…i nostri occhi ricevono vaghe
immagini delle cose, rovesciate e
distorte, ed il risultato è che noi
vediamo il mondo che ci circonda
pieno di oggetti solidi e ben distinti…e
questo è poco meno che un miracolo
Gregory, Eye and Brain, 1966
Fenomeni di costanza
• Il sistema visivo riesce a percepire un oggetto
come identico, in condizioni diverse e capaci di
far variare ampiamente l’immagine retinica
• Quando ci muoviamo cambiano le condizioni di
illuminazione, la dimensione, la forma e la
luminosità delle immagine proiettate sulla retina
dagli oggetti, eppure la nostra percezione degli
oggetti non cambia
Le sensazioni e percezioni
• Le percezioni non sono semplici registrazioni
delle proprietà fisiche degli stimoli
• L’esperienza sensoriale è una costruzione
cerebrale ma non arbitraria
• È realizzata secondo regole e limiti imposti dalle
proprietà del SN!
• È un’organizzazione accurata delle proprietà degli
oggetti che ci consente di far uso degli stessi
Le sensazioni e percezioni
• Le percezioni non sono semplici registrazioni
delle proprietà fisiche degli stimoli
• L’esperienza sensoriale è una costruzione
cerebrale ma non arbitraria
• È realizzata secondo regole e limiti imposti dalle
proprietà del SN!
• È un’organizzazione accurata delle proprietà degli
oggetti che ci consente di far uso degli stessi
I sistemi sensoriali hanno un
piano organizzativo comune
• Estraggono le stesse informazioni di base
• Il contatto iniziale è con strutture nervose specializzate: i
recettori
• I recettori trasformano l’energia dello stimolo in energia
elettrochimica (trasduzione) ovvero in segnali nervosi =
creano un linguaggio comune
• Le info sullo stimolo sono rappresentate da una serie di
potenziali d’azione (codificazione nervosa)
Organizzazione seriale vs parallela
Recettori
ordine
Seriale o gerarchica
neuroni I ordine
neuroni di II
neuroni di ordine superiore
Parallela
Caratteristiche diverso di uno stimolo complesso
vengono elaborate a livello di vie diverse
Organizzazione topografica
• Relazioni di vicinanza esistenti in periferia
sono conservate a livello del SNC
Due punti vicini nello spazio
Eccitano due recettori vicini
Proiettano a neuroni del SNC adiacenti
Stazione di relè
• Per tutti i sistemi sensoriali, ad eccezione
dell’olfatto, il talamo funge da stazione di
ritrasmissione: qui fanno sinapsi le fibre dei
neuroni di secondo ordine e a loro volta i
neuroni talamici di ogni sistema sensoriale
proiettano ad aree sensoriali primarie
specifiche
La natura della luce
• La luce è costituita da particelle chiamate fotoni,
ciascuno dei quali è un pacchetto di onde
elettromagnetiche
• Che un raggio di energia elettromagnetica sia luce
e non una radiazione X o un’onda radio dipende
dalla lunghezza d’onda (distanza tra due picchi
successivi dell’onda)
• Noi vediamo dai 400 ai 700 nanometri
Spettro del visibile
• Gran parte della luce che colpisce gli occhi
è una miscela ben equilibrata di energia a
lunghezza d’onda differenti ed è chiamata
luce bianca (es. luce del sole)
• Se filtriamo in modo da eliminare tutte le
lunghezze d’onda tranne una ristretta banda
otteniamo una luce monocromatica
Attraversare
(es.acqua, vetro) ed
essere deviata o rifratta
Assorbita
Riflessa
Pigmenti: sostanze che riflettono
selettivamente alcune lunghezze d’onda
Assorbe le lunghezze
d’onda brevi e medie,
riflette le lunghe
Assorbe le lunghezze
d’onda brevi e lunghe,
riflette le medie
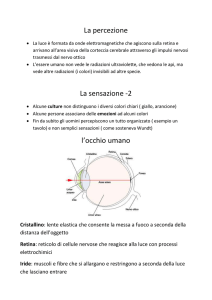
L’occhio
L’occhio è uno strumento
ottico
deputato a
