
particolarmente adatta perché di facile coltivazione, può dare più generazioni all’anno,
presenta molti caratteri discreti, si riproduce per autofecondazione.
Aal 1854 al 1864 Mendel coltivò e analizzò almeno 28.000 piante, prendendo in considerazione
sette coppie di caratteri del seme e della pianta quali laltezza, posizione dei fiori, forma e
colore dei semi. Mendel classificò e contò gli esemplari ottenuti in ciascuna generazione,
cercando di individuare l’esistenza di precisi rapporti matematici tra i diversi individui di ogni
generazione.
- Legge della dominanza
Mendel incrociò piante di pisello che da molte generazioni producevano solo fiori violetti con
piante che da molte generazioni producevano solo fiori bianchi. Il risultato era sempre il
medesimo: in F1 gli ibridi presentavano sempre fiori violetti. I risultati possono essere riassunti
nella legge della dominanza o dell’uniformità degli ibridi F1: l’incrocio tra un omozigote
dominante (linea pura dominante) ed un omozigote recessivo (linea pura recessiva) genera una
generazione F1 di eterozigoti (ibridi) fenotipicamente uguali al genitore dominante.
- Legge della segregazione
In una successiva serie di esperimenti Mendel incrociò tra loro gli ibridi F1. La seconda
generazione filiale (F2) risultò composta per il 75% di individui fenotipicamente dominanti e
per il 25% di individui fenotipicamente recessivi (rapporto 3:1).
31
Frequenza e probabilità (quadrato di Punnett)
I risultati di un incrocio si misurano dalle frequenze con cui compaiono determinati caratteri. La
frequenza è il rapporto tra il numero di volte che un determinato evento si verifica ed il
numero totale di prove effettuate. La probabilità è invece il rapporto tra il numero di eventi
favorevoli ed il numero di eventi possibili. La frequenza si avvicina tanto più al valore della sua
probabilità quanto più elevato è il numero di eventi osservati (è più probabile che la frequenza
con cui esce ‘testa’ si avvicini al 50% se effettuo 1000 lanci piuttosto che se ne effettuo 10).
Per questo motivo, per stimare la probabilità che da un incrocio si generi un particolare
carattere piuttosto che un altro, dobbiamo incrociare un numero molto elevato di individui.
Conoscendo i genotipi degli individui che si incrociano è possibile calcolare le probabilità dei
genotipi risultanti senza effettuare l’incrocio, simulando gli eventi che portano alla formazione
dei nuovi individui (meiosi e fecondazione).
Si calcolano le probabilità che durante la meiosi gli alleli si trovino su di un gamete e
successivamente si calcolano le probabilità che i gameti femminili si ‘incontrino’ con i gameti
maschili. Il quadrato di Punnett è una tabella a doppia entrata che serve a simulare tutte le
possibili combinazioni dei gameti maschili con quelli femminili (fecondazione) e a calcolare la
probabilità dei genotipi che si generano come conseguenza. I gameti maschili (con le loro
probabilità) vengono posti ad un lato del quadrato ed i gameti femminili (con le loro
probabilità) vengono posti su di un lato adiacente. All’interno del quadrato vengono effettuate
tutte le combinazioni. La probabilità di un evento composto è il prodotto delle probabilità
degli eventi semplici. Così le probabilità dei genotipi si calcolano come prodotto delle
probabilità dei singoli gameti che devono combinarsi per dare quel genotipo. Ogni individuo
eterozigote ha una probabilità del 50% (0,5) di produrre gameti con l’allele dominante e del
50% (0,5) di produrre gameti con l’allele recessivo. Ora costruiamo all’interno del quadrato le
4 possibili combinazioni che sono i 4 genotipi che possono prodursi dall’incrocio. Ognuno di essi
avrà una probabilità di formarsi del 25% (0,5 x 0,5 = 0,25). Dall’incrocio di due ibridi si
possono formare dunque un 25% di omozigoti dominanti (PP), un 50% di eterozigoti (Pp) ed
un 25 % di omozigoti recessivi (pp). Essendo gli omozigoti dominanti e gli eterozigoti
fenotipicamente uguali, l’incrocio produce il 75% di individui fenotipicamente dominanti ed il
25% di individui recessivi. I risultati di questa seconda serie di incroci possono essere riassunti
nella legge della segregazione o legge della disgiunzione: l’incrocio tra due eterozigoti
(ibridi) genera una generazione F2 costituita dal 25% di omozigoti recessivi, dal 50% di
eterozigoti e dal 25% di omozigoti dominanti. Il rapporto fenotipico dominanti/recessivi è 3:1
F1
Ibrido x Ibrido
F2
75% dominanti
25% recessivi
Le leggi di Mendel presentano delle eccezioni (dominanza incompleta e codominanza).
