Modellare il percorso visivo ventrale
Alessio Plebe ([email protected])
Department Cognitive Science; v. Concezione 8
Messina, ITALY
Sommario
Negli studi sulla visione umana è diffuso il paradigma che
vede due circuiti neuronali distinti: quello denominato ventrale, responsabile del riconoscimento di oggetti, contrapposto ad uno dorsale dedicato all’analisi spaziale e dei movimenti. Pur dovendo sottolineare la convenzionalità di questa
semplificazione, che abdica alla natura intrinsecamente olistica dei processi visivi, è proficuo sfruttare l’opportunità di
modellare computazionalmente tale sistema, consentita dalla sua relativa autonomia. Opportunità particolarmente allettante tenuto conto della posta in gioco, si tratta del sistema cognitivo che più di ogni altro tocca il problema filosoficamente centrale dei sistemi neuronali, la loro capacità
semantica, in un quadro di funzioni biologiche sufficientemente delineate. Questo lavoro illustra un modello del percorso ventrale del sistema visivo umano, basato su un’architettura neuronale artificiale, la LISSOM, che rappresenta uno
dei migliori compromessi tra plausibilità biologica e semplicità indispensabile per la modellazione di compiti cognitivi
complessi quali il riconoscimento di oggetti.
Abstract
A shared paradigm in human vision is that cortical areas are
organized into two separate pathways, one called ventral,
crucial for the visual identification of objects, and the other
ventral, specialized for the visual location of objects. This
is still a conventional division, in fact vision, like all other
brain functions, is highly holistic in nature. However, once accepted a certain shift form its faithful reproduction, the
ventral path has enough autonomy to be studied in isolation,
and this is the only way to reduce the natural system to a
complexity suitable for computational models. The opportunity is very interesting, since the objective of the ventral path,
recognition, is on the heart of the representational properties
of neural circuits, a challenging phenomena dense of philosophical implications. In this work a model of the ventral
visual pathway is proposed, based on the neural architecture LISSOM. This is a good compromise between biological
plausibility and the simplicity necessary to build a complex
model involving several cortical areas.
Moduli e modelli
Modellare implica sempre la necessità di operare semplificazioni rispetto alla realtà, ed una delle vie per attuarlo è di
considerare il fenomeno in esame scomponibile in moduli dotati di una relativa autonomia. Nella fattispecie dei modelli
computazionali la pratica diventa particolarmente spontanea
essendo il concetto di modulo del tutto familiare nel trattamento informatico dell’informazione, ed ha trovato autorevoli giustificazioni teoriche nella sua trasposizione ai fenomeni
cognitivi (Fodor, 1983). Ciò che lo stesso nome designa in
ambito neuroscientifico è piuttosto diverso, un modulo è una
popolazione neuronale che presiede ad un insieme coerente
di funzioni piuttosto omogenee, e può avere una differenziazione anatomica, ma raramente possiede le caratteristiche di
autonomia ed isolamento dell’altra accezione di modulo, soprattutto se a livello corticale. A livello cerebrale la caratterizzazione olistica se non dominante è sicuramente non trascurabile, ed è uno dei maggiori ostacoli alla sua comprensione,
non ne è immune la visione (Churchland et al., 1994). Purtuttavia, una volta preso atto dell’allontanamento dalla realtà e
dalla sua completa comprensione, può essere proficuo adottare certe convenzioni semplificatorie, in quanto uniche strade
percorribili per certi scopi modellistici.
Il caso qui preso in considerazione è il ben noto percorso ventrale nel sistema visivo umano, considerato alternativo a quello dorsale, e ritenuto dominante per il riconoscimento di oggetti (Ungerleider and Minshkin, 1982). Mentre
quello dorsale ha una definizione piuttosto controversa (Goodale and Humphrey, 1998), in cui convivono analisi spaziali, del movimento, ma anche sensibilità a proprietà di oggetti, confermando una natura decisamente olistica, il percorso ventrale pare abbastanza specifico rispetto alla funzione del riconoscimento, e quindi si presta meglio a questa
operazione.