focalizzare sulla retina le
immagini del mondo
•I sistemi diottrici più
potenti sono cornea (2/3
del potere refrattivo) ed
il cristallino
La retina
• La retina ha una struttura laminare
• La disposizione della retina è peculiare: I
fotorecettori sono distali (esterni) e le cellule
gangliari occupano gli strati prossimali (interni)
• I corpi cellulari sono contenuti in tre strati nucleari
– Stato nucleare esterno (contiene I fotorecettori)
– Strato nucleare interno (cellule bipolari)
– Strato delle cellule gangliari
La retina
• I processi sono contenuti in due strati plessiformi che
contengono la maggior parte dei contatti sinaptici
– Strato plessiforme esterno: contiene i processi di
fotorecettori, cellule bipolari ed orizzontali
– Strato plessiforme interno: contiene i processi delle cellule
bipolari, amacrine e gangliari
La retina
• I fotorecettori sono neuroni
• La retina è in stretto contatto con l’epitelio
pigmentato che assolve a tre funzioni:
– Assorbe la luce non trattenuta dalla retina
– Risintetizza i pigmenti visivi fotosensibili
– Fagocita le estremità del segmento esterno dei
fotorecettori
Strato delle
cellule
grangliari
Strato plessiforme
interno
Strato nucleare
interno
Strato plessiforme
esterno
Strato nucleare
esterno
Due tipi di problemi
• Come fanno i coni ed i bastoncelli a trasdurre la luce in
segnali elettrici e chimici?
• In che modo le cellule dei due strati successivi -cellule
bipolari, orizzontali, amacrine- interpretano questa
informazione?
Output dell’occhio
• BUIO: le cellule non sono silenti al buio (1-2/20
impulsi per secondo) CORRENTE AL BUIO
• RISPOSTA ALLA LUCE: consiste in un aumento o
una diminuzione della frequenza di scarica
–Risposta ON: con l’accensione dello stimolo luminoso
aumenta la frequenza di scarica
–-risposta OFF: inibizione durante l’illuminazione e scarica
al suo spegnimento
elettrodi
illumino
1° cellula
2° cellula
Campo recettivo:
zona di retina o porzione di campo visivo la cui
stimolazione produce una risposta
Forma e dimensioni
• I campi recettivi delle cellule gangliari e
bipolari retiniche sono circolari
• In fovea la grandezza del centro è di pochi
minuti di arco, in periferia va dai 3 ai 5 gradi
Campo recettivo:
Ripartizione
• I campi recettivi sono organizzati con un
centro ed una periferia antagonista
(antagonismo degli effetti, sommazione
spaziale)
–Cellule centro on con periferia inibitoria
–Cellule centro off con periferia eccitatoria
Vantaggi della ripartizione del campo
recettivo in due porzioni antagoniste
• Specializzazione per vedere i contrasti: più
informativi circa la presenza di un oggetto rispetto
alla quantità assoluta di luce riflessa dall’oggetto
stesso(dipende dalla sorgente luminosa)
• La visione dei contrasti inizia in retina per
minimizzare la distorsione nel corso della
trasmissione
VANTAGGIO DELLA PERCEZIONE DEI
CONTRASTI
ESTERNO INTERNO
CARTA BIANCA
120
6.0
CARATTERE NERO
12
0.6
VANTAGGIO DELLA PERCEZIONE DEI
CONTRASTI
• L’aspetto bianco o nero di un oggetto non è
in funzione della quantità di luce da essa
riflessa ma dalla quantità di luce riflessa
dall’oggetto in confronto a quella riflessa
dagli oggetti circostanti
• Così: ciò che è nero rimane nero
indipendentemente dalla intensità della
sorgente luminosa
Vantaggi nella presenza di due vie
parallele
• È più economico
– Un tipo di cellula segnala l’aumento di intensità
– L’altra segnala la diminuzione dell’intensità
Due tipi di problemi
• Come fanno i coni ed i bastoncelli a trasdurre la luce in
segnali elettrici e chimici?