La pianta della bella di notte (Mirabilis jalapa) può avere fiori rossi, bianchi o rosa. Le piante
con fiori rossi hanno due copie dell’allele R per il colore rosso e sono omozigoti dominanti RR.
Le piante con fiori bianchi hanno due copie dell’allele r per il colore bianco e sono omozigoti
recessive rr. Le piante con una copia di ciascun allele, cioè gli eterozigoti Rr, hanno fenotipo
rosa, intermedio tra i colori prodotti dai due alleli. Questo è un esempio di dominanza
incompleta.
32
Reincrocio o Test-cross
Pratica sperimentale usata per determinare il genotipo incognito di una popolazione
fenotipicamente dominante che potrebbe essere composta sia da omozigoti dominanti che da
eterozigoti. La popolazione dominante viene incrociata con individui omozigoti recessivi
(fenotipo recessivo). La percentuale di recessivi che si ottiene dal reincrocio è 0% (se la
popolazione da testare è tutta omozigote dominante), 50% (se la popolazione da testare è
tutta eterozigote).
Segregazione ed assortimento indipendente (legge dell’indipendenza)
Mendel studiò successivamente la trasmissione di due caratteri per volta eseguendo un incrocio
tra piante di linee pure che differivano contemporaneamente per due caratteri (seme giallo e
liscio x seme verde e rugoso). La generazione F1 risultò costituita esclusivamente da individui
diibridi sia per la forma che per il colore del seme, con semi gialli e lisci. Effettuata
l'autofecondazione di questi ibridi Mendel ottenne una generazione F 2 in cui erano presenti
quattro tipi di semi nel rapporto di 9:3:3:1 e precisamente: semi gialli e lisci (9), semi verdi e
lisci (3), semi gialli e rugosi (3), semi verdi e rugosi (1).
La comparsa di combinazioni fenotipiche nuove (ricombinanti verdi-lisci e gialli-rugosi), con
caratteri scambiati, permise a Mendel di affermare che le coppie di caratteri sono indipendenti
e la trasmissione di uno o l'altro degli alleli di un carattere non influenza la probabilità di
trasmettere uno o l'altro degli alleli per l'altro carattere.
I risultati di questi incroci vengono sintetizzati nella legge dell’indipendenza dei caratteri o
dell’assortimento indipendente: L’incrocio tra due linee pure che differiscono per due
caratteri produce una generazione F1 di ibridi per entrambi i caratteri che, incrociati danno una
generazione F2 in cui sono presenti tutte le combinazioni fenotipiche nella proporzione 9 D/D :
3 D/R : 3 R/D : 1 R/R (D = fenotipo dominante; R = fenotipo recessivo).
Trattandosi di caratteri indipendenti, il carattere colore del seme è portato da una coppia di
omologhi, mentre il carattere forma del seme è portato da un’altra coppia di omologhi. Durante
la gametogenesi Il cromosoma che contiene l’allele G ha il 50% di probabilità di segregare in
un gamete con il cromosoma che contiene l’allele L e il 50% di probabilità di segregare con il
cromosoma che contiene l’allele l. Ovviamente questo vale anche per l’altro omologo, che
contiene l’allele g. In questo modo, l’assortimento indipendente degli omologhi genera 4 tipi di
gameti: GL, Gl, gL, gl, ciascuno con una probabilità del 25% (1/4).
Se inseriamo i 4 tipi di gameti nel quadrato di Punnett, possiamo ottenere i genotipi possibili.
- Eredità legata al sesso
Nell’uomo molti caratteri sono controllati da geni localizzati sui cromosomi sessuali (caratteri
legati al sesso). Il cromosoma Y è approssimativamente lungo un terzo del cromosoma X
33
quindi la maggior parte dei geni del cromosoma X non hanno il corrispondente nel cromosoma
Y. Tra i caratteri X-linked alcuni responsabili di importanti malattie ereditarie (daltonismo ed
emofilia). Il daltonismo consiste nell’incapacità di distinguere alcuni colori (tipicamente rosso
e verde). L’ emofilia determina un forte ritardo nella coagulazione del sangue che provoca una
relativa facilità ad emorragie, spesso inarrestabili. Entrambi i difetti genetici sono portati da
alleli recessivi che si trovano solo sul cromosoma X. Nelle femmine l'effetto recessivo di un
allele X-linked portatore di difetti genetici è mascherato dall'allele dominante sano situato
sull'altro cromosoma X. Nei maschi, invece, gli alleli X-linked recessivi manifestano i loro effetti
perché non è presente un altro X con geni dominanti che li mascherino e sul cromosoma Y
manca l’allele.