Ma più che le sue caratteristiche idonee, la sua attrattiva
è proprio nei riguardi della funzione che sottende. Il riconoscimento è paradigmatico del passaggio dai segnali sensoriali
alla semantica, e il percorso visivo ventrale offre l’opportunità unica di attraversare, nel modellarlo, una delle proprietà
dei neuroni filosoficamente più misteriose, godendo della conoscenza più dettagliata oggi disponibile rispetto a qualunque altro sistema neuronale. Tra i punti che fanno del riconoscimento un problema filosofico aperto Shimon Edelman
(Edelman, 2002) per esempio elenca:
• l’inerente ineffabilità delle immagini;
• lo statuto ontologico discutibile degli oggetti;
• l’impossibilità di segmentazione in base a dei principi;
• evitare la modellazione matematica di funzioni mature;
• la natura olistica delle informazioni in una scena.
• progettare in modo adeguato i meccanismi di plasticità;
Va ricordato inoltre come per la maggioranza degli animali il riconoscere non si possa certo considerare la funzione
propria della visione, rintracciabile piuttosto nel controllo del
movimento. Negli uomini non soltanto lo diventa, ma risulta
anche ineluttabile: non è più possibile farne a meno, anche in
compiti dove non è necessaria (Pins et al., 2004).
• mirare ad una riproduzione coerente delle gerarchie
architetturali;
L’approccio neurocostruzionistico
La strutturazione del modello qui proposto aderisce all’idea
della corteccia organizzata in mappe, che dal momento della sua scoperta (Mountcastle, 1957) ad oggi è rimasto il suo
più fecondo paradigma esplicativo. Tale concetto corrobora
l’ipotesi modulare mediante un’evidenza oggettiva: l’omogeneità delle risposte neuronali in una determinata area rispetto
a certe stimolazioni sensoriali, di cui ne replicano alcune proprietà topologiche. Inizialmente scoperto nello studio della
sensorialità somatica, trovò il suo terreno più fertile nelle successive scoperte dei campi corticali visivi, dove l’individuazione della topologia è facilitata in modo unico dalla natura
spaziale del segnale visivo (Hubel and Wiesel, 1962).
Viene qui inoltre suggerita l’adozione di una prospettiva definibile neurocostruttivista, che dall’omonimo ben noto constructivist manifesto (Quartz and Sejnowski, 1997) trae
due punti fondamentali:
• le proprietà computazionali delle mappe corticali sono
essenzialmente equipotenti;
• lo sviluppo della forma matura delle funzioni computazionali di tali mappe è dettata soprattutto da afferenze
ambientali.
Si tratta di due principi generali riguardanti la corteccia, che
nella visione incontrano una serie notevole di constatazioni neuroscientifiche (Löwel and Singer, 2002; Sengpiel and
Kind, 2002), rendendola la modalità dove la loro adesione,
già motivata teoricamente, diventa anche dovuta alla luce delle evidenze empiriche. Ciononostante occorre aggiungere che
non si tratta affatto di una posizione universalmente condivisa, si oppone infatti alle ben conosciute istanze nativiste. Nel
campo della spiegazione neuroscientifica hanno trovato molto più spazio riguardo ad altre funzioni, come il linguaggio,
dove ovviamente incontrano una consolidata tradizione cognitiva, molto meno in visione, dove sono tuttavia marginalmente presenti nel dibattito, con qualche supporto empirico
(Smirnakis et al., 2005).
Va osservato che l’accezione attuale di neurocostruttivismo
va ben oltre la datata diatriba sull’innatismo mentale, seguendo in modo attento le recenti scoperte sulle fitte interazioni
tra componenti genetiche e processi plastici, su cui proprio
nel sistema visivo cominciano ad emergere dati significativi
(Mjdan and Shatz, 2006), ponendo quindi in un quadro ancor più realistico ed importante le prospettive di modellare lo
sviluppo (Quartz, 2003).
Queste considerazioni, unitamente al vaglio delle altre constatazioni neuroscientifiche menzionate, possono venir tradotte in prescrizioni modellistiche come quelle qui
elencate:
• tentare una ricostruzione del contesto epigenetico.