• In che modo le cellule dei due strati successivi -cellule
bipolari, orizzontali, amacrine- interpretano questa
informazione?
elettrodi
illumino
1° cellula
2° cellula
Fotorecettori: come sono costituiti?
• Segmento esterno (specializzato per la
trasduzione)
• Segmento interno (contiene il nucleo)
• Terminazione sinaptica
I fotorecettori
Il segmento esterno
• Il segmento esterno contiene i pigmenti visivi capaci
di assorbire la luce
• I pigmenti sono legati ad una proteina di membrana
• Per aumentare la superficie di membrana si
sviluppano dei dischi
–Bastoncelli: si separano dalla membrana e si dispongono
uno sull’altro nel citoplasma
–Coni: seguono di pari passo la membrana
Proprietà dei coni e dei bastoncelli
CONI
BASTONCELLI
• Visione fotopica (< quantità di
fotopigmento)
• > risoluzione spaziale (+
presenti in fovea e con minore
convergenza)
• Visione dei colori (tre pigmenti
differenti, sensibili alle diverse
lunghezze d’onda)
• Visione
scotopica
(>
quantità di fotopigmento)
• Minore risoluzione spaziale
(+
presenti
in
periferia/maggiore
convergenza)
• Acromatici
I recettori visivi
• I bastoncelli ed i coni della retina contengono pigmenti
differenti per struttura chimica e di conseguenza nelle capacità
relative di assorbire luce
– Bastoncelli (un solo tipo di pigmento )
– Coni hanno pigmenti con picchi di assorbimento !
420 nm =lunghezze d’onda corte “coni del blu”
530 nm= lunghezze d’onda medie “coni del verde”
560 nm= lunghezze d’onda lunghe “coni del rosso”
Com’è la risposta dei
fotorecettori
• I fotorecettori al buio sono depolarizzati: il
potenziale di membrana è intorno a -40 mV
• La risposta alla luce dei fotorecettori è
l’iperpolarizzazione: il potenziale di
membrana passa da -40 mV a -70 mV
AL BUIO I FOTORECETTORI SONO ATTIVI
=DEPOLARIZZATI
LUCE
I FOTORECETTORI SI SPENGONO
= SI IPERPOLARIZZANO
QUESTO SPIEGA
• Il perché se si registra dal nervo ottico si
trova una attività spontanea (non silente) al
buio
Come avviene la fototrasduzione
• La luce attiva i pigmenti visivi (il retinale cambia
conformazione e di conseguenza cambia
conformazione anche l’opsina -metarodopsina II-)
• La forma attiva della rodopsina attiva centinaia di
molecole di trasducina che stimolano la
produzione dell’enzima che catabolizza il GMPc
Come avviene la fototrasduzione
• La
riduzione
della
concentrazione
citoplasmatica del GMPc chiude i canali
GMPc-dipendenti (segmento esterno)
• Questo diminuisce l’entrata di NA+ mentre
continua il passaggio passivo del K+ fuori
la cellula (segmento interno)
• La cellula si iperpolarizza
Come avviene la fototrasduzione
IPERPOLARIZZAZIONE
TRASDUCINA
LUCE
TRASFORMA
RODOPSINA
GMPc
<Na+
TRASDUCINA
GMPc
<Na+
TRASDUCINA
GMPc
<Na+
TRASDUCINA
GMPc
<Na+
TRASDUCINA
GMPc
<Na+
BUIO=
FOTORECETTORI DEPOLARIZZATI
RILASCIO DI
GLUTAMMATO
INIBISCE
BIPOLARI
ON
ECCITA
OFF
SI IPERPOLARIZZA=
SI DEPOLARIZZA=
CESSA DI INVIARE IMPULSI
INVIA IMPULSI
LUCE=
FOTORECETTORI IPERDEPOLARIZZATI