Queste malattie si manifestano quasi esclusivamente nei maschi, mentre le femmine sono
portatrici sane (eterozigoti). Una femmina può presentare tre genotipi:
Genotipo
Fenotipo
XEXE
sana
omozigote
dominante
XEXe
portatrice sana
eterozigote
XeXe
malata
omozigote recessivo
Un maschio che non possiede l’allele sul cromosoma Y può presentare solo due genotipi:
Genotipo
Fenotipo
XEY0
sano
omozigote
dominante
XeY0
malato
eterozigote
Gli incroci che possono avvenire sono:
Femmina sana
XEXE
♂
x
Maschio malato
XeY0
Xe
Y0
XE
XEXe
XEY0
XE
XEXe
XEY0
♀
Se nasce una femmina sarà sicuramente portatrice sana (riceve il cromosoma X e malato dal
padre). Se nasce un maschio sarà sicuramente sano (i maschi non possono mai ricevere un
difetto X-linked dal padre poiché dal padre ricevono sempre il cromosoma Y che non presenta il
difetto genetico). Dunque i maschi malati trasmettono la malattia alle figlie portatrici sane.
Femmina portatrice
XEXe
♂
x
Maschio sano
XEY0
XE
Y0
XE
XEXE
XEY0
Xe
XEXe
XeY0
♀
Se nasce una femmina avrà il 50% di probabilità di essere sana (se riceve dalla madre l’allele
dominante) ed il 50% di probabilità di essere portatrice sana (se riceve dalla madre l’allele
recessivo).
Se nasce un maschio avrà il 50% di probabilità di essere sano (se riceve dalla madre l’allele
dominante) ed il 50% di probabilità di essere malato (se riceve dalla madre l’allele recessivo). I
maschi ereditano dunque la malattia dalla madre portatrice sana (trasmissione per via
materna, ed hanno solo 1 probabilità su 2 di nascere malati).
Affinché nascano femmine malate è necessario un improbabile incrocio tra una femmina
portatrice sana ed un maschio malato.
34
Femmina portatrice
XEXe
x
Maschio malato
XeY0
Xe
Y0
XE
XEXe
XEY0
Xe
XeXe
XeY0
♂
♀
Se nasce una femmina avrà il 50% di probabilità di essere portatrice sana (se riceve dalla
madre l’allele dominante) ed il 50% di probabilità di essere malata (se riceve dalla madre
l’allele recessivo). Se nasce un maschio avrà il 50% di probabilità di essere sano (se riceve
dalla madre l’allele dominante) ed il 50% di probabilità di essere malato (se riceve dalla madre
l’allele recessivo).
- Pleiotropia
Fenomeno per il quale un unico gene controlla più di un carattere all’interno del medesimo
individuo. Nella maggior parte dei casi il gene pleiotropico codifica per una proteina enzimatica
che sta a monte di diverse vie metaboliche o per una proteina strutturale che viene utilizzata
nella costruzione di tessuti ed organi diversi.
- Eredità poligenica: caratteri a variabilità continua
La maggior parte dei caratteri che manifestano una variabilità continua (caratteri
quantitativi) come peso e altezza sono controllati da numerosi geni (eredità poligenica).
Sono presenti solo due tipi di alleli (fattori): fattori positivi (+) che spingono la
manifestazione del carattere verso un estremo del suo intervallo di variabilità e fattori
negativi (-) che spingono la manifestazione del carattere verso l’altro estremo dell’intervallo di
variabilità. La manifestazione del carattere dipenderà dunque dalla particolare combinazione di
fattori positivi e negativi. I caratteri a variabilità continua presentano una particolare
distribuzione di frequenza (curva a campana o gaussiana). Se poniamo in ascissa le altezze
ed in ordinata la frequenza con cui gli individui di una determinata altezza compaiono nella
popolazione, troveremo:
gli individui che presentano i valori estremi del carattere (con tutti i fattori positivi o tutti i
fattori negativi) sono molto rari, mentre avvicinandoci ai valori intermedi gli individui
diventano sempre più frequenti. Un esempio di eredità poligenica è rappresentato dal colore
della cariosside del frumento, controllato da due soli geni. Il colore dipende dalla quantità di
pigmento presente e va dal rosso cupo al bianco attraverso 5 tonalità complessive. Se
incrociamo due individui che stanno alle due estremità opposte dell’intervallo di variabilità, una
pianta a semi rosso cupo (tutti i 4 fattori positivi: genotipo ++ ++) con una pianta a semi
bianchi (tutti i 4 fattori negativi -- --) otterremo una pianta che riceverà ovviamente un
gamete ++ da un genitore ed un gamete –- dall’altro. In F1 avremo pertanto un individuo dal
colore intermedio, con due fattori positivi e due negativi (++ --). Incrociando la progenie F1 e
ipotizzando che i due geni possano ricombinarsi liberamente (assortimento indipendente) essi
35
produrranno 4 tipi di gameti (++, +-, -+, --) che, posti ai due lati di un quadrato di Punnett,
daranno 16 individui.