Tra i metodi matematici circolanti in grado di soddisfare
questi principi sono sicuramente attraenti quelli che rientrano
nella categoria generale dei sistemi auto-organizzanti, la cui
prima adozione nel simulare fenomeni della visione naturale si deve a Christoph von der Malsburg (von der Malsburg,
1973). Può essere considerata nella sua progenie la recente
architettura LISSOM (Laterally Interconnected Synergetically Self-Organizing Map) (Miikkulainen et al., 1997), che abbina una semplicità non troppo lontana da quella della nota
SOM (Kohonen, 1995) ad un’aderenza ai meccanismi corticali biologici decisamente superiore. Nella LISSOM ogni
neurone riceve non solo afferenze, ma anche segnali di tipo
eccitatorio ed inibitorio dai neuroni vicini nella stessa mappa:
γA
(k)
xi = f
arA ,i · vrA ,i +
1 + γN I · vrA ,i
(1)
(k−1)
(k−1)
,
γEerE ,i · xrE ,i − γHhrH ,i · xrH ,i
dove è data l’attivazione x del neurone i al tempo k. Tutti i tre
termini all’interno delle parentesi comprendono un prodotto
vettoriale, esteso in un intorno circolare del neurone i, tra valori delle efficienze sinaptiche e contributi, nell’ordine di tipo
afferente eccitatorio ed inibitorio. L’esigenza di esprimere la
(1) nel tempo discreto è la sua natura ricorsiva, per l’influenza
reciproca dei neuroni vicini in funzione eccitatoria ed inibitoria, e richiede quindi una soluzione iterativa. Tutti i parametri della (1) sono appresi in modo hebbiano, modificando
le efficienze sinaptiche in modo proporzionale alla coincidenza delle attività presinaptiche e postsinaptiche, compensando
con un termine globale che simuli l’omeostasi sinaptica (Turrigiano and Nelson, 2004). Ulteriori dettagli sul calcolo sono
in (Plebe and Domenella, 2006).
Mappe e funzioni
Il modello complessivo è mostrato nella Fig. 1, la sua struttura gerarchica prende l’avvio dalle sensazioni visive a livello
della retina, nei soli canali del rosso e del verde, transitando
per il talamo, e per mappe corticali via via più elevate lungo il
percorso ventrale, ricostruito nei suoi tratti essenziali secondo le mappature neuroscientifiche attualmente più condivise
(Wandell et al., 2005). Mentre per certe aree, come la V1 e la
V2 non ci sono particolari problemi di individuazione, l’area
VO, qui presa in considerazione in quanto cruciale nell’elaborazione cromatica, manca attualmente di una denominazione
e di una delimitazione condivisa, le due tendenza attuali sono nell’usare la denominazione hV4 (human V4, in quanto
anatomicamente differente dall’area V4 nei primati, responsabile dell’elaborazione cromatica) oppure V8 (Brewer et al.,
2005), il generico VO usato qui toglie dall’imbarazzo della
scelta. L’area LOC, pur se meno problematica rispetto a quella del colore, manca pure di una sua precisa identificazione, e
LOC
corticale
V2
VO
V1
acromatico
M+ L−
M− L+
on−center
off−center
LGN
on−center
off−center
L−wavelengths
off−center
intensita‘
on−center
extracorticale
M−wavelengths
Figura 1: Il modello proposto. Le mappe LISSOM corrispondono ad aree corticali: V1 e V2 le aree visive primaria e secondaria,
VO ventro-occipitale, LOC è il complesso laterale occipitale, nella parte extracorticale LGN rappresenta il nucleo laterale
genicolato nel talamo.
la “C” nella sua denominazione corrente indica proprio come
gli studiosi preferiscano prenderla come un’insieme di campi visivi (Complex) piuttosto che una vera e propria mappa
nell’accezione di Vernon Mountcastle.
Il tentativo qui portato avanti è di indurre nelle mappe corticali del modello lo sviluppo delle funzioni che più specificatamente le caratterizzano, e che complessivamente cooperano al riconoscimento vero e proprio. Più precisamente le
funzioni associate ad ogni mappa corticale sono le seguenti:
V1 −→ domini di orientazione
V2 −→ selettività angolare
VO −→ costanza cromatica
LOC −→ invarianza rotazionale
Mentre i domini di orientazione della corteccia visiva primaria sono stati il primo processo corticale su cui è stata fatta
luce (Hubel and Wiesel, 1962), e ad oggi sicuramente il più
studiato in assoluto, gli altri sono molto meno chiari, alcuni di
scoperta molto recente, come l’invarianza nella LOC (GrillSpector et al., 1998), e ancor di più la selettività angolare nella
V2 (Ito and Komatsu, 2004), e su certuni, come la costanza
cromatica, si sa veramente poco.