NON RILASCIANO
GLUTAMMATO
VIENE MENO
INIBIZIONE
BIPOLARI
VIENE MENO
L’ECCITAZIONE
ON
OFF
SI DEPOLARIZZA=
SI IPERPOLARIZZA=
INVIA IMPULSI
CESSA DI INVIARE IMPULSI
Interneuroni
• Cellule orizzontali
– Ricevono input dai recettori
– Come i recettori sono iperpolarizzate alla luce
(connessioni eccitatorie con I recettori)
– Non hanno campi recettivi strutturati in centro e
periferia
– Output diretto sia ai fotorecettori sia alle cellule
bipolari
– Responsabili, almeno in parte, della periferia del campo
recettivo delle cellule bipolari
Interneuroni
• Cellule amacrine
– Ricevono input dalle cellule bipolari
– Probabilmente vi sono più di venti tipi
– Non hanno campi recettivi strutturati in centro e
periferia
– Output diretto alle cellule gangliari con
funzioni diverse
• Coinvolte nella organizzazione centro
periferia
• Ruolo nelle risposte specifiche al movimento
Campo visivo:
quanto vediamo?
60°
Punto di fissazione
Emicampo DX
90°
+
75°
Campo visivo:
il sopra ed il sotto
• Le immagini retiniche sono invertite
rispetto al campo visivo:
–
la metà superiore del campo visivo è
proiettata sulla metà inferiore della retina
– la metà inferiore del campo visivo viene
proiettata sulla metà superiore della retina
Campo visivo:
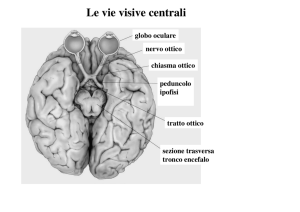
la destra e la sinistra
• Dopo
il
chiasma
ottico
la
rappresentazione del campo visivo è
controlaterale:
– il tratto ottico di sinistra
rappresentazione completa
visivo destro
– il tratto ottico di destra
rappresentazione completa
visivo sinistro
conterrà una
dell’emicampo
conterrà una
dell’emicampo
….e dopo la retina?
Nucleo soprachiasmatico
TALAMO:
NGL
MESENCEFALO
COLLICOLO SUPERIORE
•Corteccia visiva primaria
•Corteccie exstrastriate
•Infero-temporale
•Parientale posteriore
Il corpo genicolato laterale
• contiene una rappresentazione completa
dell’emicampo visivo controlaterale
• la rappresentazione è visuotopica: le relazioni
spaziali sono rispettate
• costituito da sei strati, monoculari, i due
ventrali magnocellulari e i quattro dorsali
parvocellulari
• riceve input dalla formazione reticolare, e dalla
corteccia
…a cosa rispondono le sue
cellule?
• non sostanziali modifiche rispetto alla retina
• i campi recettivi sono concentrici: lo stimolo
migliore è un puntino luminoso su uno sfondo
scuro (centro ON - periferia OFF), o un puntino
scuro su uno sfondo chiaro (centro OFF-periferia
ON)
• vie parallele centro-on, centro off, suddivise
ulteriormente in cellule P ed M
parvocellule
•
•
•
•
Campi recettivi più piccoli
> risoluzione spaziale
Visione dei colori
Rispondono più lentamente
in modo sostenuto
=
minore
risoluzione
temporale
magnocellule
• Campi recettivi più grandi
• < risoluzione spaziale
• Ai contrasti di luminanza
(anche a bassi contrasti)
• Potenziali di azione più
rapidi
con
risposte
transienti
=
ottima
risoluzione temporale
struttura laminare
..e la corteccia striata…a cosa
rispondono le sue cellule?