1 con quattro fattori positivi ++++
4 con tre fattori positivi ed uno negativo +++6 con due fattori positivi e due negativi ++-4 con un fattori positivo e tre negativi +--1 con quattro fattori negativi ----
Se riportiamo le 5 classi fenotipiche in un istogramma che evidenzi la distribuzione di
frequenza, avremo
Come si può osservare l’aspetto è gia quello della distribuzione gaussiana.
Un carattere controllato da tre geni (6 fattori) presenta in F 2 7 classi di frequenza. Per costruire
la distribuzione di frequenza dovremmo incrociare un individuo con tutti e 6 i fattori positivi
(++ ++ ++) con un individuo con tutti e 6 i fattori negativi (-- -- --). Ovviamente otterremo in
F1 individui intermedi con tre fattori positivi e tre negativi (+- +- +-). Se incrociamo tra loro gli
individui intermedi, nell’ipotesi che i fattori siano indipendenti e che quindi durante la
gametogenesi vi sia una completa ricombinazione tra i fattori positivi e negativi, avremo 2 3 = 8
tipi di gameti (+++, ++-, +-+, +--, ---, --+, -+-, -++). Posizionando i gameti ai due lati di un
quadrato di Punnett otterremo 8 x 8 = 64 combinazioni genotipiche.
- Mutazioni
Alterazioni casuali, permanenti ed ereditarie dell’informazione genetica a livello del DNA, ad
opera di agenti chimici e fisici ambientali (agenti mutageni), a causa del normale
metabolismo che produce intermedi reattivi (radicali liberi) in grado di ossidare le molecole
36
organiche, oppure a causa di errori nei processi di divisione cellulare durante la duplicazione
del DNA o durante la meiosi.
La maggior parte delle mutazioni sono sfavorevoli, in alcuni casi possono essere addirittura
letali. Le conseguenze di una mutazione dannosa sono diverse a seconda che si manifestino a
carico delle cellule somatiche o riproduttive. In prospettiva biologica le mutazioni sono però un
evento fondamentale per l’evoluzione, essendo le principali responsabili della comparsa di
nuovi geni e nuovi alleli che creano nuova variabilità genetica sulla quale può agire la selezione
naturale.
Gli agenti mutageni fisici sono calore elevato e radiazioni elettromagnetiche ad alta energia
(raggi UV, raggi X e raggi gamma); essi degradano il DNA, spezzandone i legami chimici.
Gli agenti mutageni chimici alterano la struttura molecolare delle basi causando appaiamenti
errati o si inseriscono fra le basi azotate, legandosi ad esse e causando inserzioni e delezioni
durante la replicazione del DNA.
Le mutazioni genetiche si classificano in geniche, cromosomiche e genomiche.
- Mutazioni geniche
Alterano un singolo gene, modificando una singola base azotata (mutazioni puntiformi).
Le mutazioni per sostituzione di una base azotata modificano una tripletta di basi che
codifica per un amminoacido. Gli effetti possono essere diversi; la nuova tripletta potrà
codificare per medesimo amminoacido, nessun amminoacido o un amminoacido diverso.
Nel caso la nuova tripletta codifichi per il medesimo amminoacido, la sostituzione non avrà
alcun effetto sulla struttura primaria della proteina (mutazione silente). Nel caso non codifichi
per nessun amminoacido (tripletta STOP), la proteina mutata si spezza in due frammenti e
perde la sua funzionalità (mutazione non-senso). Nel caso codifichi per un amminoacido
diverso, la mutazione di senso (missense) ha effetti sulla funzionalità proteica che
dipendono dal punto in cui è avvenuta. Se colpisce un tratto funzionale della proteina, la cui
alterazione compromette la sua attività, oppure tratti non funzionali in cui la sostituzione di un
solo amminoacido non produce effetti. Le sostituzioni amminoacidiche in tratti funzionali
possono alterare in modo più o meno grave la funzionalità proteica. Ne è un esempio l’anemia
falciforme, malattia genetica recessiva in cui i globuli rossi assumono una caratteristica forma
a falce, che è dovuta alla sostituzione di un unico amminoacido (valina al posto dell'acido
glutammico) nelle catene beta dell’emoglobina. Le sostituzioni in tratti non funzionali (neutre)
non portano ne’ vantaggi ne’ svantaggi poiché lasciano inalterata la funzionalità proteica.
Le mutazioni per inserzione provocano l’inserimento di un nuovo nucleotide. Le mutazioni
geniche per delezione provocano l’eliminazione di un nucleotide. Queste mutazioni
provocano oltre ad una modificazione del significato della tripletta di nucleotidi mutata, anche
una sfasatura nella lettura di tutte le triplette successive (mutazioni con slittamento del modulo
di lettura o frame-shift). Il cambiamento di tutti gli amminoacidi successivi al punto in cui è
avvenuta la mutazione porta per lo più alla perdita della funzionalità proteica.