Tutti questi processi nel modello emergono spontaneamente mediante esposizione ad una serie di condizioni ambientali
che vanno dalla simulazione di onde retinali spontanee prenatali a viste tridimensionali di oggetti reali, solamente la parte
extracorticale è modellata in modo rigido, replicando le celle
complesse a soppressione esterna od interna, mediante differenze di funzioni gaussiane. Come immagini reali è stata
utilizzata la libreria standard COIL-100 (Murase and Nayar,
1995), che comprende 100 oggetti comuni, ciascuno inquadrato sotto 72 diverse angolazioni, per un totale quindi di
7200 fotogrammi. Le onde retinali prenatali sono simulate
mediante macchie sfumate sotto forma di funzioni gaussiane
bidimensionali, allungate lungo un asse a orientazione casuale, e posizionate casualmente nel piano retinale. L’addestramento del modello prevede un’iniziale esposizione alle onde
retinali, sostituite successivamente, in un momento che corrisponderebbe all’apertura degli occhi, con le immagini reali.
Questa sequenza coinvolge la V1 e in parte la V2, mentre
le aree superiori VO e LOC subentrano in tempi successivi,
senza quindi essere sollecitate dalle onde retinali, coerentemente con il quadro attuale sullo sviluppo del sistema visivo
(Sengpiel and Kind, 2002; Gandhi et al., 2005).
Un aspetto di particolare interesse sui risultati del modello
è la diversificazione di funzioni ottenute con un’architettura
di base uniforme, la cui specializzazione è riposta quindi in
due fattori: il posizionamento delle mappe all’interno della
gerarchia, e la differenza in certi loro parametri strutturali,
quali le dimensioni dei campi recettivi e le diverse pesature
delle componenti afferenti, eccitatorie e inibitorie.
I domini di orientazione nella mappa V1 che vengono ottenuti dal modello replicano le loro ben note caratteristiche,
come le aree a variazione continua di direzione dell’orientazione, i punti di sella, punti di rotazione e le discontinuità, sono risultati simici a quelli già disponibili in letteratura (Miikkulainen et al., 1997). Rappresentano invece un’innovazione i domini di selettività angolare ottenute nel modello dal-
rotazione
media
10o
20o
30o
40o
50o
60o
0.904
0.838
0.781
0.729
0.686
0.648
retina
stdv
0.070
0.106
0.140
0.167
0.192
0.221
mappa LOC
media stdv
0.990
0.970
0.943
0.913
0.885
0.848
0.015
0.031
0.056
0.077
0.094
0.120
Tabella 1: Correlazione tra immagini retinali sottoposte a diverse rotazioni del punto di vista dell’oggetto, e le corrispondenti mappe LOC, i risultati sono medie e deviazioni standard
su 100 oggetti.
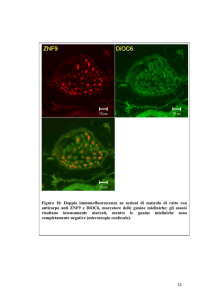
Figura 2: La mappa V2 con sovrapposto ad ogni neurone
l’angolo a cui risulta selettivo. Sono visualizzati solo i neuroni per i quali un angolo prevale su tutti gli altri possibili
almeno del 30%.
la mappa V2, che replicano in modo piuttosto fedele diverse
proprietà di un fenomeno, come detto, di recente scoperta, e
costituiscono il naturale passo successivo alla V1 nel percorso
ventrale, tenendo conto che l’angolo è la primitiva immediatamente più complessa dopo il il segmento orientato. Nella
mappa V2 del modello circa l’80% dei neuroni sviluppa una
sensibilità agli angoli, con prevalenza verso angoli ampi, proprio come riportato nello studio biologico (Ito and Komatsu,
2004).
La costanza cromatica nella VO del modello è in effetti
limitata alla sola parte più essenziale del complesso fenomeno fisico racchiuso dal termine, e consiste nella selettività, da
parte dei neuroni di questa mappa, alle componenti cromatiche della figura, in modo pressoché indipendente dalla loro
intensità.