• I campi recettivi diventano rettilinei (circolari solo
nei blob)
• Selettività e capacità di elaborazione via via
maggiori
–Cellule semplici
–Cellule complesse
•Le cellule diventano binoculari ad eccezione di
quelle dello strato di input da parte dal nucleo
genicolato laterale (strato IV)
CELLULE SEMPLICI
tripartito
_ +_ + -+
on
bipartito
+ _
off
• orientamento
•sommazione spaziale
•antagonismo
++++-
-+
-+
-+
-+
-+
-+
-+
-+
-+-+-+-+-+-+-+-+-
CELLULE complesse
•Stimolo adeguato:linea orientata opportunamente
che colpisca qualunque punto del campo visivo
on
off
-+ - + - +
-+ - + - +
-+ - + - +
-+ - + - +
-+ - + - +
-+ - + - +
-+ - + - +
-+ - + - +
Proprietà dei neuroni semplici e
complessi
SEMPLICI
• Posizione: occupano lo strato
di input (strato IV)
• Numerosità: 1/4 delle cellule
in corteccia
• Campo recettivo: strutturato
in regioni eccitatorie ed
inibitorie
• Selettività: orientamento
•
•
•
•
COMPLESSE
Posizione: occupano gli strati 23 e 5-6
Numerosità : 2/3 delle cellule in
corteccia
Non c’è questa suddivisione:
risposta a linee di orientamento
ottimale in movimento
dovunque esse cadano
Selettività: orientamento,
movimento, 10-20% direzione
movimento
ARCHITETTURA DELLA
CORTECCIA VISIVA PRIMARIA
• struttura laminare
• struttura colonnare
18
19
input
1
2,3
4
a
b
c
5
6
Strati magnocellulari
NGL
Strati parvocellulari
NGL
Strati parvocellulari NGL e
strati interlaminari
I blob
Cellule con campi recettivi circolari, sensibili alle varie lunghezze
d’onda, non sensibili al movimento, nè all’orientamento.
output
1
2,3
4
5
6
Altre aree corticali:
Corteccie extrastriate
Strutture sottocorticali:
NGL, collicolo superiore
CORTECCIA VISIVA PRIMARIA
colonne di dominanza oculare
• se si penetra verticalmente attraverso tutto lo
spessore si trovano cellule con lo stesso grado di
preferenza per uno dei due occhi
• parallelamente alla superficie la dominanza
oculare si alterna continuamente ogni 0.5 mm (in 1
mm abbiamo tutte e due)
colonne di dominanza oculare
1
7
2,3
4
5,6
1
2,3
4
5
6
NGL
CORTECCIA VISIVA PRIMARIA
colonne di orientamento
• se si penetra verticalmente attraverso tutto lo
spessore della corteccia si incontrano cellule che
preferiscono lo stesso orientamento
• parallelamente alla superficie si incontra una
sequenza regolare di cambiamento di
orientamento: ogni 0.05 mm l’orientamento
ottimale si sposta di circa 10 gradi (in 1 mm 180
gradi rappresentati)
CORTECCIA VISIVA PRIMARIA
colonne di orientamento
CORTECCIA VISIVA PRIMARIA:
le ipercolonne
• ogni parte del campo visivo è analizzata da un modulo o
ipercolonna
• questa contiene tutti i circuiti necessari per analizzare
l’informazione proveniente da quell’area di campo visivo
(piccola per la fovea; grande per la periferia)
• Colonne di orientamento (che coprono l’intero arco di 360
gradi)
• Colonne di dominanza sia per l’occhio destro che per il sinistro
• Blob: zone di corteccia nelle quali le cellule sono selettive per i
colori
CORTECCIA VISIVA PRIMARIA:
le ipercolonne
S S
D D
2 mm x 2 mm
Funzioni della corteccia visiva
primaria
• decompone il mondo visivo in brevi segmenti
lineari di orientamento diverso
• separa le informazioni concernenti il colore da
quelle che riguardano le forme ed il movimento
• combina insieme le afferenze dei due occhi come
primo passo di una serie di trasformazioni