- Mutazioni cromosomiche (aberrazioni)
Sono alterazioni della struttura dei cromosomi che interessano più geni e si producono in
conseguenza di errori durante il crossing-over per rotture ed errate ricomposizioni dei
cromosomi. Si possono così verificare perdite, duplicazioni, inversioni o trasferimenti di pezzi
più o meno estesi di cromosoma. I geni si vengono a trovare in una posizione diversa da quella
originale e, dato che la regolazione dell’attività di un gene dipende anche dalla sua
localizzazione, le mutazioni cromosomiche hanno generalmente effetti drammatici.
37
L’inversione origina da due rotture sul cromosoma, rotazione di 180° del tratto compreso tra
le rotture e sua reintegrazione nel cromosoma stesso. Si origina in genere durante
l’appaiamento degli omologhi.
La traslocazione (trasposizione) consiste nel trasferimento di un segmento di cromosoma in
una posizione errata, con un cambiamento nella localizzazione del segmento cromosomico. Può
essere intracromosomica o intercromosomica. La prima implica un cambiamento di posizione di
un tratto cromosomico entro lo stesso cromosoma. La seconda lo spostamento di un segmento
cromosomico ad un altro non omologo. Una traslocazione unilaterale tra cromosomi omologhi
produce una delezione su di un cromosoma ed una duplicazione sull’altro cromosoma. La
delezione porta alla perdita di un segmento di cromosoma; l’effetto dipende dalla grandezza
della porzione mancante e dal tipo di informazione contenuta. La duplicazione porta alla
presenza di due segmenti genici identici all’interno del medesimo cromosoma.
- Mutazioni genomiche
Sono alterazioni del numero dei cromosomi, come conseguenza della perdita o dell’aggiunta di
interi cromosomi (soprannumerari). Sono causate dalla mancata disgiunzione degli omologhi
durante la meiosi, per cui i cromosomi non si distribuiscono equamente nei gameti. La
variazione del numero corretto di cromosomi si definisce aneuploidia. La monosomia è la
presenza di un singolo cromosoma al posto di una coppia all’interno di un corredo diploide.
La trisomia è la presenza di tre omologhi al posto di una coppia all’interno di un corredo
diploide. Se la mancata disgiunzione riguarda l’intero corredo cromosomico si possono formare
cellule poliploidi (triploidi 3n, tetraploidi 4n) con comparsa di intere serie di cromosomi
soprannumerari. La poliploidia è in genere una condizione letale per gli animali, mentre nei
vegetali è diffusa (frumento, cotone, tabacco) e può produrre gigantismo e maggior
produttività.
- Aneuploidie autosomiche
Alcune anomalie degli autosomi comportano effetti gravi ma compatibili con la vita come la
trisomia 21 (sindrome di Down), la trisomia 13 (sindrome di Patau) e la trisomia 18 (sindrome
di Edwards). Le altre trisomie non sono compatibili con la vita e sono causa di aborto
spontaneo.
La sindrome di Down (trisomia 21) è causata dalla mancata disgiunzione dei cromosomi
della coppia 21. Le persone affette hanno nelle loro cellule 47 cromosomi. L’incidenza della
trisomia aumenta drasticamente con l’età della madre. Gli affetti presentano palpebre
anormali, naso schiacciato, lingua larga, mani corte e robuste, statura bassa, ritardo mentale,
probabili malformazioni cardiache, attesa di vita minore del normale.
La sindrome di Patau (trisomia 13) è caratterizzata da gravi ritardi mentali. Come la
sindrome di Down è legata all’età della madre. Gli affetti manifestano cranio ed occhi piccoli,
difetti cardiaci, dita soprannumerarie e sordità.
La sindrome di Edwards (trisomia 18) è caratterizzata da ritardo mentale e fisico. Gli affetti
manifestano cranio allungato, orecchie con basso punto di attacco, cavità orale piccola e difetti
cardiaci.
- Aneuploidie dei cromosomi sessuali
Le anomalie più frequenti dei cromosomi sessuali sono le trisomie XXX, XXY, XYY e la
monosomia X0. Essendo la Y mascolinizzante, la sua presenza indica il sesso maschile.
38
La trisomia XXY (sindrome di Klinefelter) è causata dalla non disgiunzione delle X nella
produzione dell’ovulo. Gli individui affetti sono maschi con testicoli non sviluppati, mancata
produzione di spermatozoi, assenza di pelosità e barba, tono della voce che resta acuto. In
genere si accompagnano una statura alta e funzione intellettuale leggermente deficitaria.
La monosomia X0 (sindrome di Turner) produce femmine di bassa statura, collo tozzo e
corto, organi sessuali infantili, assenza spesso di ovaie. Le capacità intellettive risultano quasi
normali.