Verso il riconoscimento
Il percorso corticale ventrale si presta ad essere interpretato come un percorso concettuale verso il riconoscimento di
oggetti. Un primo filo conduttore che dona convincimento a
questa lettura è la progressiva perdita di retinotopia, mentre
nelle prime mappe corticali esiste una netta definizione nella
topologia delle risposte neuronali, sia secondo la dimensione
radiale che eccentrica, man mano che ci si allontana dalle proiezioni talamiche tale caratteristica diviene più sfumata, con
campi recettivi ampi e poco delineati, tanto da far diventare
problematica la stessa accezione di mappa corticale. Un secondo ma non indipendente motivo conduttore è la progressiva insensibilità dalle caratteristiche locali del segnale retinale. Se queste sono caratteristiche che man mano vanno a
perdersi lungo il percorso ventrale, ciò che si guadagna sono
proprietà che orientano sempre più i circuiti neuronali verso
il processamento di entità più globali, in altre parole oggetti.
È interessare indagare quanto ciò avvenga nella mappa
LOC del modello, e in che termini si manifesti. Una delle
principali proprietà attribuite ad un’elaborazione che si collochi a questo livello è la sua invarianza rispetto alla variabilità
Figura 3: Neuroni nella mappa LOC (in basso) attivati in modo persistente su 72 diverse viste di uno stesso oggetto (in
alto).
di apparenza di uno stesso oggetto. Più precisamente nella
visione naturale non esiste invarianza, nel senso che le variazioni del punto di osservazione sono sempre percepite, esiste
una tolleranza, non assoluta, nei confronti di tutti i modi in
cui un oggetto può apparire, che trova la sua prima dimostrata manifestazione, nella visione umana, proprio nell’area
LOC (Grill-Spector et al., 1998). Il fenomeno è riprodotto
dal modello, soprattutto per quanto riguarda l’invarianza rispetto alla rotazione del punto di osservazione, che imprime
le modifiche più significative e più difficili da ovviare all’apparenza retinale di un oggetto, dettagli di quest’indagine sono
in (Plebe, 2006). Il risultato è mostrato nella Tabella 1, come
confronto tra le differenze misurate nell’immagine retinale e
nella risposta dei neuroni in LOC.
L’invarianza è la ripercussione di una capacità della mappa di rispondere ad un livello più astratto all’insieme di proprietà, geometriche e cromatiche, che caratterizzano gli oggetti. Ciò si concretizza nella selettività di alcuni neuroni a
caratteristiche poco individuabili localmente, ma riscontrabili in quanto persistenti al variare dell’apparenza dell’oggetto.
È proprio tale proprietà a permettere di analizzarli, misurando
nella mappa LOC la persistenza dell’attività dei vari neuroni,
per uno stesso oggetto, al variare delle sue pose, differenziandosi in modo statisticamente significativo dall’attivazione su
tutti gli altri oggetti.
Tre esempi sono visibili nella Fig. 3. Ciò che si verifica
nella mappa LOC è una codifica di tipo distribuito, in cui in
Figura 5: Neuroni nella mappa LOC attivati in modo persistente da tutti gli oggetti con colori specifici: a sinistra il rosso
(19 campioni), al centro il verde (10 campioni), e a destra il
blu (7 campioni), sempre su 72 diversi punti di vista per ogni
oggetto.
Figura 4: Neuroni nella mappa LOC attivati in modo persistente da oggetti di una stessa categoria, sempre in tutte e 72
le possibili loro viste: a sinistra le automobili, con 7 esemplari, al centro i vasi con 4 esemplari, a destra gli autocarri con
4 esemplari. Le immagini nella riga in alto sono esempi della
categoria.
una popolazione di neuroni l’attivazione di un certo numero
caratterizza specificatamente un oggetto, e cooperativamente tutti concorrono nel rappresentare il mondo di oggetti di
cui è stata fatta esperienza. Naturalmente in ogni singola vista diventeranno attivi nella mappa LOC anche altri neuroni,
che rappresentano le specificità di quella vista, il potenziale dei neuroni persistenti non è in alcun modo predeterminato, ma emerge unicamente dall’essere quelli che colgono
le similarità tra viste in uno stesso oggetto, in contrasto con
altri.