necessarie per la visione dalla profondità
input
1
2,3
4
a
b
c
5
6
Strati magnocellulari
NGL
Strati parvocellulari
NGL
Strati parvocellulari NGL e
strati interlaminari
Altre aree
dorsali
magnocellule
MST
MT
V3
Altre aree
ventrali
IT
Parvo.inter-blob
V4
Parvo. blob
V2
V1
Tre vie parallele
• Il sistema magnocellulare: specializzato nell’analisi del
movimento e delle relazioni spaziali fra gli oggetti. Prende
origini dalle cellule gangliari M, occupa i due strati ventrali del
NGL, resta segregato in V1, V2 e V3, per terminare in V5 (MT)
e V5a (MST). Importante per la navigazione, per dirigere i
movimenti oculari e percezione del movimento
•Sistema parvocellulare inter-blob: specializzato per l’analisi
della forme (e in qualche misura dei colori). Parte dalle cellule
gangliari P, occupa i quattro strati dorsali del NGL, resta
segregato in V1 (zone interblob) e V2, arriva in V4 e infine nella
corteccia inferotemporale (IT)
•Sistema parvocellulare blob: specializzato per la visione dei
colori. Parte dalle strati parvocellulari del NGL (ed anche dalle
cellule degli strati interlaminari), resta segregato V1 (zone blob)
e V2, ed arriva a V4
Corteccie visive extrastriate e
il movimento: studi di psicofisiologia e
lesionali
• Il movimento è rappresentato nell’area mediotemporale
(MT o V5) e mediotemporale superiore (MST)
• Oltre ai neuroni selettivi alle singole componenti del movimento
(componente perpendicolare al loro asse di orientamento), un
20%di neuroni selettivi per la direzione globale di sagome
complesse: rispondono alla direzione del movimento percepita
anche se questa è diversa da quella fisica
• La frequenza di scarica dei neuroni rispecchia sia la velocità
oltre alla direzione del movimento
• Neuropsicologia: lesioni dell’area mediotemporale alterano
selettivamente la capacità di analizzare il movimento
(akinetopsia)
Percezione del movimento
globale (illusorio)
+
=
Zihl, 1983
Corteccie visive extrastriate:
il riconoscimento delle forme
•Il riconoscimento delle forme complesse è mediato dalla
corteccia inferotemporale (IT) (sistema parvicellulareinterblob che si estende da V1 a V2 e V4)
•
le cellule dell’area inferotemporale non posseggono una
organizzazione retinotopica
• hanno campi recettivi molto estesi (25° X 25°)
• 10% selettive per particolari stimoli complessi: mani, volti
–Neuropsicologia: lesioni dell’area inferotemporale alterano
selettivamente la capacità di analizzare le fisionomie e le
forme complesse (agnosia)
Collage di volti e pattern casuali vs.
collage composti di soli pattern casuali:
aree inferotemporali
Corteccie visive extrastriate:
il colore
• La visione del colore è mediata dalla via che parte dagli
strati parvocellulari del NGL, proietta ai blob del II e III
strato di V1, poi dopo V2 arriva a V4, aree nella quale vi
sono molte cellule che rispondono al colore
•Campi recettivi molto più grandi con cellule recettive sia
per l’orientamento che il colore
•Area importante per la percezione sia del colore che della
forma (arrivano le afferenze anche della via parvocellulare
inter-blob)
•Lesioni della V4 producono acromatopsia, spesso
accompagnata da disturbi della percezione della forma
(agnosia)
Aree mediali
(V4)
Vs. pattern di varie
tonalià di grigio
Aree più laterali,
MT umana
Stazionario vs
movimento