Gli affetti della trisomia XXX sono femmine quasi normali sul piano fisico e riproduttivo, ma
manifestano spesso consistenti ritardi mentali.
Gli affetti della trisomia XYY sono maschi che, a parte la tendenza a superare mediamente i
180 cm di statura, non manifestano sintomi fisici particolari. E’ causata da una mancata
disgiunzione dei due cromatidi che formano il cromosoma Y durante la seconda divisione
meiotica.
- Malattie genetiche
Le malattie monogeniche sono causate dall’alterazione di un unico gene e seguono le leggi
della ereditarietà mendeliana. La maggior parte di esse è portata da alleli recessivi. Si
classificano in:
- Malattie autosomiche recessive
- Malattie autosomiche dominanti
- Malattie X-linked (legate al cromosoma X)
- Malattie Y-linked (legate al cromosoma Y)
Le malattie geniche autosomiche recessive sono causate da alleli recessivi presenti negli
autosomi. La malattia si manifesta solo se sono presenti entrambi gli alleli (omozigoti
recessivi). Gli eterozigoti sono portatori sani. Ognuno dei genitori porta un allele normale e uno
difettoso e ha una probabilità del 50% di produrre un gamete con un allele difettoso. Ne
consegue che la progenie avrà genotipi nella proporzione 1:2:1 e fenotipi nella frequenza di
3:1. Vale a dire che il 75% della progenie apparirà normale e il 25% evidenzierà un fenotipo
recessivo (malato). Esempi di malattie autosomiche recessive sono la fenilchetonuria, fibrosi
cistica (mucoviscidosi), anemia falciforme (drepanocitosi), beta talassemia, albinismo.
La fibrosi cistica (CF) è caratterizzata dall’infezione cronica delle vie aeree conseguente a
mutazioni in un gene localizzato sul cromosoma 7.
La fenilchetonuria (PKU) è caratterizzata da un’aumentata concentrazione nel sangue e
nell’urina dell’amminoacido fenilalanina e dei suoi derivati e da grave ritardo mentale.
La talassemia major (Morbo di Cooley) è una grave forma di anemia ereditaria diffusa in
tutto il bacino del Mediterraneo.
Le malattie geniche autosomiche dominanti sono causate da alleli dominanti presenti negli
autosomi. Sono affetti anche gli eterozigoti che portano l’allele malato solo in uno dei due
cromosomi ereditati dai genitori. Sono estremamente improbabili individui malati omozigoti
dominanti, poiché dovrebbero essere figli di genitori entrambi malati. Ogni affetto ha il 50% di
probabilità di avere figli malati ad ogni gravidanza. Sono esempi l’Acondroplasia, una forma
di nanismo, corèa di Huntington (ballo di SanVito) che causa una progressiva degenerazione
del sistema nervoso e la distrofia muscolare miotonica.
Malattie geniche legate al cromosoma X (X-linked)
Si tratta di malattie portate da alleli recessivi o dominanti localizzati solo sul cromosoma X. Il
cromosoma Y più piccolo del cromosoma X non porta l’allele corrispondente; di conseguenza il
rischio della malattia è diverso nei due sessi. Dato che il maschio presenta un solo cromosoma
X la presenza di un gene mutante dà luogo inevitabilmente alla manifestazione morbosa. Le
malattie ereditarie legate al sesso non possono essere trasmesse da maschio a maschio,
mentre il padre le trasmette a tutte le figlie.
Le principali forme ereditarie legate al sesso di tipo recessivo sono l’emofilia A, la distrofia
muscolare tipo Duchenne, il deficit di glucosio-6-fosfato-deidrogenasi, il daltonismo, la
sindrome dell’X-fragile.
Le malattie geniche Y-linked sono molto rare. Ne sono affetti solo i maschi che trasmettono
la malattia solo ai figli maschi.
Evoluzionismo
L'idea di una natura dinamica contrapposta a una natura statica è una visione molto antica.
L'idea che la Terra, le montagne, i mari, le specie col tempo si trasformino, per affermarsi ha
dovuto sconfiggere un paradigma fissista molto radicato, secondo il quale l'universo è stato
39
fatto in modo definitivo e statico ed ogni cambiamento è solo un evento eccezionale e
catastrofico all’interno di un mondo stabile. L’affermarsi di concezioni evoluzionistiche è
strettamente legato alla “scoperta del tempo profondo”. Per gli uomini del Seicento la terra
non aveva più di 6000 anni. La difficoltà di concepire la storia della Terra secondo intervalli di
tempo più estesi rendeva impossibile una spiegazione di tipo evolutivo dei fenomeni naturali.