Viceversa esistono anche popolazioni di neuroni che tendono ad attivarsi in misura analoga per oggetti similari, sempre
nel totale di tutte le loro viste, e ciò può essere misurato in
modo analogo, contrastando la persistenza in oggetti di uno
stesso gruppo, rispetto alla risposta per tutti gli altri oggetti,
sempre presentati in tutte le loro possibili viste. Il risultato è
mostrato in tre esempi nella Fig. 4.
Le caratteristiche che la mappa LOC estrae implicitamente
sono di ordine superiore, basandosi su segnali che sono già
caratterizzati dal processo di proprietà evidenziate dalle mappe sottostanti. È prevedibile quindi che in qualche modo i
neuroni presentino una codifica ancor più globale rispetto a
tali proprietà, per esempio quelle di costanza cromatica. A
tale scopo è stato applicato lo stesso metodo a gruppi di oggetti selezionati per avere una dominanza cromatica tipica, ed
è effettivamente emersa la presenza di un numero consistente di neuroni che codificano selettivamente per certi colori, i
risultati sono mostrati nella Fig. 5.
I fenomeni appena descritti mostrano come la parte terminale del percorso visivo ventrale qui modellato raggiunga un
genere di codifica prossimo a quello che possiamo definire
riconoscimento di oggetti. È bene chiarire che è purtuttavia
una funzione ben lontana dal coincidere con una rappresentazione vera e propria di oggetti o ancor meno di categorie o
proprietà, come la colorazione verde. Lo è anzitutto per la sua
collocazione corticale, nell’essere ancora interamente parte di
una modalità specifica, quella visiva, ma ancor di più per aver
reso tale modalità impermeabile, nell’assunzione iniziale di
voler modellare il percorso ventrale in totale autonomia.
Come anticipato nell’introduzione tale ipotesi è feconda,
consentendo una modellazione computazionale, ma imprecisa, essendo anche il percorso ventrale già contaminato da proiezioni non dipendenti dai soli segnali retinali ma da altre parti del cervello. Quindi già a livello dell’area LOC biologica
sono tutt’altro che trascurabili gli effetti dovuti all’esperienza
corporea complessiva che si correla ogni volta alla semplice
visione degli oggetti, incluso il ruolo emozionale che giocano
tagli oggetti per l’individuo. Ma ancor di più, una completa
caratterizzazione rappresentazionale degli oggetti come entità non può che aver luogo oltre l’area occipitale, laddove si
fondono in modo sostanziale le diverse modalità, come nella
corteccia prefrontale (Freedman et al., 2001).
E parlando di visione nell’uomo è evidente quanto sia carente un quadro modellistico dei processi di riconoscimento
che ignori la componente linguistica, che pervade il nostro
modo di segmentare il mondo in entità.
Pur nella piena consapevolezza di tali limitazioni, si ribadisce l’utilità dell’indagine computazionale sul percorso ventrale, che è unico nel dettaglio disponibile sulla ripartizione
dei suoi circuiti neuronali, e di cui si può dimostrare l’effetto
di composizionalità di funzioni ben precise, in una gerarchia
che arriva, come visto, sufficientemente vicino a ciò che Martha Farah ha definito il miracolo del riconoscimento di oggetti
(Farah, 2000).
Riferimenti bibliografici
Brewer, A. A., Liu, J., Wade, A. R., and Wandell, B. A. (2005).
Visual field maps and stimulus selectivity in human ventral
occipital cortex. Nature Neuroscience, 8:1102–1109.
Churchland, P. S., Ramachandran, V., and Sejnowski, T. (1994).
A critique of pure vision. In Koch, C. and Davis, J., editors, Large-Scale Neuronal Theories of the Brain. MIT Press,
Cambridge (MA).
Edelman, S. (2002). Constraining the neural representation of the
visual world. Trends in Cognitive Sciences, 6:125–131.
Farah, M. J. (2000). The Cognitive Neuroscience of Vision. Basil
Blackwell, Oxford (UK).
Fodor, J. (1983). Modularity of Mind: and Essay on Faculty
Psychology. MIT Press, Cambridge (MA).
Freedman, D. J., Riesenhuber, M., Poggio, T., and Miller, E. K.
(2001). Categorical representation of visual stimuli in the
primate prefrontal cortex. Science, 291:312–316.
Gandhi, S. P., Cang, J., and Stryker, M. P. (2005). An eye-opening
experience. Nature Neuroscience, 8:9–10.
Goodale, M. A. and Humphrey, G. K. (1998). The objects of actions
and perception. In Tarr, M. J. and Bülthoff, H. H., editors,
Object Recognition in Man, Monkey, and Machine. MIT Press,
Cambridge (MA).
von der Malsburg, C. (1973). Self-organization of orientation
sensitive cells in the striate cortex. Kibernetic, 14:85–100.
Grill-Spector, K., Kushnir, T., Edelman, S., Itzchak, Y., and Malach,
R. (1998). Cue-invariant activation in object-related areas in
the human occipital lobe. Neuron, 21:191–202.
Wandell, B. A., Brewer, A. A., and Dougher, R. F. (2005). Visual
field map clusters in human cortex. Philosophical transactions
of the Royal Society of London, 360:693–707.
Hubel, D. and Wiesel, T. (1962). Receptive fields, binocular interaction, and functional architecture in the cat’s visual cortex.
Journal of Physiology, 160:106–154.
Ito, M. and Komatsu, H. (2004). Representation of angles embedded within contour stimuli in area V2 of macaque monkeys.
Journal of Neuroscience, 24:3313–3324.
Kohonen, T. (1995). Self-Organizing Maps. Springer-Verlag, Berlin.
Löwel, S. and Singer, W. (2002). Experience-dependent plasticity of
intracortical connections. In Fahle, M. and Poggio, T., editors,
Perceptual Learning. MIT Press, Cambridge (MA).
Miikkulainen, R., Bednar, J. A., Choe, Y., and Sirosh, J. (1997).
Self-organization, plasticity, and low-level visual phenomena
in a laterally connected map model of the primary visual cortex. In Goldstone, R. L., Schyns, P. G., and Medin, D. L.,
editors, Psychology of Learning and Motivation, volume 36,
pages 257–308. Academic Press, New York.
Mjdan, M. and Shatz, C. J. (2006). Effects of visual experience on
activity-dependent gene regulation in cortex. Neurocomputing,
in press.
Mountcastle, V. (1957). Modality and topographic properties of
single neurons in cats somatic sensory cortex. Journal of
Neurophysiology, 20:408–434.
Murase, H. and Nayar, S. (1995). Visual learning and recognition of
3-d object by appearence. International Journal of Computer
Vision, 14:5–24.
Pins, D., Meyer, M. E., Foucher, J., Humphreys, G., and Boucart,
M. (2004). Neural correlates of implicit object identification.
Neuropsychologia, 42:1247–1259.
Plebe, A. (2006). Learning visual invariance. In Verleysen, M.,
editor, ESANN 2006 – 14th European Symposium on Artificial Neural Networks, pages 71–76, Evere (BE). d-side
Publications.
Plebe, A. and Domenella, R. G. (2006). Early development of visual
recognition. BioSystems, 86:63–74.
Quartz, S. R. (2003). Innateness and the brain.
Philosophy, 18:13–40.
Biology and
Quartz, S. R. and Sejnowski, T. J. (1997). The neural basis of cognitive development: a constructivist manifesto. Behavioral and
Brain Science, 20:537–596.
Sengpiel, F. and Kind, P. C. (2002).
The role of activity
in development of the visual system. Current Biology,
12:818–826.
Smirnakis, S. M., Brewer, A. A., Schmid, M. C., Tolias, A. S.,
Schüz, A., Augath, M., Inhoffen, W., Wandell, B. A.,
and Logothetis, N. K. (2005). Lack of long-term cortical reorganization after macaque retinal lesions. Nature,
435:300–307.
Turrigiano, G. G. and Nelson, S. B. (2004). Homeostatic plasticity in
the developing nervous system. Nature Reviews Neuroscience,
391:892–896.
Ungerleider, L. and Minshkin, M. (1982). Two cortical visual systems. In Ingle, D. J., Goodale, M. A., and Mansfield, R. J. W.,
editors, Analysis of visual behavior. MIT Press, Cambridge
(MA).