L’evoluzionismo interpreta infatti i fenomeni del presente come il risultato del succedersi di
piccole, lentissime e impercettibili variazioni che in tempi lunghissimi possono generare enormi
cambiamenti. Il 1785 fu decisivo nella storia del pensiero evoluzionistico grazie all’opera del
geologo scozzese James Hutton fondatore della moderna geologia. La storia della Terra può
essere spiegata sulla base del ripetersi di semplici fenomeni geologi che si avvicendano
uniformemente e ciclicamente in tutti i momenti della storia geologica e in tutte le parti del
globo (uniformismo). Questi processi richiedono tempi lunghissimi. In questo senso va
interpretata l’affermazione “il presente è la chiave per spiegare il passato”. Charles Lyell
(1797-1875) raccolse ed approfondì la lezione di Hutton e introdusse il principio
dell’attualismo (gli eventi del passato si spiegano con forze ancora operanti e osservabili). Le
modificazioni avvenute sulla Terra nelle ere geologiche passate dovevano essersi verificate con
le stesse modalità e con la stessa intensità degli eventi geologici osservabili nel presente. I
cambiamenti sono lenti e costanti (gradualismo) e se gli effetti sono stati assai pronunciati
ciò è dovuto all’accumularsi, in tempi molto lunghi, di tanti piccoli eventi. Il fattore tempo è di
fondamentale importanza poiché anche piccole variazioni, sommate per intervalli di tempo
sufficientemente lunghi, possono produrre effetti enormi.
Evoluzione biologica
Verso la fine del ‘700 la maggior parte degli scienziati accettava ormai l’idea che la complessa
storia della vita sulla terra si fosse necessariamente svolta nell’arco di tempi lunghissimi e che i
fossili rappresentassero le vestigia di specie estinte. L’idea di un’evoluzione biologica dei
viventi cominciava ad avere un terreno fertile su cui crescere. Le due teorie evolutive avanzate
furono il lamarckismo e il darwinismo. L’idea dominante fino alla prima metà dell’Ottocento
fu quella della fissità delle specie, cioè della loro immutabilità. Il fissismo fu codificato dal
naturalista svedese Carlo Linneo che nel 1751, nella sua opera Philosophia botanica, affermava
che le forme viventi furono create per progetto divino (“Species tot sunt diversae quot diversas
formas ab initio creavit infinitum Ens”).
La prima teoria articolata e coerente dell’evoluzione biologica si deve a J. B. Lamarck. Le sue
ipotesi furono pubblicate nell’opera “Philosophie zoologique” (1809). Lamarck ritenne che ci
potesse essere una derivazione degli organismi più complessi e più recenti da quelli più
semplici e più antichi che dipendeva per Lamarck da due fattori:
- i nuovi caratteri compaiono negli esseri viventi in risposta alla necessità e nuovi bisogni
indotti da un ambiente in continua trasformazione. I nuovi caratteri si sviluppano con l'uso,
mentre regrediscono e scompaiono se inutilizzati (principio dell’uso e non uso).
- i nuovi caratteri acquisiti vengono trasmessi per eredità alle generazioni successive
(principio dell’ereditarietà dei caratteri acquisiti).
Il suo esempio più famoso è quello della giraffa moderna che si sarebbe evoluta da antenati a
collo corto, costretti ad allungarlo per raggiungere le foglie più alte dei rami in periodi di scarsa
disponibilità alimentare. Questi antenati trasmisero il collo allungato ai loro discendenti i quali
continuarono ad allungarlo e così via. Alla luce delle conoscenze di genetica sappiamo che un
organismo può effettivamente perfezionare con l'uso un certo organo come, d'altra parte, può
giungere all'atrofia di certe funzioni se non adeguatamente utilizzate, ma ogni modificazione
dell'organismo che non si rifletta in una corrispondente modificazione del corredo cromosomico
non può essere ereditata e dar luogo a nessuna forma di evoluzione. I caratteri acquisiti sono
modificazioni del fenotipo e non del genotipo.
Darwin e la selezione naturale
Charles Darwin e Alfred Russel Wallace formularono contemporaneamente e separatamente
una teoria evolutiva dei viventi per selezione naturale. Nel 1859 Darwin pubblicò, dopo un
lavoro di elaborazione durato circa venti anni "On the origin of species by mean of natural
selection", un'opera fondamentale che rivoluzionò ogni precedente teoria. L'originalità del suo
lavoro sta nell'aver dato un'interpretazione scientificamente soddisfacente dei meccanismi
evolutivi. Fu fondamentale per la formulazione della teoria darwiniana la conoscenza del lavoro
dell'economista inglese Thomas Malthus. "Saggio sul principio di popolazione" (1798) e delle
ipotesi attualiste sostenute dal geologo inglese Charles Lyell, nei suoi "Principi di geologia".
Nel suo "Saggio" Malthus afferma che mentre la popolazione umana si sviluppa secondo una
progressione geometrica, quindi con andamento esponenziale, le risorse crescono seguendo
40
una progressione aritmetica. E' quindi inevitabile che si arrivi ad una situazione in cui le risorse
non sono sufficienti per tutta la popolazione. Darwin intuì che le conclusioni di Malthus,
secondo cui il cibo e le risorse sono un fattore limitante per la crescita delle popolazioni, erano
vere per tutte le specie e non solo per la specie umana. Ad esempio Darwin calcolò che una
coppia di elefanti sarebbe in grado di generare in 750 anni circa 19 milioni di individui. Poiché
invece il numero degli elefanti rimane costante, Darwin ne dedusse che gli elefanti attualmente
presenti devono essere stati "scelti" tra tutti quelli che si sarebbero potuti generare. Il processo
attraverso cui i viventi sono scelti per perpetuare la loro specie è la selezione naturale. Dopo
un lavoro di elaborazione durato più di vent’anni, Darwin sintetizzò le sue osservazioni in una
teoria organica dell'evoluzione, basata sulla selezione naturale, i cui punti essenziali possono
essere così riassunti:
1) tutti gli organismi producono una progenie di gran lunga superiore alle capacità che
l'ambiente ha di sostenerla. Per questo motivo tra tutti gli individui che nascono solo
alcuni sono in grado di raggiungere la maturità ed hanno la possibilità di riprodursi;
2) in ogni generazione sono presenti individui casualmente diversi gli uni dagli altri, con
una grande variabilità di caratteri;
3) alcuni caratteri possono rendere l'individuo che li porta più adatto a sopravvivere in quel
determinato ambiente. Altri possono risultare indifferenti o addirittura dannosi;
4) tra gli individui di una generazione solo quelli che presentano casualmente i caratteri
che li rendono più adatti sopravvivono ed hanno così la possibilità di prolificare. Ciò
consente loro di trasmettere i loro caratteri alla discendenza, mentre i caratteri degli
individui inadatti si estinguono con essi.
L'ambiente si comporta come un filtro in grado di selezionare, di generazione in generazione,
solo gli organismi viventi più adatti alla sopravvivenza.
Darwin capovolge la teoria di Lamarck. Non è l'ambiente che induce i cambiamenti
evolutivi negli organismi, ma i cambiamenti sono già presenti casualmente in ogni
generazione sotto forma di variabilità individuale. L'ambiente attua solo una
selezione su tale variabilità.
Per Darwin l’ambiente gioca un ruolo puramente di filtro, mentre per Lamarck esercita un ruolo
attivo, inducendo la variazione dei caratteri.
Poiché non tutti gli individui possono giungere a maturità, essendo le risorse naturali
insufficienti, si instaurerà una competizione tra gli individui per la conquista delle risorse, la
lotta per la sopravvivenza, mutuata dalla lettura di Malthus. A questo punto saranno
"naturalmente selezionati" e sopravviveranno, perpetuando così i loro caratteri solo gli
individui più adatti a quel determinato ambiente. Per Darwin la selezione naturale era analoga
al processo di selezione praticato per il bestiame in cui gli uomini scelgono gli esemplari di
piante o di animali da far riprodurre in base alle caratteristiche che sembrano loro più utili; nel
caso della selezione naturale l'ambiente si sostituisce all'uomo.
La selezione naturale proposta da Darwin è un meccanismo assolutamente casuale e
adirezionale, che esclude la necessità di un progetto in natura.
- Prove dell’evoluzione
paleontologia
La datazione assoluta dei fossili indica che quelli più antichi sono meno complessi dei fossili più
recenti.
anatomia comparata
Osservando organi omologhi di classi diverse di esseri viventi, è possibile, pur nell’apparente
diversità, rilevare uno stesso progetto sviluppato con variazioni sul tema, come se ciascuna
classe si fosse sviluppata da un comune progenitore. Le strutture omologhe sono parti di
organismi di gruppi sistematici diversi composte da elementi simili per forma e numero ed
hanno rapporti anatomici simili.
embriologia comparata
Confrontando embrioni differenti, ad esempio quelli delle classi dei vertebrati prima citati, nei
primissimi stadi di sviluppo, si osserva una somiglianza impressionante. In tutti sono ad
esempio presenti le fessure branchiali e la coda, anche in quelli che nella forma adulta non
presentano tali caratteristiche. Negli embrioni degli uccelli e delle balene, animali che da adulti
sono privi di denti, compaiono abbozzi dentali che poi regrediscono.
citologia
Non può essere casuale che tutti i viventi siano costruiti da mattoni praticamente identici, le
cellule che presentano la stessa organizzazione strutturale e funzionale.
biochimica
41
Anche le sostanze chimiche complesse, necessarie alla sopravvivenza degli esseri viventi, sono
praticamente le stesse per tutte le specie. Il codice genetico nel quale sono immagazzinate
tutte le informazioni necessarie alla vita è universale, il significato delle triplette è lo stesso per
tutti gli esseri viventi.
42